142 책임저자:황원덕, 614-052, 부산시 진구 양정2동 산45번지
동의대학교 한의과대학 내과학교실 Tel: 051-850-8625, Fax: 051-853-4306 E-mail: [email protected]
접수일:2008년 5월 28일, 게재승인일:2008년 6월 19일
Correspondence to:Won Deok Hwang
Department of Internal Medicine, Dongeui University College of Oriental Medicine, San 45, Yangjeong 2-dong, Jin-gu, Busan 614-052, Korea Tel: +82-51-850-8625, Fax: +82-51-853-4306
E-mail: [email protected]
부자 추출물에 의한 인체 간암세포의 Apoptosis 유도에 관한 연구
동의대학교 한의과대학 내과학교실, 1생화학교실 및 대학원 바이오물질제어학과
임동진ㆍ최영현1ㆍ황원덕
Induction of Apoptoticcell Death by Ethanolic Extract of Aconiti Tuber in Human Hepatocarcinoma Cells
Dong Jin Lim, Yung Hyun Choi1 and Won Deok Hwang
Department of Internal Medicine, Dongeui University College of Oriental Medicine,
1Department of Biochemistry, Dongeui University College of Oriental Medicine and Department of Biomaterial Control, Dongeui University Graduate School, Busan 614-052, Korea
Aconiti tuber has been used in oriental herbal medicine since ancient times for analgesic, anti-rheumatic and neurological indications. In the present study, it was investigated the biochemical mechanisms of anti-proliferative effects by the methanol extract of A. Tuber (MEBJ) in HepG2 and Hep3B human hepatocarcinoma cells. It was found that MEBJ could inhibit the cell proliferation of HepG2 and Hep3B cells in a concentration-dependent manner, which was associated with induction of apoptotic cell death such as formation of apoptotic bodies and increased populations of apoptotic-sub G1 phase. However, Hep3B cell treated with MEBJ reduced cell viability and induced apoptosis significantly more than in HepG2 cells. Apoptosis of Hep3B cells by MEBJ was connected with an up-regulation of pro-apoptotic Bax expression and a down-regulation of anti-apoptotic Bcl-2 and Bcl-xL in a dose-dependent manner. MEBJ treatment up-regulated the levels of Fas mRNA and protein, and the expression levels of IAPs family members such as cIAP-1 and cIAP-2 were down-regulated by MEBJ treatment. In addition, MEBJ treatment induced the proteolytic activation of caspase-3 and a concomitant degradation of poly (ADP-ribose) polymerase protein. Though further studies will be needed to identify the active compounds and mechanisms that confer the anti-cancer activity of MEBJ, these findings suggest that MEBJ may be a potential chemotherapeutic agent for the control of human cancer cells. (Cancer Prev Res 13, 142-152, 2008)
Key Words: Aconiti tuber, Apoptosis, Bcl-2, Caspase-3
서 론
Programed cell death로 알려진 apoptosis는 개체의 발생 이나 항상성 유지 조절에 중요한 역할을 하는 정상적인
생리학적 현상이다. 최근 apoptosis 조절에는 많은 유전자 들이 관여하고 있으며, 이들 유전자 산물들의 기능과 특 징을 조사함으로서 apoptosis 현상의 분자생물학적 설명 이 가능해 지고 있다. 따라서 효과적인 암의 치료와 예방 에 있어서 비정상적인 세포나 암세포의 apoptosis 유발은
많은 치료제의 표적이 되고 있다.1) Apoptosis는 세포주기 교란 의존적 및 비의존적으로 일어날 수 있으며, apo- ptosis의 현상으로 세포질 및 염색질 응축, 세포막 수포화 현상, DNA 단편화 등이 수반되는데 이러한 현상은 세포 내부의 정교한 신호전달에 의해 조절된다.2) Apoptosis 조 절에 관여하는 가장 중요한 인자는 Bcl-2 family에 속하는 단백질들로서 apoptosis의 유도와 억제를 동시로 조절하 는 인자들로 구성되어 있다.3) 또한, caspase라고 이름 붙 여진 protease 역시 apoptosis 유발에 중요한 역할을 하는 데, 이들은 정상적으로 증식 중인 세포에서는 proenzyme 형태로 존재한다.4,5) 이들은 apoptosis 유도 신호에 의해 활성화되어 직접 또는 간접적으로 세포 내 존재하는 많 은 표적 단백질의 분해에 관여한다.5∼7) 이외에도 apo- ptosis에 관여하는 여러 인자들 중에서 inhibitor of apo- ptosis proteins (IAPs) family에 속하는 단백질들은 외부 신 호에 의한 세포 내 anti-apoptotic 활성을 지닌다. 이들 중 몇몇 IAPs는 caspase와의 직접적인 결합을 통하여 그들의 apoptotic 활성을 억제할 수 있는 것으로 알려져 있다.8) 附子(부자, Aconiti Tuber)는 미나리아재비과에 속한 다 년생 草本인 재배종 바곳의 塊根에 附生한 子根으로 한 의학에서는 回陽救逆, 補火助陽, 溫中止痛, 逐風寒濕邪, 補腎命火하는 효능이 있다고 알려져 있어, 亡陽으로 虛 脫한 證, 命門火衰, 下元虛冷의 證, 三焦厥逆, 六府寒冷, 寒濕痿躄 및 噎膈反胃, 癥瘕積聚에 사용되었다.9∼11) 약 리학적으로는 강심, 항부정맥, 혈관 확장 등의 심혈관계 에 대한 작용, 항한냉, 진통 및 진정, 국소 마취 작용 등 신경계에 대한 작용, 항염 작용 및 면역 증강 작용이 있 다고 알려져 있다.10,12) 최근 Nemoto 등13)의 연구에 의하 면 부자 열수 추출물이 정상 세포에는 독성이 없는 범위 내에서 radical scavenging 활성을 보이면서 인체 구강 종 양세포의 증식을 효과적으로 저해하였다고 보고한 바 있다. 이는 부자 추출물이 강력한 항암 효능을 가질 수 있음을 의미하지만 부자 추출물에 대한 항암 작용에 관 한 구체적인 연구가 진행된 바는 없다. 본 연구실에서는 부자 추출물의 항암 효능에 관한 연구의 일환으로 신장 암세포의 증식에 미치는 다양한 부자 추출물의 영향을 조사한 바 있으며, 조사된 추출물 중 methanol 추출물이 가장 강력한 암세포 증식 저해 효과가 있었음을 보고한 바 있다.14)
본 연구에서는 부자 추출물의 항암작용에 관한 부가 적인 자료를 얻기 위하여 부자의 methanol 추출물이 인 체 간암세포의 증식에 미치는 영향을 조사하였으며, 간 암세포의 증식 억제가 apoptosis 유도와 연관성이 있음을 확인하였다. 또한 apoptosis 조절에 중요한 유전자들의 발
현 및 활성 변화를 조사하여 부자 methanol 추출물에 의 한 간암세포의 apoptosis 유도에 관한 기전 해석을 시도하 였다.
