약 학 회 지 제 39권 제 6 호 654*-660(1995) Yakhak Hoeji Vol. 39. No. 6
알록산 유도 당뇨환쥐의 폐에서 황산화계의 변화
최형호* • 고광삼 ■임동윤
조 선 대 학 교 의 과 대 학
(Received September 12. 1995)
T h e C h a n g e s o f A n t io x id a n t E n z y m e s i n th e L u n g o f A llo x a n - in d u c e d D ia b e tic R a t s
H y u n g -H o C h o i*, K w a n g - S a m K o h a n d D o n g - Y o o n L im College of Medicine, Chosun University, Kwang Ju 501-759, Korea
Abstract— The present study was attempted to investigate the mechanism of oxidative cellular injuries which occur in diabetic rats by determining changes of antioxidant enzymes activity in the lung of al
loxan-induced diabetic rats, the contents of glutathione in the lung, liver, blood samples, and y-glu- tamylcysteine synthetase activities in the liver. Superoxide dismutase activities (SOD), including Cu.
Zn-SOD and Mn-SOD. decreased in the lung of diabetic rats compared with those of normal control rats. However, activities of catalase and glutathione peroxidase (GP50 activities were not affected in the lung of diabetic rats. In diabetic rats, glutathione contents in the lung, liver, and blood samples, as well as the activities of y-glutamylcysteine synthetase in the livers which is known to be the key en
zyme of glutathione biosynthesis, decreased significantly. From these experimental results, it is thought that the decrease in SOD activities in the lung, glutathione contents and y-glutamylcysteine synthetase activities in some tissues in alloxan-induced diabetic rats may be the crucial cause of vul
nerability to oxidative cellular injuries.
Keywords □ Superoxide Dismutase, Cellular Injury. n-Induced Diabetes.
Sup ero xid e d ism utase (S O D ), ca ta la se . perox
id a se 동항산화효소와ceru lo p lasm in , tra n s fe rrin , la c to fe rrin , alb u m in , haptoglobulin 둥비효소계항 산화 단백절 및 g lu tath io n e , tocopherol, P-caro - te n e , ascorbic acid , u ric acid , b iliru b in 등비단백 성항산화물질은생체에서생성된유리기의소거에관 여하는물질이기 때문에. 계속적으로서서히 진행되는 항산화계기능외감소는점진적인세포손상을초래하여 세포노화를 촉진시키고, 동맥경화중. 퇴행성 관절염.
당뇨병. 암등의 발생에도관련이 깊은것으로 알려져 있다/" ®
Strep to zo to cin과a llo xa n은췌장의 p -ce ll을선텍
* 본 논 문 에 관 한 문 외 는 이 저 자 에 게 로
062-220-3659 (팩 ^ ) 062-227-4693
적으로파괴시켜당뇨를유발하는데. a llo xa n의득성기 전은superoxide ra d ic a ls의생성으로인한세포손상 과밀접한 관련이 있으며, 또한free ra d ic a ls에의한 세포손상은당뇨병외발생과당뇨합병중. 즉혈관장애.
망막중 (re tin o p a th y ). 신중 (n ep h ro p ath y) 등의발
생과깊은연관이있는것으로알려져왔다.®"® stre p -
tozotocin 유발 당뇨 흰쥐의 적혈구 glutathio ne peroxidase (G P X ) 활성도는 중가되고. 간 g lu- tathio n e 량은감소되며 , 간, 신장및적혈구에서copp- er and zinc-containing superoxide dism utase (C u . Zn -SO D ) 활성도가 감소되고, m anganese contain
in g superoxide dism utase (M n -SO D ) 활성도는 중 가되어있으며 , 췌장에서ca ta la se 와C u ,Z n -S O D 활 성이유의하게중가된다는보고가 있 다 . " 그 러 나 현 재까지 당뇨모델에서 폐를 이용한항산화효소에 대한
654
알록산유도당뇨흰쥐의 폐에서항산화계의변화
연구는 알려진 바가 많지 않다.
