Vol. 14, No. 1, May, 2006
□ 종 설 □서 론
줄기세포(stem cell)는 단 하나의 세포가 분화하 여 장기가 될 수도 있고 성체에서 손상된 세포를 대체하여, 난치성 질환의 치료 가능성을 제시하므 로서 재생(regenerative) 의학의 중심을 차지하고 있다1). 많은 관심과 연구가 진행되고 있으나 현재 까지는 다른 연구 분야들처럼 실제 임상에 적용되 기 위해서는 해결되어야 할 과제들이 많이 있다.
이에 줄기세포와 관련된 기본적인 내용과 실제 소 아신경 질환에서의 줄기세포 치료의 전망을 살펴보 고자 한다.
총 론
1. 줄기세포란?
줄기세포는 기본적으로 다음의 두 가지 특성을 가지고 있다. 첫째, 미분화 상태(undifferentiated state)에서 무한히 복제 세포 분열을 통해 증식할 수 있다(self renewal, self replication, clonogenic proliferation). 둘째, 일정한 생리적 또는 실험적 상황에서 특정 세포, 예를 들어 신경세포, 심장 세 포 또는 췌장 세포로의 분화가 가능하다1).
하지만 줄기세포의 분화능을 표현하는 용어는 구분이 필요하다. 수정된 난자는 자궁에 생착하게
줄기세포 기본과 소아신경 질환에서의 전망
인제대학교 의과대학 상계백병원 소아과, 간질센터, 고려대학교 의과대학 구로병원 소아과*
강 훈 철·은 백 린*
= Abstr act =
The Baiscs and the Prospect of Stem Cells for Pediatric Neurologic Diseases
Hoon-Chul Kang, M.D. and Baik Lin Eun, M.D.*
Department of Pediatrics, Epilepsy Center, Sanggye Paik Hospital,
Inje University College of Medicine, and Department of Pediatrics
*, Guro Hospital, Korea University College of Medicine, Seoul, Korea
Stem cells are a special kind of cells that have a unique capacity to renew themselves and to give rise to specialized cell types. Researchers have for years looked for ways to use stem cells in order to replace cells and tissues that are damaged or diseased. In 1998, for the first time, it became possible to derive this kind of pluripotent stem cells from early human embryos and grow them in culture. Nearly at the same time scientists were beginning to explore human pluripotent stem cells from fetal and adult tissues, so- called adult stem cells. This promising area of science is also leading neurologists to in- vestigate the possibility of cell-based therapies to repair damaged brain cells. Among the pediatric neurologic diseases, infarcted brain, leukodystrophy, inherited metabolic diseases and neurotransmitter diseases are the immediate candidates of the stem cell therapy.
This document covers a basic information about stem cells and a potential use of the stem cell therapy for pediatric neurologic diseases.
Key Words : Stem cells, Pediatric neurologic diseases
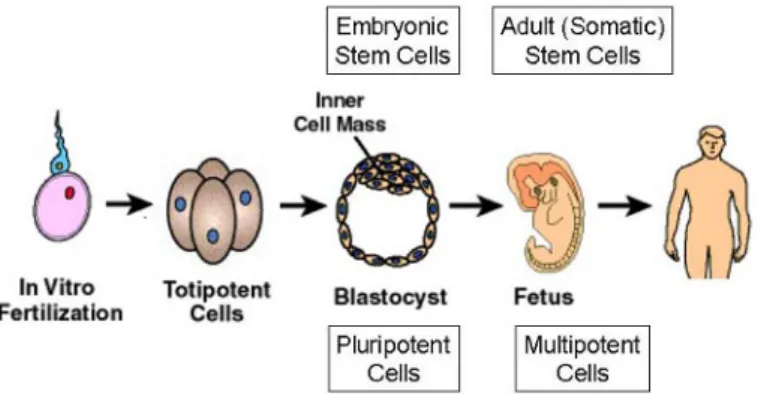
되면 배아 뿐만 아니라 배아외 조직인 태반이나 탯줄을 만들게 되고 결국 포유류에 존재하는 200 종 이상의 세포와 완성된 장기가 될 수 있다. 그러 므로 수정된 난자의 분화능은 totipotent(Toti는 라 틴어 기원으로 entire의 의미)하다고 한다. 배아줄 기세포(embryonic stem cell)는 외배엽(ectoderm), 중배엽(mesoderm), 내배엽(endoderm)에서 기원하 는 모든 종류의 세포를 만들 수 있어 pluripotent (Pluri는 라틴어 기원으로 several 또는 many의 의미)하다고 한다. 한글로는 공히 전능이라고 표현 된다. 이에 반해 정상 성체 조직 내에 존재하는 성 체줄기세포(adult stem cell)는 세포가 존재하는 조직내의 한가지 세포로만 분화될 수 있기 때문에 unipotent(Uni는 라틴어 기원으로 one의 의미)하 다고 하며, 조직이 손상되었을 경우 성체줄기세포는 pluripotent하지는 않지만 조직의 다양한 세포로 분화되어 multipotent한 능력을 보인다(Fig. 1)2).