재료 및 방법
1. 세포배양 및 부자 methanol 추출물의 처리
본 실험에 사용된 간암세포(HepG2 및 Hep3B)는 한국 생명공학연구소(KRIBB, 대전)에서 분주 받아 10% fetal bovine serum (FBS, Gibco BRL, Grand Island, NY)에 1%의 penicillin 및 streptomycin (Gibco BRL)이 포함된 90%의 RPMI-1640 배지(Gibco BRL)를 사용하여 37oC, 5% CO2 조 건 하에서 배양하였다. 부자의 methanol 추출물(metha- nolic extract of A. Tuber, MEBJ)은 선행 조건에 제시한 방 법에 준하여 준비하였으며14) dimethylsulfoxide (DMSO, Amresco, Solon, OH)에 용해하여 여과를 거쳐 배지에 희 석하여 처리하였다.
2. MTT assay에 의한 암세포 증식 억제 조사
세포 배양용 6 well plate에 HepG2 및 Hep3B 세포를 5×104개/ml로 분주하고 24시간 동안 안정화시킨 후, MEBJ를 배지에 희석하여 각 well 당 0.2∼1.0 mg/ml의 농 도로 처리하였다. 72시간 후 배지를 제거하고 tetrazolium bromide salt (MTT, Amresco)를 0.5 mg/ml 농도가 되게 배 지로 희석하여 2 ml씩 분주하고 3시간 동안 CO2 incu- bator에서 배양시킨 다음 MTT 시약을 깨끗하게 제거하 고 DMSO를 1 ml씩 분주하여 well에 생성된 formazin을 모 두 녹인 후 96 well plate에 200μl씩 옮겨서 ELISA reader (Molecular Devices, Sunnyvale, CA)로 540 nm에서 흡광도를 측정하였다. 측정은 모두 세 번을 하였으며, 그에 대한 평균값과 표준 오차를 Sigma Plot 4.0 프로그램(SPSS Ins.) 으로 구하였다.
3. Hemocytometer를 이용한 세포 생존율의 측정
MTT assay에서와 동일 조건에서 EMBJ가 처리된 배지 에서 배양된 세포를 부유시킨 다음 phosphate-buffered saline (PBS)를 각 well 당 적정량을 첨가하여 세포를 모은 후 2,000 rpm으로 5분간 원심분리 하였다. 상층액을 제거 하고 세포만 남긴 다음 다시 PBS를 1 ml 첨가하여 잘 섞 은 후 세포 부유액과 0.5% trypan blue (Gibco BRL)를 동량 으로 섞어 2분간 처리하였다. Pasteur pippette의 모세관 현상을 이용하여 세포를 hemocytometer로 옮긴 후 도립 현미경(Carl Zeiss, Germany)을 이용하여 푸른색으로 염색 된 세포를 죽은 세포로 추정하고 염색되지 않은 세포를
살아있는 세포수로 계수하였다.
4. DAPI staining에 의한 세포핵의 형태 관찰
다양한 MEBJ 처리에 의한 암세포의 apoptosis 유발 여 부 확인을 위한 핵의 형태적 변화를 관찰하기 위하여 준 비된 세포를 모은 다음, 37% formaldehyde 용액과 PBS를 1:9의 비율로 섞은 fixing solution을 모아진 세포에 500 μl 첨가하여 충분히 섞은 후, 상온에서 10분 동안 고정 하였다. 1,000 rpm으로 5분간 원심 분리한 후 상층액을 제거하고 PBS 200μl를 넣어서 충분히 섞은 후, slide glass 위에 80μl 정도 떨어뜨려 900 rpm에서 5분간 cytospin하 였다. PBS로 2∼3회 washing하고 PBS가 다 마르기 전에 0.2%의 Triton X-100 (Amresco)을 첨가하여 상온에서 10 분간 고정하였다. 그 후 다시 PBS로 washing하고 4',6- diamidino-2-phenylindole (DAPI, Sigma, St. Louis, MO) 용액 을 세포가 고정된 slide glass 위에 적당량을 떨어뜨린 후 빛을 차단하고 상온에서 염색시켰다. 15분 정도 염색시 킨 후, PBS로 DAPI 용액을 충분하게 세척하고 증류수로 재빨리 세척한 다음 absolute alcohol을 이용하여 탈수과정 을 거친 slide glass 위에 mounting solution을 처리한 후 형 광 현미경(Carl Zeiss)을 이용하여 400배의 배율로 각 농도 에 따른 핵의 형태 변화를 관찰하였다.