당뇨병 환자에서 적혈구 SO D 활성도가 증가하는 반면 G P X 활성도 번화는 없으나, 소아 인슬린 외존성 당뇨병 환자(ID D M )에서 적혈구 C u ,Z n -S O D 및 G P X 활성 이 감소하는 것으로 보고된 바 있으나*"*, 당 뇨병환자외 적혈구 g lu ta th io n e 량 및 G P X 활성도가 감소되었고 다형핵 백혈구(p o lym o rp h o nu clear le - u ko cyte s) 의 su p e ro xid e an io n 생성량이 중가되어 있고 C u ,Z n -S O D 및 M n-SO D 활성도가 유의하게 감소된다는 보고도 있어 당뇨병 환자의 항산화계는 장 기에 따라 번화외 유형이 서로 다르게 나타남을 알 수 있다.*®^*® 비효소계 항산화 물질인 g lu tath io n e은 g lu ta m y lcy ste in e sy n th e ta se 와 g lu ta th io n e s y - n th e ta se 에 의헤서 g lu ta m ic a icd , cyste in e 및 g- ly cin e 으로부터 합성되는 trip e p tid e 로서, 여러가지 해득반응, 단백질이나 D N A 의 합성, am ino a cid 의 이동반응 및 th io l기의 저장등 생물학적으로 중요한 여 러가지 반응에 직접 관여한다고 한다,*®* 본 실험에서 는 allo xa n 을 투여 하여 당뇨병 이 유발된 흰쥐 의 폐에 서 c a ta la se , SO D 및 G P X 활성도와 g lu tath io n e 량을 측정하여 항산화계의 변화롤 관찰하고, g lu ta th io n e 및 y g lu ta m y lc y s te in e sy n th e ta se 활성을 측정하여, 당뇨병에서 비효소계 항산화 물질인 g lu - ta th io n e 량외 변화에 머처는 ■ yglutam ylcysteine sy n th e ta se 의 영향을 관찰하고자 하였다.
실험 방법
실험동물의 처치- 실험동물은 S p rag u e-D aw ley종 수컷 흰쥐로, 정상 대조군과 allo xan 투여 당뇨군으로 나누어, 당뇨군은 allo xan (50 m g/kg)을 2일 간격으 로 3회 복강내로 주사하여 당뇨병을 유발시키고 3주 후 에 혈당을 측정하여 혈당량 180m g/dl blood 이상인 흰쥐를 당뇨군으로 분류하고, 간과 폐를 적출하여 d ry ice-eth ano l 용액에 즉시 냉동시키고, 정상대조군도 같 은 방법으로 간과 폐를 처치하여. -80°C 냉동고에 보존 하였다.
조효소액의조제 - 적출된 간과 페조직 일정 량을 미 리 냉각된 10배 (W /V ) 외 100 m M T ris - H C l b u ff
e r (p H 7 .4 )를 가하여 , p o lytro n hom ogenizer로 균 질화시키고, 4°C 에서 10,00 0X 담로 30분간 원심분리하 여 상층액을 효소활성 측정의 조효소액으로 사용하였
다 .
또한 간과 폐조직에 동량 (W /V )외 10 % (W /V ) su lfo sa lic y lic a d d (S S A )를 가하고, 조직을 마쇄시켜 lO.OOOxg로 4°C 에서 15 분간 원심분리하여 상등액을 g lu tath io n e 정량 시료로 사용하였다. 하대정맥으로부 터 혈액을 채취하여 동량의 20% (V /V ) S S A 룰 혼합하 여 lO.OOOxg로 4°C에서 15분간 원심분리하여 상등액 을 혈액 g lutathio ne 정량 시료로 사용하였다.
항산화효소활성도측정 - S O D 활성도는 C rap o 등 의 방법*'"으로 cytochrom e C 를 이용해서 측정하여 cytochrom e C 의 환원 속도를 50% 억제하는 효소의 양을 1 u n it로 표시하였다. M n -S O D 활성도는 효소반 응액에 K C N 을 최종농도가 2 m M 이 되게 가하여 상기 와 같은 방범 으로 측정 하였다.
C a ta la se 활성도는 A e b i의 방법^®에 따라 H2O2량
을 측정하였고, 효소의 활성도는 1분 동안에 1 n M 의 H2O2를 분헤키는 효소의 양을 1 u n it로 하였다.
G P X 활성은 F lo h e 등외 방법®에 의해 측정했다.
즉 1 m M E D T A 를 함유한 100 m M p o tassiu m p h o sp h ate b u ffe r (p H 7 .0 ) 2 .5 m /에 3 m M G S H , 20 m M N aN a, g lu ta th io n e re d u c ta se 0.72 U , N A D P H 0.45 m M 과 조효소 용액 10 uZ를 넣고 37°C 에서 5분간 방치한 후 0 .4 5 m M H2O2를 가하여 340 n m 에서 3분 동안 흡광도의 변화를 측정하였으며. 비효소적 반응은 상기와 같은 조건에서 단지 0.45 m M H2O2를 가하지 않고 반응시켜 흡광도의 번화를 측정하였고, 효소활성도는 비 효소적 .반응에 의한 흡광 도 번화률 감한 값을 다옴식에 의 헤서 계산하였다.