줄기세포는 크게 나누어 배아줄기세포과 성체줄 기세포로 나눌 수 있다. 배아줄기세포는 인간 배아 줄기세포의 경우 수정 후 4-5일째 배반포(blasto- cyst) 단계에서 배반포 내 주머니배공간(blastoc- oel)의 끝에 자리잡고 있는 30-34개 정도의 세포로 구성된 속세포덩이(inner cell mass)로부터 유도 (derivation)된다. 이에 반해 성체줄기세포는 이미 분화된 조직이나 장기에 존재하면서 자가 복제 증 식이 가능하고, 일반적으로는 상황에 따라 성체줄 기세포가 존재하고 있는 조직이나 장기의 주요 세 포로 분화한다. 성체줄기세포는 탯줄 혈액(umbili-
cal cord blood) 또는 태아(fetus)에서도 분리(iso- lation)할 수 있으므로 최근에 성체줄기세포라는 용 어보다 좀 더 정확하게 체줄기세포(somatic stem cell)라고 표현하기도 한다. 이외 태아의 생식샘 능 선(gonadal ridge)에서 유도한 배아생식세포(em- bryonic germ cell)도 줄기세포의 특성을 가지고 있는데, 성체줄기세포보다는 배아줄기세포와 유사 하여 배아줄기세포에 포함시키기도 한다1).
속세포덩이를 배반포에서 제거하여 배아줄기세 포주(embryonic stem cell line)를 얻는 과정을 일 반적으로 유도(derivation) 또는 확립(establish- ment)이라는 용어로 표현하고, 성체줄기세포는 이 미 존재하고 있는 세포를 분리(isolation)한다고 한 다. 더불어 수일에 걸쳐 배아줄기세포가 증식(pro- liferation)하여 배양 접시를 가득 채우게 되면 다 른 여러 개의 배양 접시에 세포를 나누는데 이러 한 과정을 계대배양(subculturing)이라고 하고 계 대배양의 한 주기를 대이음(passage)이라고 한다.
인간 배아줄기세포의 경우 세포 이전(transfer)이 라는 과정을 통해, 쥐 배아줄기세포는 효소를 이용 한 나눔(split)이라는 과정을 통해 다음 대이음으로 계대배양하게 된다3). 한글 용어는 저자가 한글 의 학 용어 사전을 참고하여 임의로 기술한 것이므로, 괄호 안에 영문 표기를 숙지하여 줄기세포와 관련 하여 정확한 용어를 사용하여야 할 것이다(Fig. 2).
2. 배아줄기세포
쥐 배아줄기세포는 20여년 전 이미 쥐 배아에서
Fig. 1. Sorts of stem cells and differentiation potential.
확립되었다4). 쥐 배아줄기세포의 생물학적 특성에 대한 연구가 수년간 진행되면서, 1998년에는 수의 사인 James Thomson에 의해 인간 배아에서도 줄 기세포가 확립되기에 이르렀다5). 인간 배아줄기세 포의 유도는 기존의 불임 치료를 위해 시행하였던 시험관에서 수정된 난자로부터 획득하였고, 이후 공여자의 허락 하에 연구용 목적으로 제공되고 있 다. 전술한 바와 같이 배반포 내의 주머니배공간의 끝에 자리잡고 있는 30개 정도의 세포로 구성된 속세포덩이(inner cell mass)에서 유도하게 되는데, 유도 방법은 크게 3가지이고 장단점이 있다6, 7). 일 반적으로 사용하던 전통적인 방법은 면역 수술인 데, 우선 투명 구역(zona pellucida)을 효소 처리 해서 제거하고 항인간 전체혈청항체(anti-human whole serum antibody)와 기니픽(guinea pig) 보 체를 이용하여 영양외배엽(trophectoderm)을 녹인 후 속세포덩이를 분리하는 방법이다. 하지만 면역 수술은 과정이 쉽고 영양외배엽이 선택적으로 제거 될 수 있다는 장점에도 불구하고, 속세포덩이가 작 거나 형성이 엉성할 때는 분리가 제대로 되지 않
을 뿐 아니라 인간 배아줄기세포가 이종 시약으로 부터 인간 배아줄기세포가 이종 병원체에 오염될 우려가 있다. 두번째는 투명 구역만을 제거한 상태 에서 배반포 전체를 배양하는데, 속세포덩이의 상 태가 좋지 않을 때에도 사용할 수 있어 속세포덩 이의 손실이 없고 이종 시약에 의한 오염의 우려 도 없다. 하지만 영양외배엽을 같이 배양하게 되어 속세포덩이의 성장을 방해할 수 있다. 최근에 김 등8)은 부분 배아 배양법으로 전체 배아 배양법의 장점을 유지하면서 영양외배엽을 수작업으로 제거 하는 방법을 사용하고 있다. 이종 병원체에 의한 오염의 우려는 결국 인간에게 세포를 이식하고자 하는 세포 치료의 목적에 치명적인 단점이 될 수 있으므로 다소 번거롭더라도 인간 배아줄기세포의 유도는 배아 배양법이 적당하다고 생각된다.