5. DNA flow cytometry에 의한 분석
정상 및 MEBJ 처리에 따른 세포주기 빈도의 변화를 조사하기 위하여 MEBJ를 처리한 배지에서 72시간 동안 배양시킨 세포들을 PBS로 씻어 내고 0.05% trypsin-EDTA 를 처리하여 부유시킨 다음 1,000 rpm으로 10분간 원심 분리하여 상층액을 버리고 세포들만 모았다. 여기에 다 시 PBS를 첨가하여 충분히 씻은 다음 1,000 rpm으로 10 분간 원심분리 한 후 상층액만 버리고 남은 세포에 0.5 ml의 PBS로 잘 부유시키고, 차가운 ethanol 0.5 ml을 첨가 하여 4oC에서 1 시간 동안 고정시켰다. 5×106개의 고정된 세포들을 원추형 vial에 넣어서 1,000 rpm으로 수 분간 원 심 분리하여 상층액을 제거하고, 1% bovine serum albumin (BSA, Sigma)이 함유된 PBS로 2∼3회 washing 과정을 거친 후 다시 수 분간 원심 분리하였다. 세포 침전물을 1%
BSA가 함유된 PBS 0.8 ml로 부유시키고 DNA intercalating dye propidium iodide (PI, concentration, 50μg/ml, Sigma)와 0.1 mg/ml의 RNase (Sigma)를 처리하여 암실, 4oC 조건에 서 1시간 동안 염색과정을 거쳤다. 이를 PBS로 두 번 washing 과정을 거친 후, 부유액을 만들고 35μm pore size 의 nylon mesh에 부유액을 pipette으로 통과시켜 단일 세 포로 분리시킨 후 DNA flow cytometry (Becton Dickinson,
San Jose, CA)에 적용시켜 형광반응에 따른 histogram을 ModiFit LT program을 사용하여 분석하였다.
6. RT-PCR에 의한 mRNA 발현의 분석
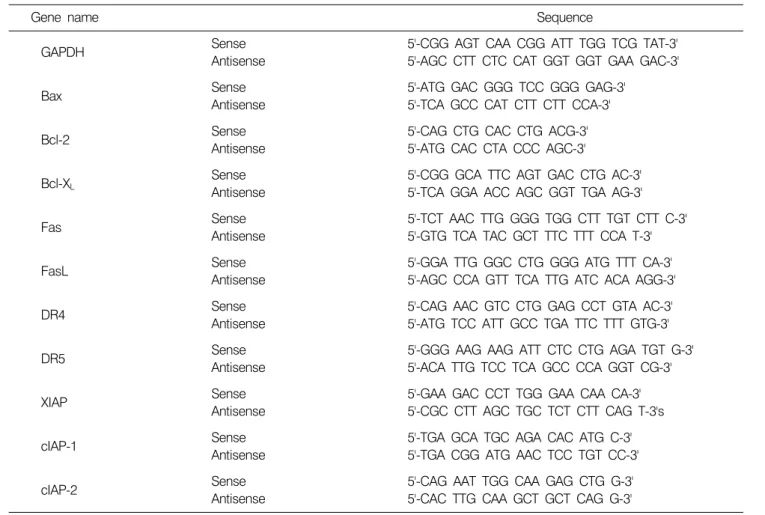
상기와 동일한 조건에서 준비된 세포를 PBS로 세척하 고 TRIzol reagent (Invitrogen Co., Carlsbad, CA)를 4oC에서 1시간 동안 처리하여 total RNA를 분리하였다. 분리된 RNA를 정량한 후, 각각의 primer (Table 1), DEPC water 그리고 ONE-STEP RT-PCR PreMix Kit (Intron, Korea)를 넣 고 Mastercycler gradient (Eppendorf, Hamburg, Germany)를 이용하여 증폭하였다. 각 PCR 산물들의 양적 차이를 확 인하기 위하여 1x TAE buffer로 1% agarose gel을 만들고 well 당 각각의 primer에 해당하는 PCR 산물에 DNA gel loading solution을 섞어서 loading 한 후 100 V에서 전기영 동을 하였다. 전기영동으로 DNA 분리가 끝난 gel을 ethi- dium bromide (EtBr, Sigma)를 이용하여 염색한 후 UV 하 에서 확인하였다. 이때 housekeeping 유전자인 glyceraldehyde- 3-phosphate dehydrogenase (GAPDH) 유전자를 internal con- trol로 사용하였다.
7. Western blot analysis에 의한 단백질 발현의 분석
정상 및 MEBJ가 처리된 배지에서 자란 세포들을 PBS 로 씻어 내고 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM pheny- methylsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 첨가하여 4oC에서 30 분간 반응시킨 후, 14,000 rpm으로 30 분간 원심분리하여 그 상층액을 취하였다. 상층액의 단백질 농도는 Bio-Rad 단백질 정량 시약(Bio-Rad, Her- cules, CA)과 그 사용방법에 따라 정량 한 다음 동량의 Laemmli sample buffer (Bio-Rad)를 섞어서 sample을 만들었 다. 이렇게 만든 동량의 단백질을 sodium dodecyl sulphate (SDS)-polyacrylamide gel을 이용하여 전기영동으로 분리하 였다. 분리된 단백질을 함유한 acrylamide gel을 nitrocellu- lose membrane (Schleicher and Schuell, Keene, NH)으로 electroblotting에 의해 전이시킨 후, 5% skim milk를 함유 한 PBS-T (0.1% Tween 20 in PBS)에 담가 상온에서 1시간 정도 incubation하여 비특이적인 단백질들에 대한 block- ing을 실시하고 PBS-T로 15분정도 세척하였다. 준비된 membrane에 1차 항체를 처리하여 상온에서 1시간 이상 또는 4oC에서 over night 시킨 다음 PBS-T로 세척하고 처 리된 1차 항체에 맞는 2차 항체를 사용하여 상온에서 1 시간 정도 반응시켰다. 다시 PBS-T로 세척하고 Enhanced Chemiluminoesence (ECL) 용액(Amersham Life Science Corp., Arlington Heights, IL)을 적용시킨 다음 암실에서 X-ray
Table 1. Gene-specific primers for RT-PCR in the present study
Gene name Sequence
Sense 5'-CGG AGT CAA CGG ATT TGG TCG TAT-3'
GAPDH
Antisense 5'-AGC CTT CTC CAT GGT GGT GAA GAC-3'
Sense 5'-ATG GAC GGG TCC GGG GAG-3'
Bax Antisense 5'-TCA GCC CAT CTT CTT CCA-3'
Sense 5'-CAG CTG CAC CTG ACG-3'
Bcl-2
Antisense 5'-ATG CAC CTA CCC AGC-3'
Sense 5'-CGG GCA TTC AGT GAC CTG AC-3'
Bcl-XL
Antisense 5'-TCA GGA ACC AGC GGT TGA AG-3'
Sense 5'-TCT AAC TTG GGG TGG CTT TGT CTT C-3'
Fas Antisense 5'-GTG TCA TAC GCT TTC TTT CCA T-3'
Sense 5'-GGA TTG GGC CTG GGG ATG TTT CA-3'
FasL Antisense 5'-AGC CCA GTT TCA TTG ATC ACA AGG-3'
Sense 5'-CAG AAC GTC CTG GAG CCT GTA AC-3'
DR4 Antisense 5'-ATG TCC ATT GCC TGA TTC TTT GTG-3'
Sense 5'-GGG AAG AAG ATT CTC CTG AGA TGT G-3'
DR5 Antisense 5'-ACA TTG TCC TCA GCC CCA GGT CG-3'
Sense 5'-GAA GAC CCT TGG GAA CAA CA-3'
XIAP Antisense 5'-CGC CTT AGC TGC TCT CTT CAG T-3's
Sense 5'-TGA GCA TGC AGA CAC ATG C-3'
cIAP-1
Antisense 5'-TGA CGG ATG AAC TCC TGT CC-3'
Sense 5'-CAG AAT TGG CAA GAG CTG G-3'
cIAP-2
Antisense 5'-CAC TTG CAA GCT GCT CAG G-3'
film에 감광시켜 특정단백질의 양을 분석하였다. Immu- noblotting을 위해 2차 항체로 사용된 peroxidase-labeled donkey anti-rabbit 및 peroxidase-labeled sheep anti-mouse immunoglobulin은 Amersham Life Science에서 구입하였다.