A = 0 .8 6 8 ( [N A D P H ]/(G S H ]o t) ( V j/ V ,)
여기서 A : 효소 활성도, [N A D P H ]: N D A P H 농도 번화, [GSHGo: G S H 의 처옴농도. t ; 반응 시간, V i: 반 응액의 용량 및 V , ; 효소액외 용량이다.
Glutathione 정량 - G lu ta th io n e 정량은 T ie tze ®의 D T N B -G S S G red uctase recyclin g 방법에 의해 측정 하였다. 이 방법은 조직내 총 glutathio ne (G S S G + G S H )의 정량에 이용되는 에민한 방법으로 측정원리는 G S H 와 D T N B 가 반응하여 G S S G 와 5-th io -2-n i- trobenzoic acid (T N B )를 형성하고 (1 ), G S S G 와 N A D P H 가 glutathione red uctase (G R ) 의 작용으로 G S H 로 환원되는 (2) 재순환 반응 (re cyclic reactio n ) 을이용하여.
Vol. 39, No. 6, 1995
C u , Z n - S O D M n - S O D
L 니 ng
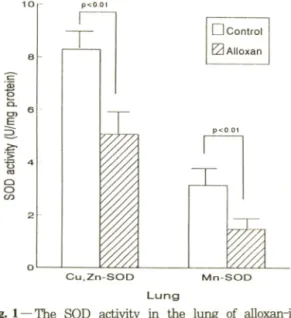
Fig. 1 — The SOD activity in the lung of alloxan-in
duced diabetic rats. Rats were treated by in
traperitoneal Injection of alloxan (50 mg/kg) 2 times every 2 days. Three weeks after the final injection, they were sacrificed by cervical dislo
cation. Cu.Zn-SOD and Mn-SOD activities were determined by Crapc/s method. Results are expressed as meanrbS.D. from 7 rats. Sta
tistical difference were calculated by comparing enzyme activities between control and alloxan- treated groups.
결 과
A llox an으로유발시킨 당뇨환쥐의 페 SO D 찰성도 변화 - A llo x a n 을 복강으로 투여하여 3주 후 당뇨가 유발된 7마러 흰쥐의 페 SO D 활성도룰 측정한 결과.
당뇨 흰쥐의 페 C u .Z n -S O D 활성도는 5 .0 8 ± 0 .9 2 U /m g p ro te in 이었고. 정상 흰쥐의 페 C u .Z n -S O D 활성도는 8 .2 9 ± 0 .7 2 U /m g p ro tein 으로, 폐 C u ,Z n - SO D 활성도는 당뇨 흰쥐에서 감소되었다(p< 0.01).
또한 당뇨 흰쥐의 페 M n-SO D 활성도는 1.50± 0.35 U /m g p ro tein 이었고. 정상 흰쥐의 페 M n-SO D 활성 도는 3 .1 5 ± 0 .4 5 U /m g protein으로. M n-SO D 활성 도는 당뇨군의 폐에서 낮은 활성도를 나타냈다(F ig .l).
A lloxan으로유발시킨 당뇨환쥐의 페 catalase 황성 도 변화- A ilo xa n 을 복강으로 투여하여 3주 후 당뇨 가 유발된 7마리 흰쥐의 폐 catalase 활성도를 측정한 결과, 당뇨 흰쥐의 페 catalase 활성도는 89.72± 6.29 m U /m g p ro tein 이었고, 정상 흰쥐의 페 catalase 활 성도는 94.10± 7.33m U /m g protein으로. 폐 cata-
Pharm. Soc. Korea T N B 생성량이 G S S G + G S H 량과 비례하므로
T N B 링을 405 nm 에서 측정하여 계산 하였다. O xid i*
zed g lu tath io n e (G S S G )은.반응식 (2 )를 이용하여 N A D P H 가 산화되는 정도를 측정하였다. 즉. 100 m M p o tassiu m phosphate b u ffer. p H 7 .0 , 1 m M E D T A . 50 uM N A D P H 와 sam ple 100 u /를 25°C로 한 다옴. glutathio ne red uctase를 0 .2 U /m /되게 하 고 340 nm 에서 흡광도의 변화를 측정하여. G S S G sta n d a rd 룰 ca lib ra tio n 한 다옴 계산하였다.