배양 접시로 옮긴 속세포덩이는 배양 접시에서 계대배양을 하는데 세포들은 분열을 반복하면서 접 시 표면으로 퍼져나간다. 접시 표면은 미리 항분열 처리된 쥐 배아 피부 세포로 영양 세포층(feeder layer)을 만들어 두어, 속세포덩이가 표면에 잘 붙 Fig. 2. Schematic diagram of stem cell research from establishment
or isolation of stem cells to a clinical trial. Abbreviations : ES, embry- onic stem; AS, adult stem; FACS, fluorescence-activated cell sorting.
고 더불어 배양액 내로 여러 가지 영양분을 제공 한다. 하지만 최근에는 쥐 세포에서 여러 가지 바 이러스 또는 다른 거대분자들이 인간 배아줄기세포 를 오염시킬 가능성이 있기 때문에, 제라틴 겉칠이 된 접시에 외부에서 배양액을 넣어주는 방법을 사 용한다. 수일에 걸쳐 속세포덩이는 증식을 하여 배 양 접시를 가득 채우게 되고 수개월에 걸쳐 계대 배양을 한다. 6개월 이상이면 30개 정도이던 속세 포덩이 세포들이 수백만개 세포로 증식하게 되는 데, 세포들이 미분화 상태에서 전능을 보이고 유전 적으로 정상을 유지하면 배아줄기세포주가 확립되 었다고 한다. 일단 확립된 세포주는, 또는 이 전이 라도, 일부를 동결 처리해서 다른 실험실에 보낼 수 있고 실험에 이용될 수 있다3).
배아줄기세포주를 만드는 과정에서 세포가 배아 줄기세포의 기본적인 특성을 유지하는지 검사하여 야 하고, 이 과정을 특성화(characterization)라고 한다. 아직은 모두가 인정하는 표준 방법은 없지만 인간 배아줄기세포를 배양하고 있는 실험실에서는 다음의 몇 가지 검사들이 시행된다9). 1) 수개월간 세포를 계대배양하면서 현미경으로 세포 모양을 관 찰하고 세포가 장기간 미분화 상태에서 자가 복제 기능을 유지하고있는 지 확인한다. 2) 미분화 세포 를 확인할 수 있는 표지 인자를 이용한다. Oct-4, SSEA(stage-specific embryonic antigen)-1, 3, 4 와 TRA(tumor rejection antigen)-1-60, 81, al- kaline phsphatase가 대표적인 표지 인자이며, SSEA-1은 쥐 배아줄기세포에는 있지만 인간 배 아줄기세포에는 없고, SSEA-3, 4와 TRA-1-60, 81은 반대이다. Oct-4는 미분화 세포가 만들어내 는 전사 요소(transcription factor)로서, 세포 분화 와 배아 발달의 과정에서 유전자들의 on과 off의 시간 일정(time schedule)을 조절하여 전사 여부를 결정하는 중요한 인자이다. 3) 염색체를 현미경으 로 검사하여 염색체 이상 여부를 검사한다. 하지만 현미경으로 유전자 이상까지 발견하지는 못한다.
4) 세포의 동결, 해동, 대이음 과정을 거친 후에도 계대배양이 되는 지를 검사한다. 5) 인간 배아줄기 세포의 전능을 확인하기 위해 자발적으로 세포들을
분화시켜보거나, 특정 세포로 분화하게끔 세포를 조정해보기도 하고, 기형종(teratoma) 형성을 확인 하기 위해 면역이 억제된 쥐에 세포를 투여하기도 한다. 기형종은 특징적으로 분화가 되거나 부분적 으로 분화된 다양한 세포들의 복합체로서, 배아줄 기세포의 전능을 확인하는 지표가 된다.