8. In vitro caspase activity 측정
Caspase의 in vitro 활성 측정을 위한 colorimetric assay kits 는 R&D Systems (Minneapolis, MN)에서 구입하였으며, 제 시된 방법에 준하여 활성의 증감 여부를 조사하였다. 이 를 위하여 정상 및 MEBJ가 처리된 배지에서 72시간 배 양된 세포를 모은 뒤 단백질을 추출하고 정량하여 각각 100μg의 단백질을 fluorogenic peptide 기질 100 uM이 함 유된 extraction buffer [40 mM HEPES (pH 7.4), 20% glycerol (v/v), 1mM EDTA, 0.2% NP-40 and 10mM DL-DTT] 50μl 에 혼합하였으며, microtiter plate에 다시 extraction buffer 에 희석하여 각 sample 당 총 volume이 100μl가 되게 하 였다. 실험에 사용된 기질은 caspase-3은 asp-glu-val-asp (DEVD)-p-nitroaniline, caspase-8은 Ile-Glu-Thr-Asp (IETD)- p-pNA이었으며, caspase-9는 Leu-Glu-His-Asp (LEHD)-pNA
였다. 준비된 plate를 37oC에서 2 시간 동안 incubation 시 킨 후 ELISA reader를 이용하여 405 nm의 흡광도를 이용 하여 반응의 정도를 측정하였다.
결 과
1. 인체 간암세포의 증식에 미치는 MEBJ 영향
HepG2 및 Hep3B 인체 간암세포의 증식에 미치는 MEBJ의 영향을 측정하기 위하여 MTT assay에 의한 세포 생존률을 측정한 결과는 Fig. 1에 나타낸 바와 같다. 결과 에서 알 수 있듯이 HepG2 세포의 경우 0.6 mg/ml 처리군 에서 대조군에 비하여 약 70% 정도의 증식률을 보였으 며, 1.0 mg/ml 처리군의 경우 60% 정도의 증식률을 보였 다. 그러나 Hep3B 세포의 경우 MEBJ에 대한 감수성 HepG2 세포에 비하여 높게 나타나 0.6 mg/ml 처리군에 서 60% 이하의 증식률을 보였으며, 1.0 mg/ml 처리군의 경우 17% 정도의 증식률을 보였다. 즉 동일한 조건에서 비교된 MEBJ의 증식 억제 효과 실험에서 HepG2 세포에 비하여 Hep3B 세포에서 증식억제 효과가 매우 높게 나
Fig. 1. Growth inhibition of human hepatocarcinoma cells after treatment with methanol extract of A. Tuber (MEBJ). HepG2 and Hep3B cells were plated at 5×104 cells per/well of 6 well plate and incubated for 24 h. The cells were treated with variable concentrations of MEBJ for 72 h. The growth inhibition was measured by the metabolic-dye-based MTT assay.
Results are expressed as the means±SD of three independent experiments.
Fig. 2. Induction of morphological changes by MEBJ treatment in human hepatocarcinoma HepG2 and Hep3B cells. The cells were treated with various concentrations of MEBJ for 72 h and cell morphology was visualized by an inverted microscopy.
Magnification, ×200.
타났음을 알 수 있었다.
2. 인체 간암세포의 형태에 미치는 MEBJ의 영향
MEBJ 처리에 따른 간암세포의 증식억제가 apoptosis 유발과 연관이 있을 것으로 기대되어 MEBJ 처리 후 암 세포 전체의 형태변화를 비교하고 암세포 핵의 형태변 화를 DAPI 염색을 통하여 조사하였다. Fig. 2A에 나타낸 바와 같이 도립 현미경하에서 관찰된 결과에 의하면 EMBJ 처리 농도가 증가하면 할수록 Hep3B 세포의 mem- brane shrinking 현상을 관찰할 수 있었으며, 밀도 감소와 함께 세포 내 과립 형성과 같은 apoptosis가 유발되었을 경우 관찰되는 세포변형 현상을 관찰할 수 있었다. 또한 정상 조건에서 배양된 세포에 비하여 MEBJ가 처리된 경 우 세포의 길이가 길어지고 denrite와 같은 분지의 형성 이 많아졌다. 그러나 HepG2 세포에서는 이러한 암세포 의 형태 변화의 정도가 심하게 나타나지는 않아 MEBJ 처리에 따른 세포 증식 억제와 부합되는 결과임을 알 수 있었다. 그리고 Fig. 2B의 결과에서 알 수 있듯이 정상 배지서 자란 세포들은 핵의 전체가 완전한 형태로 염색 되는 양상을 보였으나, MEBJ을 처리한 경우 처리 농도 가 증가할수록 apoptosis 유발 특이적인 핵 내 DNA 단편 화에 의한 염색질의 응축된 형태인 apoptotic body의 형성 정도가 증가되었으며, 이는 Hep3B 세포에서 더욱 높게 나타났다(Fig. 3).