y-Glutam ylcysteine synthetase 황성도 측정 -
y-
G lu ta m y lc y ste in e sy n th e ta se 활성도는 M e iste r 등21>의 방법에 의해서 측정하였다. 측정방법의 원리는 g lu ta m in e 과 c y ste in e 이 Y~ g lutam ylcysteine s y - n th e ta se 의 작용으로 A T P 를 소모 하면서 y -g lu ta - m y lc y ste in e 을 함성한다. (3 ) 이때 생성된 A D P 는 p h o sp h o e n o lp yru vate 가 p y ru v a te k in a s e 에 의해 서 p y ru v a te 로 전환 될때 A D P 가 A T P 로 전환되고
(4 ). p y ru v a te 는 L D H 에 의해서 N A D H 를 N A E T 로 산화시키면서 la c tic acid 로 전환된다. (5 ) 따라서 N A D H 가 N A iy 로 산화된 량을 자외선 분광흡광 광 도계를 이용하여 3 4 0 n m 에서 놓광도를 측정하여 계산 하였다.
g lu tam in e + g lycin e+ A T P --- > y-g lu -cys+ A D P (3) y-Glutamylcysteine synthetase phosphoenol p yru vate + A D P —► pyruvate+ A T P (4)
pyruvate kinase
P y ru v a te + N A D H ---> la ctic acid + N A D ^ (5) LDH
효소활성도는 N A D H 흡광계수 6.22/m M /Cm 을 이 용하여 단위 시간당 산화되는 N A D H 의 나m ole 수로 표시하였다.
단백질 정량 - 단백질의 정 량은 Lo w ry 등"®외 방범 에 의해서 6 0 0 n m 에서 흡광도를 측정하고 표준 단백 시료를 같은 방법으로 측정하여 단백질량을 계산하였 다.
이상의 실험결과의 유의성 검정은 Stu d en t/s t- te st를 이용하여 상호비교, 검정하였다.
최형호■고광삼■임동윤
2 G S H + 2 D T N B ^ G S S G + 2 T N B (1) G S S G + N A D P H + 2 G S H + N AD P^ (2)
GR
CH Control Q Alloxan
64 2
{uiBOJd 6E/n
l
aos
알록산유도 당뇨흰쥐의 폐에서 항산화계의 번화
Table I ― The activity of catalase in the lung of al
loxan-treated rats
Table i n — The glutathione levels in the livers of al-
Enzyme activity (mU/mg protein)
Control Alloxan
Catalase 94.10±7.33 89.72±6.29
Catalase activity was determined by Abei's method.
The other methods and legends are the same as in Fig. 1. Values are mean±S.D., n =7.
Table II — The activity of GPX in the lung of alloxan- treated rats
Enzyme activity (mU/mg protein)
Control Alloxan
CPX 116.87+8.51 124.36±6.83
CPX (glutathione peroxidase) activity was determined by Flohe's method, others are the same as Fig. 1.
Values are mean±S.D., n=7.
la se 활성도는 당뇨군에서 약간 감소하였으나 두 비교군 사이에 유의한 차이를 나타내지는 않았다(Tab le I) .
A llo xan 으로 유발시킨 당뇨 흰쥐의 페 G P X 활성도 변화 - A llo xa n 을 복강으로 투여하여 3주 후 당뇨가 유발된 7마리 흰쥐의 폐 G P X 활성도를 측정한 결과.
당뇨 흰쥐의 폐 G P X 활성도는 124.36+ 6.83 m U / mg p ro tein 이었고, 정상 흰쥐의 폐 G P X 활성도는 116.87± 8.51 m U /m g p ro tein 으로, 페 GP5C 활성도 는 당뇨군에서 약간 중가하였으나 두 비교군 사이에 유 의한 차이를 나타내지는 않았다(Tab le II) .
A llo xan 으로유발시킨당뇨 S 쥐에서간, 페, 및혈액 의 glutathione 량 변화 - A llo xa n 을 복강으로 투여하 여 3주 후, 당뇨가 유발된 흰쥐외 간. 폐. 및 혈액에서 g lu ta th io n e 량을 측정한 결과, 당뇨 흰쥐 간의 총 g lu - tath io n e^ 몬 15.98+ 2.55 나m ole/g tissu e 로 정상 흰 쥐 간의 총 g lutathio ne량 24.94± 3.75 |im ole/g tis - su e 에 비하여 약 35.9% 가 감소되었고. 당뇨군의 산화 g lu tath io n e 량은 2 .6 6 + 0 .3 4 나m ole/g tissu e 로 정상 대조군의 산화 g lu tath io n e량 4.37± 0.47 |im ole/g tissu e 에 비하여 약 39.1% 가 감소 되었으나. 총 g lu - tath io n e 에 대한 산화 g lu tath io n e 량의 비율은 거의
번화가 없었다(Tab le I II ) .