인간 배아줄기세포가 치료에 적용되려면 세포를 배양하는 적당한 조건이 절대적이다. 여러 가지 배 양 방법들이 쥐 배아줄기세포에서의 경험에서 응용 되었다. 20여년 전 최초 쥐 배아줄기세포가 면역 수술 방법으로 유도된 이 후, 시토카인(cytokine) mouse embryonic fibroblast(MEF) 영양 세포층 을 이용하여 미분화 상태를 유지하며 배양하였다
10). 이후 leukemia inhibitory factor(LIF)가 발견 되면서 MEF 영양 세포층 없이 LIF만으로 세포 미분화 상태를 유지할 수 있게 되었다11). 1998년 인간 배아줄기세포가 확립되었을 때는 항분열 처리 된 MEF 영양 세포층을 이용하였고, 배양액으로 20% fetal bovine serum(FBS)가 첨가된 Dulbec- co'd modified Eagle's액, 1 mM glutamine, 0.1 mM β-mercaptoethanol, 1% nonessential am- ino acids가 이용되었다5). 하지만 MEF 영양 세포 층을 이용한 배양 방법은 배아줄기세포의 자발적인 분화가 일부에서 관찰되었고 무엇보다도 이종 병원 체 오염의 우려가 있다. 불행하게도 인간 배아줄기 세포는 쥐에서와는 달리 LIF로 미분화 상태를 유 지 할 수가 없다. 그러므로 미분화 상태를 유지하 면서 배양을 할 수 있게 하는 다른 성장 요소를 찾고자 하였는데, 예를 들어 TGFβ1 신호, BMP 신호 방해, 또는 basic fibroblast grow factor (bFGF) 등이 보고되었다12, 13). 하지만 나름의 장단 점으로 인해, 인간에서부터 얻은 재료를 이용한 세 포 배양법의 개발 노력은 계속되고 있다.
이전이라는 과정을 통해 인간 배아줄기세포의 계대배양이 이루어진다. 이전 방법은 collagenase, trypsin, dispase 등의 효소를 이용하는 방법과, micropipette이나 앞을 뾰족하게 만든 Pasteur pi- pette을 이용하여 물리적으로 배아줄기세포만을 골 라서 이전하는 방법이 있다. 물리적인 방법은 배양
과정에서 일부 분화된 세포를 골라낼 수 있어 세 포의 순도를 높일 수 있다는 장점이 있는 반면, 시 간이 많이 소요되고 과정이 힘들어 많은 세포를 동시에 옮길 수 없다14). 반면 효소를 이용한 방법 은 빠르고 간편하지만, 효소에 의해 분리된 세포 덩어리 크기의 변이가 심하고 이미 분화된 세포의 오염도 가능하다. 그러므로 우선 물리적 방법으로 분화된 세포를 골라내고 나서 효소를 이용하는 방 법도 고려될 수 있다15). 하지만 최근에는 효소를 이용한 전이 방법이 염색체 이상의 발현율을 증가 시킨다는 보고가 있어16), 세포 치료에 실제 적용하 기 위해서는 번거롭더라도 물리적 방법이 추천된 다. 계대배양을 하면서 발생하는 염색체 이상의 원 인에 대해서는 연구가 필요하겠지만, 계대배양 과 정에서의 염색체 이상은 세포 치료의 실제 적용에 있어 해결되어야 할 과제이고, 분석되지 않은 유전 자 결함까지 고려한다면 실제 세포 치료까지의 과 정이 쉽지 않다는 것을 실감할 수 있다.