3. MEBJ 처리에 의한 간암세포의 생존율 저하 및 apoptosis 유도
MEBJ 처리에 따른 간암세포의 증식억제는 암세포의 생존율 저하 및 apoptosis 유도와 연관성이 있는 것으로 나타나 이를 정량적으로 비교하기 위하여 다양한 농도 의 MEBJ로 처리한 배지에서 자란 세포를 trypan blue로 염색하고, 염색되지 않은 살아있는 세포를 hemocytometer 를 이용하여 계수하여 정상 배지에서 배양된 암세포의 수와 비교하였다. 또한 동일한 조건으로 배양된 세포들 을 대상으로 DNA flow cytometry 분석을 통한 apoptosis가 일어난 세포군에 해당되는 세포주기의 sub-G1기에 해당 되는 세포들의 빈도를 정량적으로 비교하였다. Fig. 4A 에 나타낸 바와 같이, HepG2 세포에 비하여 Hep3B 세포 에서 MEBJ 처리에 따른 생존율의 저하 현상이 매우 뚜 렷하였으며, Fig. 4B에서 알 수 있듯이 MEBJ의 처리 농도 증가에 따라 apoptosis가 유발된 세포의 빈도가 증가되었
Fig. 3. Induction of apoptotic bodies by MEBJ treatment in human hepatocarcinoma HepG2 and Hep3B cells. The cells grown under same condition as Fig. 2 were sampled, fixed and stained with DAPI for 15 min. The stained nuclei were then observed under a fluorescent microscope using a blue filter.
Magnification, ×400.
Fig. 4. Inhibition of cell viability and increase of sub-G1 cell population by MEBJ treatment in human hepatocarcinoma HepG2 and Hep3B cells. (A) HepG2 and Hep3B cells were treated with variable concentrations of MEBJ for 72 h. The cells were trypsinized and the viable cells were scored by hemocytometer counts of trypan blue-excluding cells. (B) The cells were exposed for 72 h with increasing concentration of MEBJ. Then the cells were collected and stained with PI for flow cytometry analysis. The percentages of cells with hypodiploid DNA (sub-G1 phase) contents represent the fractions undergoing apoptotic DNA degradation.
Results are expressed as the means±SD of three independent experiments.
음을 알 수 있었다. 그러나 최고 농도인 1.0 mg/ml 처리 군에서 Hep3B 세포의 경우 55% 정도로 관찰되었음에 비하여 HepG2 세포의 경우 10% 내외로 나타나 MEBJ의 apoptosis 유도에서도 Hep3B 세포에서 감수성이 높음을 알 수 있었다.
4. Death receptors, Bcl-2 및 IAPs family의 발현에 미치는 MEBJ의 영향
이상의 결과에서 MEBJ가 HepG2 세포보다 Hep3B 세 포에서 증식억제 및 apoptosis 유발 효과가 높게 나타났으 므로, MEBJ에 의한 apoptosis 유발 시 관찰되는 주요 apoptosis 조절 관련 유전자들의 발현 변화를 RT-PCR 및 Western blotting의 방법을 이용하여 조사하였다. 조사된 유전자 중 Bcl-2 family에 속하는 인자들은 apoptosis 유발 조절에 가장 대표적 조절인자로 알려져 있는데, 그 중 Bcl-2와 Bcl-xL는 anti-apoptotic 인자로서 apoptosis의 유발 을 억제하는 기능을 가지며, Bax와 Bad는 pro-apoptotic 인 자로 apoptosis의 유발과 관계가 있다. Fig. 5에 나타낸 결 과에서 알 수 있듯이 apoptosis를 유발하는 것과 관련이 있는 Bax 유전자의 발현이 MEBJ 처리에 의하여 전사와 번역 수준 모두 증가 하였으며, Bad 역시 번역 수준에서 대조군에 비하여 증가되었음을 알 수 있었다. 반면 Bcl-2 는 전사 및 번역 수준에서 모두 감소되었으며, Bcl-xL의 발현은 번역 수준에서 감소하였지만, 전사 수준에서는 큰 변화를 관찰 할 수 없었다. 다음으로 조사된 death
Fig. 5. Effects of MEBJ treatment on the levels of apoptosis-related gene products in human hepatocarcinoma Hep3B cells. (A) After 72 h incubation with MEBJ, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with indicated primers and the reaction products were subjected to electrophoresis in 1.0% agarose gels and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells grown under same condition as (A) were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
receptor pathway에 관여하는 인자 중 Fas의 발현이 MEBJ 처리 농도의 증가에 따라 매우 증가되었으나 나머지 인 자들[Fas legand (FasL), death receptor (DR)4 및 DR5]은 큰 변화가 없었다. 한편 apoptosis에 관여하는 여러 인자들 중 최근 밝혀진 또 다른 class인 inhibitor of apoptosis proteins (IAPs) family에 속하는 여러 유전자 산물들 중 어 떤 IAPs는 caspase와의 직․간접적인 결합을 통하여 그들의 apoptotic 효소 활성을 억제함으로서 apoptosis 유발을 막 을 수 있는 것으로 밝혀져 있다. 조사된 IAPs family 인자 중 XIAP는 큰 변화가 없었으나, cIAP-1 및 cIAP-2의 경우 는 MEBJ 처리에 따라 발현양이 다소 감소되었음을 알 수 있었다. 따라서 MEBJ 처리에 의한 변화된 이들 인자
들은 MEBJ에 의한 apoptosis 유도에 어느 정도 관련되어 있음을 나타내어 주는 결과이다.
5. Caspase의 발현 및 활성에 미치는 MEBJ의 영향
Caspase 군에 속하는 여러 효소들 역시 apoptosis 유발에 중요한 조절인자로서 apoptosis 유도를 활성화 시키는 신 호에 의해 활성화되면 세포 내에 존재하는 많은 표적 단 백질의 분해에 관여한다. 본 연구에서는 MEBJ에 의한 이들 효소들의 관련성 여부를 조사하기 위하여 현재까 지 알려진 caspase 중 apoptosis 유도 경로에 핵심적인 역 할을 하는 caspase-3, -8 및 -9의 발현 및 활성에 미치는 MEBJ의 영향을 조사하였다. Fig. 6A의 결과에서 볼 수
Fig. 6. Activation of caspases and degradation of PARP protein by MEBJ treatment in human hepatocarcinoma Hep3B cells.