당뇨 흰쥐 폐의 총 g lu tath io n e량은 3.37± 1.02
|xm ole/g tissu e c로 정상 흰쥐 페의 총 g lu tath io n e량 5.19± 0 .63 fim ole/g tissu e 에 비아여 약 35.1% 가 감 소되었고, 당뇨군의 산화 g lu tath io n e량은 0.32+ 0 .06
|im ole/g tissu e 로 정상 대조군의 산화 g lu tath io n e 량
loxan-treated rats
Groups
Glutathione contents(^mole/g tissue)
,GSSG/(GSH+
G SH +G SSG GSSG GSSG) (%)
Control Alloxan
24.94±3.75 4.37±0.47 15.98±2.55* 2.66±0.34*
17.5 16.7 The amount of glutathione was determined by Tietze^s method. Values are mean+S.D.. n = 7
*: P<0.01 vs control
Table IV —— The glutathione levels in the lung of al
loxan-treated rats
Groups
Glutathione contents(^lmole/g tissue)
.GSSG/(GSH +
G SH +G SSG G SSG GSSG) i%)
Control Alloxan
5.19±0.63 0.16±0.05 3.37±1.02* 0.32±0.06*
3.1 9.5 The amount of glutathione was determined by Tietze^s method. Values are mean±S.D., n = 7
*: P<0.01 vs control
Table V -— The glutathione levels in each of the blood samples from alloxan-treated rats
Groups
Glutathione contents(나moie/g tissue)
.G SSG AG SH +
G SH +G SSG GSSG GSSG) {%)
Control Alloxan
5.15± 0.90 0.26+ 0.01 2.98±0.37* 0.33±0.10*
5.1 11.1 The amount of glutathione was determined by Tietze's method. Values are mean±S.D., n = 7
* ; P<0.01 vs control
0 .1 6 ± 0 .0 5 M-mole/g tissu e 에 비하여 약 2배가 증가 되었으며, 당뇨군에서 총 g lu ta th io n e 에 대한 산화 g lu ta th io n e 량의 비율은 정상 대조군에 비해서 3.1 배 가 중가되었다 (T a b le IV ).
당뇨 흰쥐 혈액의 총 g lu ta th io n e 량은 2 .9 8 ± 0 .3 7
|im o le /m / blood로 정상 대조군 혈액의 총 g lu ta t- h io n e 량 5 .1 5 ± 0 .9 0 |xm o le/m i blood에 비하여 약 4 2 .1 % 가 감소되었고, 당뇨군의 산화 g lu ta th io n e 량 은 0.3 3 ± 0 .1 (H iin o le/m Z blood로 정상 대조군의 0 .2 6 ± 0 .0 1 |iTnole/m / blood에 비하여 약 2 6 .9 % 가 중가 되 었으며 . 당뇨군의 총 g lu ta th io n e 에 대한 산화 g lu ta th io n e 량의 비율온 정상 대조군에 비해서 약 2 .2 배가 중가되 었다 (T a b le V ) .
AUoxan으로 유발시킨 당뇨 환쥐에서 간의 ^giu- tam ylcysteine synthetase 황성도 변화 - A llo x a n을복 강으로 투여하여 3주 후 당뇨기■ 유발된 흰쥐의 간 g lu ta m ylcyste in e sy n th e ta se 활성도를 죽정한 결 과 . 2 8 .4 6 ± 2 .3 0 U /m g p ro tein 이었고, 정상 흰쥐의 간
Vol. 39, No. e, 1995
Control Alloxan-treated
Fig. 2 — The activity of y-glutamylcysteine synthetase in the liver of alloxan diabetic rats. Values are mean±S.D., n=7.
Y -g lu tam ylcyste in e syn th e tase 활성도는 35.19±
3.62 U /m g p ro te in 이어서, 효소 활성도는 대조군에 비하여 당뇨군에서 약 19% 가 감소된 효소 활성도를 나 타냈다(F ig . 2 ).
고 찰
유러산소는 S O D 에 의해 H2O2와 02로. H2O2는 c a ta la se 와 G P X 에 의해 HgO로 전환되므로 S O D . c a ta la se 및 G P X 는 유러산소의 득성으로부터 세포률 보호하는 매우 중요한 효소이며. 이둘 효소의 활성이 감소되면 산소유리기에 의한 D N A 나 단백질 및 지질 손상이 초래된다고 한다.*^® 사럽체막 호흡사슬에서 생성되는 su p ero xid e와 과산화수소는 산소분압이 중 가하거나. 호흡사슬 구성물이 환원상태 일때 , uncou
p le rs (pro to pho res , ionophores 및 C a *'*')등에 의해 서 생성 량이 중가된다."® 폐는 산소의 분압이 높아 폐 세포내에서 생성되는 산소유리기가 많고, 활성화된 탐 식세포(a c tiv a te d phagocytes)에서 생성되는 ra d
ic a ls 량도 많아서 폐내외 항산화계는 폐손상과 밀접한 연관이 있다.^*
본 실험에서 A llo xa n 투여로 당뇨가 유발된 흰쥐의 폐 C u , Zn -SO D 활성도는 정상 대조군에 비하여 당뇨 군에서 38.7% 가 감소된 효소 활성을 나타냈으며 , M n- SO D 활성도는 간 과 폐에서 모두 당뇨군에서 유의하게 낮은 효소 활성도률 나타냈고, 페 catalase 와 G P X 활 성도는 당뇨군과 대조군 사이에 유의한 차이를 나타내 지 않았다.