배아줄기세포를 치료 목적에 적당한 세포로 분 화를 유도하게 되는데, 몇 가지 주요한 방법들이 있다. 우선 미분화 상태를 유지하게끔 하는 필수 요소들을 제거하고 자연스럽게 세포들이 덩어리지 게 하면서 배아체(embryoid body)를 만들게 하면, 자궁 내에서와 비슷한 3차원 구조가 만들어지면서 세포끼리의 상호 작용으로 다양한 분화가 시작된 다. 내배엽에서 기원한 세포들이 배아체의 바깥 층 에서 우선 분화되고, 안쪽에서 외배엽 기원의 세포 들이 분화되면서 가운데 빈 공간이 생겨 낭(cyst- ic) 배아체라는 특징적인 모양을 갖추게 된다. 낭 배아체를 부유액(suspension) 배양하면 외배엽과 내배엽 기원 세포 사이로 중배엽 세포들도 분화된 다. 이 후 선택적인 배양액들을 통해 원하는 세포 만을 정제(purification)한다17). 최근 Lee 등18)은 배 아체를 기본으로 쥐 배아줄기세포를 중뇌와 후뇌의 신경세포(neuron)로 분화하는 “5단계” 방법을 개발 하였다. 배아체에 화학적으로 일정한 배양액을 처 리하여 신경세포, 별아교세포(astrocyte), 희소돌기 아교세포 등 신경 관련 세포로의 분화가 가능한 신경 전구체(neural precursor)만을 선택하고 다시
bFGF로 증식 후 분열촉진물질(mitogen)을 제거하 여 최종적으로 신경세포로의 분화를 유도한다. 배 아체를 기본으로 한 분화 방법은 정상 배아 발달 의 과정과 유사한 자연스러운 방법이고 오염의 가 능성은 적으나 얻을 수 있는 세포 수(yield)와 분 화 과정이 효율적이지 못하다는 단점이 있다. 영양 세포층을 이용하여 분화를 유도할 수도 있는데, Kawasaki 등19)이 개발한 같이배양(coculture) 방 법은 쥐 배아줄기세포를 stromal cell-derived inducing activity(SDIA)를 발현하는 PA6 영양 세포층과 함께 배양함으로서 배양 12일째 전체 배 아줄기세포의 약 50%가 Tuj1-양성 신경세포로 분 화되고 신경세포의 30%가 도파민 신경세포로 분 화될 수 있다. 유전자 조작 및 특정 신호 분자를 처리하면 쥐 배아줄기세포의 90%를 중뇌 기원의 도파민 신경세포로 분화시킬 수도 있다20). 역시 쥐 배아줄기세포에서 확립된 방법들을 인간 배아줄기 세포에 응용하게되는데, PA6 영양 세포층과의 같 이배양을 이용한 인간 배아줄기세포의 도파민 신경 세포로의 분화와 더불어, MS5 기질 세포주에 의한 같이배양 방법도 보고되고 있다21). Fig. 3은 저자 들이 쥐배아줄기세포를 PA6 영양 세포층에서 신 경 전구체로 분화시킨 예이다22). 이 외에도 인간 배아줄기세포 배아체에서 선택한 신경 전구체를 bFGF로 증식한 다음 신경구(neurosphere)를 만든 후 정상 체내 신경세포 발생 과정에 정상적으로 포함되는 신호를 주어 다양한 신경세포로 분화하기 도 하고23), 이외 각종 형태발생(morphogenesis) 유도 기질들이 신경세포 분화 유도를 위해 사용된 다24). 가장 이상적인 분화 방법은 정상 배아 발달 의 과정과 가능한 비슷하고, 오염의 가능성이 없으 면서도 분화 방법이 간편하고 쉬우며 많은 수의 원하는 세포로의 분화가 가능하여야 한다. 그러나 아직은 모든 조건을 갖춘 분화 방법은 없다.
배아줄기세포의 유전자 조작은 세포의 효율적인 분화 또는 분화된 세포에서의 특정한 세포의 효율 적인 분리를 위해 시행될 수 있고, 분화 과정의 추 적 및 유전자 기능 평가, 배아줄기세포의 면역성을 변경시킬 목적으로도 연구되고 있다25). 배아줄기세
포에 유전자를 도입시키는 방법으로는 바이러스 감 염, electroporation, 그리고 핵산전달감염(transfec- tion) 방법이 있다26). 하지만 인간 배아줄기세포에
유전자를 도입하기 위해서는 몇 가지 어려움들이 있다. 첫째 단일한 클론(clone)의 세포주를 얻기가 힘들고, 둘째 electroporation 자체가 세포의 생존 Fig. 3. (A) Schematic procedures for in vitro differentiation into neural precusors from mouse embry- onic stem cells on PA6 stromal feeder cell layer. (B) After co-culturing in a embryonic stem cells dif- ferentiation medium I for 3 days, cell bodies were noted (a), at 7 days, embryonic stem cells derived neural rosettes were noted (b), at 1 day after changing into medium II for transplantation, 95% of the embryonic stem cells colonies (c, e) were costained with nestin (d) and NCAM (f) positive cells, at 6 days after medium II, βIII tubulin-positive cells were noted in 30% of embryonic stem cells (g), mean- while, O4 (h) or GFAP-positive cells (i), respectively 5%, and below 1% of embryonic stem cells were detected at 12 days after medium II. Scale bar of a-f, 100 µm. Scale bar of g-i, 50 µm. Cited from Kang et al22). Abbreviation : ES, embryonic stem.