(A) After 72 h incubation with MEBJ, the cells were lysed and then cellular proteins were separated by SDS- polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) After 72 h incubation with MEBJ, aliquots were incubated with substrates, DEVD-pNA, IETD-pNA and LEHD-pNA, for in vitro caspase-3, -8 and -9 activity, respectively, at 37oC for 1 h.
The released fluorescent products were measured. Data are means±SD of three independent experiments.
있듯이 MEBJ의 처리에 의하여 불활성형인 pro-caspase-3 의 발현에는 큰 변화가 없었으며 active-caspase-3의 발현 은 MEBJ 처리에 농도 따라 발현의 정도가 점차 증가하 였음을 알 수 있었다. 그러나 caspase-8 및 -9의 경우는 MEBJ 처리에 따른 큰 변화를 보여주지 못하였다. 따라 서 in vitro caspase activity assay를 통하여 caspase의 활성 정 도를 직접 분석한 결과, Fig. 6B에 나타난 바와 같이 MEBJ가 처리된 배지에서 배양된 Hep3B 세포에서는 처 리 농도 의존적으로 caspase-3의 활성이 매우 증가되어 1.0 mg/ml 처리군에서는 대조군에 비하여 약 4.7배 증가 되었다. 그러나 caspase-8 및 -9의 경우 최고 1.4∼1.5배 수 준으로 증가되어 Western blotting 분석에 의한 결과와 유 사하게 유의적인 활성의 증가를 관찰할 수 없었다. 한편 DNA repair나 genomic stability의 유지에 매우 중요한 역 할을 하는 poly (ADP-ribose) polymerase (PARP) 단백질은 caspase-3의 활성화와 연관된 apoptosis가 유발되었을 경우 caspase-3에 의하여 특이하게 분해가 일어나는 가장 대표 적인 apoptosis 표적 단백질이다. 따라서 MEBJ의 처리에 의한 Hep3B 세포의 apoptosis에 caspase-3의 활성화가 직접 관여하는지의 여부를 조사하기 위하여 PARP의 발현에 미치는 MEBJ의 영향을 조사한 결과는 Fig. 6A에 나타낸 바와 같다. 제시된 결과에서 알 수 있듯이 MEBJ의 처리 농도 증가에 따라 PARP 단백질의 단편화 정도가 점차 증가함을 알 수 있었으며, 이는 MEBJ 처리에 따른 apo- ptosis 유도에 caspase-3이 매우 중요한 역할을 하고 있음
을 보여주는 결과이다.
고 찰
본 연구에서는 인체 간암세포주인 HepG2 및 Hep3B 세포를 대상으로 부자 추출물의 항암기전에 관한 연구 를 실시하였다. 이를 위하여 선행연구에서 특히 항암활 성이 높았던 부자의 methanol 추출물(MEBJ)을 대상으로 apoptosis 유도 활성을 비교하였으며, 그와 연관된 유전자 들의 발현 변화를 조사하였다. 본 연구의 결과에 의하면 MEBJ는 HepG2 세포보다 Hep3B 세포에서 증식억제 및 생존율 저하에 대한 감수성이 높게 나타났다. 이러한 암 세포 증식억제 및 생존율 감소가 apoptosis 유발과 밀접한 연관성이 있을 것으로 기대되어 이에 대한 증거를 제시 하기 위하여 암세포의 핵 형태 변화를 관찰한 결과, Fig.
3에 나타난 바와 같이 정상 배지에서 자란 세포들은 핵 의 전체가 완전한 형태로 염색되는 양상을 보였으나, MEBJ을 처리한 경우 처리 농도가 증가할수록 apoptosis 유발 특이적인 핵 내 DNA 단편화에 의한 염색질의 응축 된 형태인 apoptotic body의 형성 정도가 증가되었다.15,16) 이는 MEBJ의 처리에 따른 간암세포의 증식 억제 및 생 존율 감소와 연관된 세포독성의 증가는 apoptosis 유발과 밀접한 연관성이 있음을 보여주는 결과로 생각된다. 아 울러 Fig. 4B에 나타난 바와 같이 flow cytometry 분석에 의한 apoptosis 유발 세포군에 해당하는 sub-G1기에 속하
는 세포들의 빈도 역시 MEBJ 처리 농도 의존적으로 증 가되어 MEBJ 처리에 따른 암세포의 증식 억제는 apop- tosis 유발과 직접적인 연관이 있음을 알 수 있었다.
Apoptosis를 유발하는 경로는 크게 death receptor path- way와 mitochondrial pathway의 두 가지로 구분할 수 있는 데, death receptor는 세포사멸의 기능을 갖는 tumor nec- rosis factor (TNF) 수용체군을 의미한다.17) 그중 현재 가장 잘 알려진 다섯 종류 중 하나가 APO-1 또는 CD95라고도 알려진 Fas에 의한 신호전달계이며, TNF 수용체군에 작 용하는 대표적인 리간드(ligand) 중의 하나가 FasL이 다.18,19) FasL는 type II transmembrane protein으로서 TNF receptor superfamily인 Fas에 결합하여 apoptosis와 연관된 immune homeostasis에 중요한 역할을 한다. 한편 TNF- related apoptosis inducing ligand (TRAIL 또는 Apo2L)는 10 여 년 전에 최초로 발견되어졌으며, TNF superfamily에 속 한다. TRAIL은 모두 다섯 종류의 ligand와 결합을 하는데 정상세포의 경우는 osteoprotegerin, DcR1 (TRAIL-R3) 및 DcR2 (TRAIL-R4)와 같은 세 종류의 decoy receptor와 결합 을 함으로서 apoptosis를 일으키지 못하지만 암세포에서 는 DR4 (TRAIL-R1) 및 DR5 (TRAIL-R2)와 결합을 하여 apoptosis를 일으키는 것으로 알려져 있다.20,21) TRAIL이 DR4 및 DR5와 결합을 하면 procaspase-8과 Fas associated death domain (FADD)을 포함하는 death inducing signaling complex (DISC)를 형성함으로서 apoptosis를 유도한다.22) Fig. 5의 결과에서 볼 수 있듯이 FasL, DR4 및 DR5는 MEBJ 처리 여부에 관계없이 큰 변화가 관찰되지 않았지 만, Fas의 경우 전사 및 번역 수준에서 모두 MEBJ 처리에 의하여 매우 증가되었다. 이러한 결과는 MEBJ에 의한 apoptosis 유도에 Fas/FasL system이 관여할 가능성을 보여 주는 것으로 이에 관한 추가적인 연구가 요구되어 진다.