Loven등은 streptozotocin 당뇨 흰쥐외 간, 신장 및 적혈구 C u . Zn-SO D 활성도는 감소되었으나. M n- SO D 활성 도는 중가된것으로 보고하였으며 '^ W oh- aieb 등은 streptozotocin 당뇨 흰쥐의 췌장 C u , Zn- SO D 와 catalase 활성은 감소되었고, G P X 활성둑 변 화가 없었으며 , 간 C u , Zn-SO D . catalase 및 G P X 활 성이 모두 감소되었고. 심장에서 catalase 활성은 중가 되었으나 C u . Zn-SO D 와 G P X 활성 변화는 없어서.
streptozotocin 당뇨 흰쥐에서 항산화효소 활성의 변화 는 장기에따라 달랐는데®, 본 실험결과 alloxan 당뇨흰 쥐의 폐 항산화효소 활성도 번화가 항산화효소의 중류에 따라서 다르게 나타남을 알 수 있다.
G lu ta th io n e (L rY -g lu ta rn y l-L rc y ste in y lg ly c in e ) 은 세포내에 널리 분포되어 있는 trip ep tid e로서 , 세포 내 -S H (th io l group)중 많은 부분을 차지하는 물질이 며. 생리적 기능은 단백질이나 D N A 의 합성, r g lu - ta m y l am ino acid 등과 같은 물질의 이동. 효소활성의 조절 및 활성산소나 유리기에 의한 세포손상예방 등 생 물학적오로 중요한 여러반응에 직접 또는 간접적으로
관여 한다고 한다, 캔
A llo xa n 당뇨흰쥐의 간, 페 및 혈액에서 g lu tath io n e 량을 측정한 결과, 총 g lutathio ne 량은 정상 대조군에 비하여 당뇨 흰쥐의 간에서 약 36.1% . 폐에서 약 35.1% . 혈액에서 약 42.09^가 각각 감소되었다.
G lu ta th io n e 은 T -g lu tam ylcyste in e syn th e tase (8 )와 g lu tath io n e syn th e tase (9 )에 의헤서 g lu tam ic acid , cyste in e 및 g ly cin 을 기질로 이용하여 세 포내에서 합성된다.*®
G lu ta m ic acid + C y s te in e + A T P > y -G lu ta m y lcyste in e + A D P (8) y-glutamylcysteine synthetase
y -G lu ta m y lc y ste in e + G ly c in + A T P ^ G lu ta th io n e + A D P (9) Glutathione synthetase
J. Pharm. Soc. Korea 최형호•고광삼•임동윤
0000 4321lujalojc
i roui/
nl a
j!a!p
« aseJ3£UA S au!81SAOIAilnl0A
알록산유도당뇨흰쥐의폐에서항산화계의 번화
y-G lu ta m ylcyste in e syn th e ta se 활성은 생러상태 에서 G S H 에 의해서 조절되며. 생쥐나 흰쥐에 실험적
으 로 y-g lu tam ylcysteine syn th etase 활성 억제제인 bu th io n in e su lfo xim in e 을 주사하면 간. 신장 및 혈액 등에서 glutathio ne 량이 급속히 저하되므로. b u th i
o nine su lfo xim in e 은 세포내 g lu tath io n e을 저하시킬
목 적 으 로 유용하게 쓰이고 있는데. y-g lu tam ylcys- te in e syn th e tase 활성이 저하되면 간이나 혈액의 glutathio ne 량이 급격히 감소되며. 담즙이나 뇨외 G S H 량도 감소된다.^ ^' 또한 흰쥐에 sodium se - le n ite 를 투여하면 Y -g lu tam ylcyste in e syn th e tase 활성도가 중가되어 g lutathio ne 량이 중가된다는 보고 도 있다."®
본 실험 결과 allo xan 당뇨흰쥐의 간 Y~glutainy- Icystein e syn thetase 활성도는 정상흰쥐의 간에 비하 여 약 19%가 감소된것으로 나타났다. 따라서* 간과 혈액 및 폐 glutathione 량 감소는 간 7~glutam ylcysteine syn thetase 활성도가 감소됨으로 인해 간에서 g lu- tathio ne 합성 량이 저하되어 간. 혈액 및 폐 g lu - tathio ne 량이 저하된것으로 생각된다.