율에 악영향을 미칠 수 있으며, 셋째 리포솜(lipo- some) 매개 핵산전달감염은 효율적이지 못하다26). Lentivirus 매개체 방법은 효율적인 유전자 도입이 가능하고 장기간의 유전자 발현이 가능하며 단백질 을 직접 전달하는 방법도 연구되고 있으나27), 근본 적으로 세포 치료를 위해 이식될 세포에 이러한 조작을 가하는 것이 어떠한 결과를 초래할 지는 연구되어 있지 않다.
3. 성체줄기세포
성체줄기세포는 이미 분화된 조직이나 장기에 존재하면서 자가 복제가 가능하고 세포가 존재하고 있는 특정 조직이나 장기의 주요 세포로 분화가 가능한 세포이다28). 배아줄기세포와는 달리 성체줄 기세포의 기원은 분명하지 않지만, 최근 다양한 조 직에서 성체줄기세포가 발견되고 있다. 1960년대 이미 골수에 조혈모세포(hematopoietic stem cell) 와 더불어 골수 간세포(stromal cell)가 골, 연골, 지방, 섬유 연결 조직을 만드는 세포 군으로 분화 할 수 있음을 알았고, 혈액 조성 줄기세포는 30년 동안 이식을 위해 사용되어 왔다29). 당시 쥐 뇌에 서 신경세포로 분화되며 분열중인 세포가 발견되었 으나, 최근까지도 성체 뇌에서의 신경 줄기세포의 존재는 인정되지 못하였다가 1990년대에 이르러서 야 뇌의 주요한 세 가지 세포 형태인 별아교세포, 희소돌기아교세포, 신경세포를 만들 수 있는 줄기 세포의 존재가 인정되었다30).
성체줄기세포는 다양한 장기와 조직에서 발견되 기는 하지만 매우 적은 수가 존재한다. 그리고 조 직의 손상이나 질병으로 인해 세포가 자극될 때까 지 오랫동안 휴지기를 유지한 채 특정한 장소에 존재한다. 지금까지 알려진 성체줄기세포를 가지고 있는 조직은 뇌, 골수, 말초 혈액, 혈관, 골격 근육, 피부와 간이고, 일반적인 분화 과정은 다음과 같다
28). 1) 조혈모세포는 각종 혈액 세포로 분화하여, 적혈구, B 임파구, T 임파구, 자연세포독성세포 (natural killer cell), 호중성구, 호염기구, 호산성 구, 단핵세포, 탐식세포와 혈소판으로 분화되고, 2) 골수 간세포는 골 세포, 연골 세포, 지방 세포와
건(tendon)에 존재하는 세포와 같은 연결 조직 세 포(connective tissue cell)로 분화된다. 3) 뇌에 존 재하는 신경줄기세포는 신경세포와 별아교세포(as- trocyte), 희소돌기아교세포(oligodendrocyte)로, 4) 소화관의 심부움(deep crypt)에 존재하는 상피 줄 기세포는 흡수 세포(absorbtive cell), 술잔 세포 (goblet cells), Paneth 세포, 장관분비 세포로, 5) 표피(epidermis)의 기저층(basal layer)과 모발낭 (hair follicle)의 기저에 존재하는 피부줄기세포는 각질세포(keratinocyte)로 분화할 수 있고, 피부 표 면으로 이동하여 보호층을 형성한다. 소포(follicu- lar) 줄기세포는 모발낭과 표피로 분화할 수 있다.
성체줄기세포를 확인하고 검사하기 위해 주로 다음의 3가지 방법을 자주 사용한다28). 첫째 생체 조직에 있는 세포에 분자 표식자(molecular mark- er)로 표시하여, 분화된 세포 종류를 확인한다. 둘 째 생체 조직에서 후보가 되는 세포를 제거하고 세포 배양 과정에서 표시를 한 뒤 다시 다른 동물 에 이식하여 세포들이 기원했던 조직의 세포를 만 드는지를 추적한다. 셋째 후보가 되는 세포를 분리 하여 성장 요소를 추가하거나 새로운 유전자를 세 포에 넣어 분화시킨 후 어떠한 종류의 세포로 분 화되는 지를 확인한다. 더불어 줄기세포의 기본적 인 조건으로 하나의 성체줄기세포는 유전적으로 동 일한 세포, 즉 복제 세포주여야 하며, 적어도 세포 가 존재하는 조직의 다양한 세포로 분화할 수 있 는 분화능을 가지고 있어야 한다.