그리고 Bcl-2 family에 속하는 몇 가지 중요한 인자들은 apoptosis 유발 조절에 가장 대표적인 유전자로 알려져 있 는데, 그 중 Bcl-2나 Bcl-xL은 anti-apoptotic 분자로서 apo- ptosis의 유발을 억제하는 기능을 가지며, Bax나 Bad 등은 pro-apoptotic 분자로 그들의 발현 증가는 apoptosis의 유발 과 관계가 있다.3,23) 이들 두 유전자는 세포 내 소기관 중 mitochondria로부터의 cytochrome c를 유리시켜 cysteine- related protease인 caspase, 및 DNA의 단편화와 연관된 endonuclease 등의 활성을 조절하며,4,24) 서로 dimer의 형태 로 존재하지만 그들의 발현 수준에 변화가 초래되면 apoptosis가 유발되는 것으로 알려져 있다.5,25) 본 연구의 결과에 의하면, Bax 유전자의 발현이 MEBJ 처리에 의하 여 전사와 번역 수준에서 모두 증가되었으며, Bad 단백 질의 발현 역시 MEBJ 처리에 의하여 대조군에 비하여
증가되었음을 알 수 있었다. 반면 Bcl-2와 Bcl-xL의 발현 은 전체적으로 MEBJ 처리에 의하여 감소되는 경향성을 보여 주었다. 이는 결국 MEBJ 처리에 의한 apoptosis이 유 발에는 최소한 Bcl-2 family가 중요한 역할을 하고 있음을 의미하는 것이며, anti-apoptotic 인자들에 비하여 pro- apoptotic Bax 및 Bad의 상대적 발현 증가로 인한 apoptosis 유발 관련 효소들의 활성화가 이루어지고 있음을 시사 하여 주는 것이다. 그리고 IAPs family에 속하는 여러 인 자들은 곤충세포에서 바이러스 감염에 의한 apoptosis를 억제하기 위한 baculoviral 단백질군들로서 외부 신호에 의한 세포 내 anti-apoptotic 활성을 지닌다. 그리고 이들 중 어떤 IAPs 인자는 caspase와의 직접적인 결합을 통하 여 그들의 apoptotic 활성을 억제할 수 있을 것으로 밝혀 져, IAPs family는 Bcl-2 family와 함께 caspase 활성 조절을 통한 apoptosis 유도 신호전달에 결정적인 역할을 하는 것 으로 알려져 있다.8,26,27) 따라서 MEBJ 처리에 의한 apop- tosis 유발에 IAPs family가 관여하는지의 여부를 조사한 결과, Fig. 5에 나타낸 바와 같이 MEBJ의 처리에 따라 XIAP는 큰 변화가 없었으나, cIAP-1 및 cIAP-2의 발현은 감소되어 caspase의 활성을 증가시켜 줄 가능성을 높음을 보여주었다.
대표적인 apoptosis 유발 protease인 caspase family에 속하 는 단백질들은 세포에서 핵과 mitochondria의 외막에 불 활성 상태인 proenzyme 형태로 존재하다가 Bcl-2 및 IAPs family 인자들의 발현 변화와 연관된 apoptosis 유도를 활 성화시키는 신호에 의해 활성화된 protease로 전환되어 직접 또는 간접적으로 세포 내에 존재하는 많은 표적 단 백질의 분해에 관여하는 것으로 알려져 있다.6,28,29) 따라 서 caspase의 활성화 자체가 apoptosis의 유발에 대한 또 다른 증거가 될 수 있다. 따라서 지금까지 알려진 caspase 중 대표적인 caspase-3, -8 및 -9의 발현 및 활성에 미치는 MEBJ의 영향을 조사한 결과, Fig. 6에서 알 수 있듯이 조 사된 caspase 중 caspase-3의 활성이 가장 많이 증가되었 다. Apoptosis가 유발되었을 경우 특이하게 분해가 일어 나는 몇 가지 표적 단백질 중 PARP 단백질은 DNA repair 나 genomic stability의 유지에 매우 중요한 역할을 하는 것으로 알려져 있으며, 활성화된 caspase-3에 의하여 단백 질의 분해가 일어나면 PARP 기능이 상실되며, 정상적인 세포의 경우 PARP 단백질은 116 kDa의 분자량을 가지지 만 apoptosis가 일어난 경우 85 kDa 크기의 단편이 관찰되 거나 주 band의 발현이 감소된다.6,7) Fig. 6A의 결과에서 알 수 있듯이 PARP의 발현은 caspase-3의 활성 증가와 연 관되어 MEBJ의 처리 농도 의존적으로 단편화 현상이 뚜 렷하게 관찰되어 MEBJ 처리에 의한 apoptosis 유도에
caspase-3이 중요하게 관여함을 알 수 있었다.
이상의 결과에서 MEBJ 처리에 의한 간암세포에서의 apoptosis 유발은 Fas의 발현 증가, Bcl-2 및 IAPs family에 속하는 인자들의 발현 변화와 그에 따른 caspase의 활성 화와 연관된 caspase 기질 단백질의 분해에 의해서 일어 남을 알 수 있었다. 비록 본 연구의 결과에서 caspase-8의 활성이 크게 증가하지는 않았지만, MEBJ에 의한 간암세 포의 apoptosis 유발에는 death receptor 및 mitochondria pathway를 통하는 유전자들의 발현에 전반적으로 영향 을 미쳤음을 의미하는 것으로 추정된다.