세포내에서 소모되는 산소의 95% 이상이 사립체에 서 이용되는데.. 이중 96~ 99% 가 cytochrom e o x- id a se 에 의해서 소모되고. 나머지 1-•년%가 su p - ero xid e나 과산화수소로 전환되는데^\ 폐는 산소분압 이 높아서 superoxide나 과산화 수소의 생성 량이 많을 것으로 사료된다. A llo xa n 당뇨백서에서 폐 SO D 활성 감소와 glutathio ne 량의 감소는 세포내에 생성된 su - p ero xid e의 제거■ 및 c a ta la se 에 비해서 K m v a lu e 가 낮은 G P X 의 활성에 필요한 glutathio ne의 감소로 인 한 과산화 수소의 소거 가 층분치 못하여. allo xan 투여 당뇨흰쥐의 폐에서 산소유리기의 세포손상에 대한 감 수성이 중가 되는 요인으로 작용할 것으로 생각된다.
감사의 말씀
본 연구는 1993년도 조선대학교 학술연구비 지원에 의해 수행되었음을 밝히는 바업니다.
문 헌
1) F rid o v ic h , I . : B io lo g ica l effe cts o f th e s u peroxide rad ical. Arch. Biochem. Biophys. 247, 1
(1986).
2) M iquel, J.- H istorical introduction to free rad ical and an tio xid an t biom edical research . In CRC handbook of free radical and antioxidants in biomedi
cine, voi I. M iquel, J . Q u in tan ilh a, A . T . and Weber. H . eds, C R C Press, Flo rida p. 3 (1989).
3) P ro c to r. P . H . : F re e ra d ic a ls an d h u m a n d ise ase . C R C handbook of free radical and an
tioxidants in biomedicine, voi I, M iquel, J . Q u in - ta n ilh a . A T . and W eber. H . eds, C R C P re ss.
Flo rid a p. 209 (1989).
4) Lip p m a n , R . D . : F re e rad ical-in d u ced lip o - peroxidation and aging. In CRC handbook of free radical and antioxidants in biomedicine, voi I , M i- quel. J . Q u in tan ilh a, A T . and W eber. H . eds.
C R C P re ss. Flo rid a p. 187 (1989).
5) H a lliw e ll, B . and G utterridge, J . M . C . : R oles of free rad icals and catalytic m etal ions in h u m an disease: A n overview . Methods enzymolgy, F le is c h e r, S . and P a ck e r, L . eds. A cad em ic P re ss, In c. N ew Y o rk 186, 1 (1990).
6) C rouch, R . K .. G andy. S . E ., K im se y. G L G a l
b raith . R . A ., G alb raith . G . M . P . and B u se , M . G . : Th e inhibitio n of isle t superoxide d ism utase by diabetogenic drugs. Diabetes30. 235 (1981).
7) Le n se n . S . and P a n te n . U. : A llo xan : h isto ry and m echanism of action. Diabetologia 31, 337 (1988).
8) S to cke r. R . and F r e i. B . : End og eno us a n tio xid an t defences in hum an blood p lasm a. In Oxidative stressSies H ed. Academ ic P re ss. N ew Y o rk p. 213 (1981).
9) Godin. D . V .. W ohaieb. S . A .. G arn ett. M . E . and Goum eniouk, A . D .: A n tio xid an t enzym e a lte ra tio n s in e xp e rim e n tal and c lin ic a l d i
abetics. Molecular and Cellular Biochemistry 84, 223 (1988).
10) M atkovics, B ., V arg a. S . I., Szabo. L . an d W i- ta s. H . : Th e effect of diabetes on the a c tiv itie s o f th e peroxide m etabo lism enzym es. Horm.
Metab. Res. 14. 77 (1982).
11) Lo ven, D . P ., Schedl. H . P ., O berley, L . W ., W ilson . H . D .. B ru ch . L . and N ieh au s. C . N .;
Superoxide dism utase a ctiv ity in the in te stin a l m ucosa of the streptozotocin-diabetic ra t. En
voi 39, No. 6, 1995
최형호•고광삼• 임동윤
docrinology 111. 737 (1982).