최근에는 성체줄기세포의 세포 기원과 다른 조 직의 세포로의 전분화(transdifferentiation) 능력, 가소성(plasticity)이 보고되고 있다31). 조혈줄기세 포가 신경세포와 별아교세포, 희소돌기아교세포, 골 격 근육 세포, 심장 근육 세포와 간세포로 분화될 수 있고, 골수 간세포는 심장 근육 세포와 골격 근 육 세포로의 전분화가 가능하다32). 뇌 줄기세포는 혈액 세포와 골격 근육 세포로의 전분화가 보고되 었다33). 이러한 성체줄기세포 가소성의 기전이 밝 혀지고 조절될 수 있다면, 보다 다양한 조직으로부 터 손쉽게 줄기세포를 획득하여 세포 치료에 사용 할 수 있을 것이다.
하지만 성체줄기세포를 실제 치료에 이용하기 위해서는 아직도 해결해야 할 많은 과제들이 있다
28). 우선 “얼마나 많은 종류의 성체줄기세포가 조 직에 존재하는가? 체내 성체줄기세포의 기원은 어 디인가? 배아줄기세포의 잔여 세포들인가? 아니면 다른 경로를 통해 발생하였는가? 왜 그리고 어떻 게 성체줄기세포는 주위의 모든 세포들이 분화한 상태에서 미분화 상태를 유지하고 있는가? 성체줄 기세포는 정상적으로도 가소성을 보이는가? 아니 면 연구자들의 실험적인 조절이 필요한가? 가소성 을 설명하는 줄기세포의 증식과 분화를 조절하는 신호는 무엇인가? 성체줄기세포를 조직에 이식하 기 위해 충분한 수를 증식시킬 수 있는가? 특정한 성체줄기세포가 모든 종류의 기관이나 조직의 세포 로 분화가 가능한가? 즉 가소성의 한계는 어디까 지인가? 생체 내에서 성체줄기세포가 손상된 조직 으로 이동하게끔 하는 신호는 무엇인가?” 등이다.
5. 배아줄기세포와 성체줄기세포의 장단점 인간 배아줄기세포와 성체줄기세포는 세포에 근 간한 재생 치료에 사용되기에 앞서 각각의 장단점 을 가지고 있다1). 배아줄기세포는 모든 종류의 세 포가 될 수 있는 전능을 가지고 있는데 비해 성체 줄기세포는 일반적으로 그들이 기원한 조직의 다양 한 세포로만 분화되는 경향이 있고, 전분화능이 보 고되고 있지만 배아줄기세포에 비해 한계가 있다.
배아줄기세포는 세포 치료에 충분할 만큼의 많은 수의 세포가 실험실에서 비교적 쉽게 배양되는데 반해 성체줄기세포는 성체 조직에서 분리할 수 있 는 세포 수가 적고, 증식시키는 방법도 완전히 확 립되어 있지 않다. 성체줄기세포의 가장 큰 장점은
환자의 세포를 배양하여 다시 그 환자에게 줄 수 있고, 배아줄기세포에 비해 윤리적인 문제에서 비 교적 자유롭다. 배아줄기세포는 최근 핵 치환술, 특히 환자 맞춤형 핵 치환술의 개발로 면역 거부 문제가 해결되는 듯하였으나, 연구 자체가 허위로 드러난 바 있다. 특히 배아줄기세포의 전능으로 인 해 성체줄기세포에 비해 기형종이 형성될 우려가 크다. 물론 원하는 세포 전구체로의 부분적인 분화 후 이식하여 기형종의 발생률을 현저히 낮출 수 있지만34), 이 역시 이종 세포 이식에 비해 동종 세 포 이식의 경우 발생률이 증가할 수 있어35), 인간 세포 치료를 위해 인간 배아줄기세포가 이식되어야 함을 고려할 때, 해결되어야 할 과제라고 할 수 있 다(Table 1).
6. 인간 줄기세포의 실제 이용 가능성과 한계 인간 배아줄기세포에 대한 연구는 인간 배아와 태아의 정상적인 발달 과정에서 많은 정보를 얻을 수 있고 또한 줄 수도 있다. 미분화된 줄기세포가 특정 세포로 분화되는 과정의 핵심적인 조절자는 유전자들의 적절한 on, off라고 할 수 있다36). 가장 심각한 의학적 상황인 종양과 선천성 기형 역시 비정상적인 세포 분열과 분화에 기인하므로 유전적 분자생물학적 조절에 대한 이해가 좀더 향상된다면 이러한 질환들이 어떻게 발생하였으며 치료를 위해 어떤 조치들이 필요한 지에 대한 이해를 증진시킨 다37).