결 론
본 연구에서는 HepG2 및 Hep3B 인체 간암세포를 이 용하여 부자 methanol 추출물(MEBJ)의 항암기전 해석의 일환으로 apoptosis 조절에 관여하는 몇 가지 주요 인자들 의 발현 변화를 조사하였다. MEBJ는 HepG2 세포에서보 다 Hep3B 세포의 증식을 효과적으로 억제하였으며, 생 존율의 저하 및 apoptosis 유도 활성에서도 Hep3B 세포에 서 높은 감수성을 보여 주었다. MEBJ에 의한 apoptosis 유 도의 여부는 apoptosic body의 형성 및 sub-G1기에 속하는 세포 빈도의 증가로 확인하였으며, Hep3B 세포에서 MEBJ에 의한 apoptosis 유도에는 Bcl-2 family에 속하는 pro-apoptotic Bax의 전사 및 번역수준에서의 증가 및 anti-apoptotic Bcl-2 및 Bcl-xL의 발현감소와 Fas의 발현 증 가 및 IAPs family 인자들의 발현 감소와 연관성이 있었 다. MEBJ 처리는 또한 caspase-3의 활성화에 따른 PARP 단백질의 단편화를 유발시켰다. 비록 본 연구의 결과로 부자 추출물의 항암기전을 논하기는 어렵지만, 부자 추 출물의 생화학적 항암기전 해석을 이해하고 향후 수행 될 추가 실험을 위한 기초 자료로서 그 가치가 매우 높 을 것으로 생각된다.
참 고 문 헌
1) Barisic K, Petrik J, Rumora L. Biochemistry of apoptotic cell death. Acta Pharm 53, 151-164, 2003.
2) Fulda S, Debatin KM. Apoptosis signaling in tumor therapy.
Ann NY Acad Sci 1028, 150-156, 2004.
3) Antonsson B, Martinou JC. The Bcl-2 protein family. Exp Cell Res 256, 50-57, 2000.
4) Fan TJ, Han LH, Cong RS, Liang J. Caspase family proteases and apoptosis. Acta Biochim Biophys Sin 37, 719-727, 2005.
5) Rupinder SK, Gurpreet AK, Manjeet S. Cell suicide and caspases. Vascul Pharmacol 46, 383-393, 2007.
6) Kaufmann SH, Desnoyers S, Ottaviano Y, Davidson NE,
Poirier GG. Specific proteolytic cleavage of poly(ADP-ribose) polymerase: an early marker of chemotherapy-induced apoptosis. Cancer Res 53, 3976-3985, 1993.
7) Lazebnik YA, Kaufmann SH, Desnoyers S, Poirier GG, Earnshaw WC. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 371, 346-347, 1994.
8) Salvesen GS, Duckett CS. IAP proteins: blocking the road to death's door. Nat Rev Mol Cell Biol 3, 401-410, 2002.
9) 楊東喜. 本草備要解析. 新竹. 서울, 國興出版社, pp 226-233, 1985.
10) 辛民敎. 臨床本草學. 서울, 永林社, pp 263-264, 1994.
11) 한종현, 김기영. 한방약리학. 서울, 의성당, pp 271-277, 2004.
12) 박영순. 한방의 약리해설. 서울, 아카데미서적, pp 136- 137, 2002.
13) Nemoto Y, Satoh K, Toriizuka K, Hirai Y, Tobe T, Sakagami H, Nakashima H, Ida Y. Cytotoxic and radical scavenging activity of blended herbal extracts. In Vivo 16, 327-332, 2002.
14) Kim JH, Choi YH, Hwang WD. Inhibition of cell proliferation and induction of apoptosis by methanolic extract of Aconiti Tuber in human renal cell carcinoma cells. Korean J Orietal Int Med 29, in press, 2008.
15) Evans VG. Multiple pathways to apoptosis. Cell Biol Int 17, 461-476, 1993.
16) Shi L, Nishioka WK, Th'ng J, Bradbury EM, Litchfield DW, Greenberg AH. Premature p34cdc2 activation required for apoptosis. Science 263, 1143-1145, 1994.
17) Sheikh MS, Huang Y. Death receptors as targets of cancer therapeutics. Curr Cancer Drug Targets 4, 97-104, 2004.
18) Klas C, Debatin KM, Jonker RR, Krammer PH. Activation interferes with the APO-1 pathway in mature human T cells.
Int Immunol 5, 625-630, 1993.
19) Daniel PT, Krammer PH. Activation induces sensitivity toward APO-1 (CD95)-mediated apoptosis in human B cells.
J Immunol 152, 5624-5632, 1994.
20) Ashkenazi A, Dixit VM. Death receptors: signaling and modulation. Science 281, 1305-1308, 1998.
21) Kimberley FC, Screaton GR. Following a TRAIL: update on a ligand and its five receptors. Cell Res 14, 359-372, 2004.
22) Tibbetts MD, Zheng L, Lenardo MJ. The death effector domain protein family: regulators of cellular homeostasis. Nat Immunol 4, 404-409, 2003.
23) Osford SM, Dallman CL, Johnson PW, Ganesan A, Packham G. Current strategies to target the anti-apoptotic Bcl-2 protein in cancer cells. Curr Med Chem 11, 1031-1039, 2004.
24) Lowe SW, Ruley HE, Jacks T, Housman DE. p53-dependent apoptosis modulates the cytotoxicity of anticancer agents. Cell 74, 957-967, 1993.
25) Donovan M, Cotter TG. Control of mitochondrial integrity by Bcl-2 family members and caspase-independent cell death.
Biochim Biophys Acta 1644, 133-147, 2004.
26) Holcik M, Gibson H, Korneluk RG. XIAP: apoptotic brake
and promising therapeutic target. Apoptosis 6, 253-261, 2001.
27) Cheng JQ, Jiang X, Fraser M, Li M, Dan HC, Sun M, Tsang BK. Role of X-linked inhibitor of apoptosis protein in chemoresistance in ovarian cancer: possible involvement of the phosphoinositide-3 kinase/Akt pathway. Drug Resist Updat 5, 131-146, 2000.
28) Rao L, White E. Bcl-2 and the ICE family of apoptotic regulators: making a connection. Curr Opin Genet Dev 7, 52-58, 1997.
29) Vegran F, Boidot R, Oudin C, Riedinger JM, Lizard-Nacol S.
Implication of alternative splice transcripts of caspase-3 and survivin in chemoresistance. Bull Cancer 92, 219-226, 2005.