12) Loven, D .. Schedl, H ., W ilson. H ., Daabees, T . T .. Stegink, L . D ., D iekus, M . and O berley. L .:
E ffe ct of in su lin and oral glutathione on glu
tathione levels and superoxide dism utase ac
tiv itie s in organs of rats w ith streptozotocin-in - duced diabetes. Diabetes 35, 503 (1986).
13) B ru n o , H ., Stefan, 1. M a rk .L . and G osta H .:
C u Z n superoxide d ism utase. M n superoxide dism utase, catalase and glutathione peroxidase in lym phocyte and erythrocytes in in sulin-de
pendent diabetic children. Acta Endocrinologica 102. 235 (1983).
14) U ze l. N ., S iv a s , A .. U y s a l, M . and O z, H .;
Eryth ro cyte lipid peroxidation and glutathione peroxidase a ctivitie s in patients w ith diabetes m ellitu s. Horm. Metabol. Res. 19. 89 (1987).
15) N ath. N .. C h a ri. S . N . and R a th i, A . B .; S u peroxide dism utase in diabetic polym orphon
uclear leukocytes. Diabetes 33, 586 (1984).
16) M eister, A . and Anderson, M . E . : Glutathione, Ann. Rev. Biochem. 52, 711 (1983).
17) Crapo, C . H .. M cCord, J . M . and Frid o vich, L : Preparation and assay of superoxide dism utase.
Methods in enzymol.ed. Fle isch er. S . and Pack
e r, L . Academ ic press. New Y o rk 53, 382 (1978).
18) A e b i, H . : C a ta la se . Methods of Enzymatic Analysis, Bergm eyer. H .U . Bergm eyer. J . and G ra B l, M . eds, 3rd ed, V erlag chem ie, 2, 673 (1974).
19) Flo h e. L .. W olfgang, A . and G u nzler, W . A . : A s
sa y of glutathione peroxidase. Methods in en
zymatic analaysis, Packer. L . ed. New Y o rk. A - cadem ic Press. In c. 105. 114 (1984).
20) Tietze , F .: En zym atic method for quantitative d eterm in atio n of nanogram anounts o f total and oxidized glutathione. Anal Biochem. 27. 502 (1969).
21) T a te , S . S . and M e iste r, A . : y -g lu ta m y ltr- anspeptidase from kid ney. In : M eister A . ed.
Methods in enzymol. New Y o rk ; Academ ic press
113. 400 (1985).
22) Lo w ry, O . H ., Rosenbrough, N J .. F a rr. A . L . and R an d all, R . J . : Protein m easurem ent w ith fo lin phenol reagent, /. Biol Chem, 193, 256 (1951).
23) Bo veris, A . and Cadenas, E / Production of su peroxide radicals and hydrogen peroxide in m i
tochondria. Superoxide dismutase, vol II. Oberley LW . ed.. C R C Press. Flo rid a p. 15 (1982).
24) Frido vich, L : A ntioxidant defenses in the lung.
Ann. Rev. Physiol. 48. 693 (1986).
25) Wohaieb, S . A . and Goddin, D . V .: Alterations in free rad ical tissue-defense m echanism s in streptozotocin-induced diabetes in ra t. Effect of in su lin treatm ent. Diabetes 36, 1014 (1987).
26) G riffith , O. W ., Bridges. R . J . and M eister, A .:
Evidence th a t the gam m a-glutam yl cycle func
tions in vivo using in trace llu lar glutathione: ef
fects of am ino acids and selective inhibition of enzym es. Proc. Natl Acad. Sci. 75, 5405 (1978).
27) Seelig, G . F . and M eister. A . : Glutathione biosyn
thesis'. yg lutam ylcystein e synthetase from ra t kidney. In Methods in enzynwl, M eister. A . ed., New Yo rk. Academic press. 113. 379 (1985).
28) G riffith , O. W .. Anderson. M . E . and M eister. A j Inhibition of glutathione biosynthesis by prothion- ine sulfoxim ine (S ■긴-propyl homocysteine su l- foxim ine), a selective inhibito r of gam m a-glu- tam ylcysteine synthetase, J. Biol Chem. 254, 1205
(1979).
29) G riffith . O . W . and M eister, A . ■ Potent and specific in h ib itio n of glutathione synthesis by b u th io n in e su lfo x im in a (S -n -b u ty l hom o
cysteine su lfo xim in e), }. Biol. Chem. 254, 7558 (1979).
30) C hung, A . S . and M ain es, M . D. : E ffe ct of selenium on glutathione m etabolism . Induction of gam m a-glutam ylcysteine synthetase and glu
tathione reductase in the ra t liv e r, Biochem.
Pharmacol 30, 3217 (1981).
Pharm. Soc. Korea