새로운 약물은 인간 줄기세포에서 분화된 다양 한 세포를 이용하여 시험 될 수 있다. 약물들 간의 효과를 비교하기 위해서 세포의 조건들이 동일해야 하는데, 분화와 관련된 신호들에 대한 최신 지견은
Table 1. Comparison between Embryonic and Adult Stem Cells
Embryonic stem cells Adult stem cells
Pluripotent
Relatively easy expansion in culture Teratoma formation
Immue rejection Ethical controversy
Multipotent
Difficult to identify, isolate, purify and expansion Less tumorogenic
Possible patient's own cells(marrow, cord blood) Avoidable from ethical problem
이러한 검사를 위해 일관성 있는 동일한 세포로의 분화가 가능하게끔 할 수 있다38, 39).
인간 줄기세포를 사용하기 위해 선행되어야 할 가장 중요한 과제는 역시 세포 치료를 위해 사용 될 수 있는 세포나 조직을 만드는 데 있다. 오늘 날, 장기 또는 조직 이식이 활발하게 이루어지고 있으나 이식 가능한 장기 조직의 공급에는 한계가 있다. 줄기 세포는 바로 특정 세포로 분화하여 Parkinson 병, Alzheimer 병, 척수 손상, 뇌졸중, 화상, 심장 질환, 당뇨, 골관절염, 류마치스 관절염 등의 질환에 세포이식을 하기 위한 세포나 조직의 공급원으로서의 역할을 할 수 있다1).
이렇게 광범위한 난치성 질환에 세포 치료의 희 망을 현실로 이루기 위해서는, 연구자들은 성공적 인 분화와 이식 및 생착에 필요한 특성을 가지도 록 줄기세포를 재연성 있게 만들어 낼 수 있어야 하며, 적어도 다음의 사항들이 충족되어야 한다.
여기에는 1) 세포 치료에 충분한 수와 양의 세포가 만들어질 것, 2) 원하는 세포 형태로 분화할 것, 3) 이식 후 생존할 것, 4) 이식 후 주위 조직과 연결 될 것, 5) 제공자 생존 기간 동안 적절한 기능을 할 것, 6) 어떤 식으로든 제공자에게 해가 되지 않 을 것 등이다. 특히 줄기세포로서의 기본적인 특성 이 훨씬 강한 배아줄기세포의 연구에 있어서는 안 전성과 면역 거부와 관련된 사항들이 효과와 더불 어 세밀히 연구되어져야 한다2).
각 론
줄기세포 이식 또는 내부 줄기세포를 이용한 신
경계 질환의 치료는 미래 의학으로서 많은 관심의 대상이다. 줄기세포 이식 후 임상적으로 호전되는 기전은 손상된 세포가 대체되는 것 뿐만 아니라 다양한 기전에 의할 것으로 예상된다(Table 2)34). 이미 일부 신경계 질환에서는 동물 실험과 함께 임상 시험이 진행되어 희망적인 결과들을 보고하고 있다34). 세포 치료에 가장 근접해있는 신경계 질환 은 성인 신경 퇴행성 질환인 Parkinson 병인데40), 소아에서 발생하는 신경전달물질 질환을 포함한 소 아운동장애의 세포 치료 가능성을 Parkinson 병에 서의 줄기세포 연구 성과를 응용할 수 있을 것이 다. 이 외 난치성 소아신경 질환으로 뇌경색41), 백 질이영양증(leukodystrophy)과 유전성 대사 질환 이 줄기세포 치료의 대상이 될 수 있다42).
1. Parkinson 병
Parkinson 병은 흑색줄무늬(nigrostriatal) 도파 민 신경세포의 퇴행에 의해 발생한다. Parkinson 병 환자에게 인간 태아 중뇌 조직을 이식한 개방 표지(open label) 임상 연구에서 희망적인 결과가
보고되어43, 44), 최근 두 건의 겉보기 수술-조절
(sham operation-controlled) 연구가 시행되었으나 세포 치료에 의한 의미있는 호전을 관찰할 수 없 었고 오히려 일부 환자에서 다른 행동 이상을 보 이기도 하였다45, 46). 하지만 이러한 임상 시험을 통 해 향후 Parkinson 병 세포 치료를 위해 나아갈 방 향을 설정할 수 있는 계기가 되었으며, 임상에 적용 할 이상적인 세포는 적어도 다음의 조건을 충족하 여야 한다34). 첫째 이식할 세포는 도파민을 분비하 고 조절할 수 있어야 하며 분자 생물, 형태 그리고
Table 2. Possible Mechanisms underlying Impromvent after Cell Transplantation. Cited from Lindvall et al.34)
Mechanism Mode of action
Tissue damage
Correction of biochemical deficit Growth factor secretion
Local reinnervation
Reconstruction of neural circuitries
Inflammation-induced stimulation of host plastic response Release of missing transmitter(‘monipump’)
Stimulation of plastic responses and improved survival and func- tion of host neurons
Restoration of synaptic transmitter release
Re-establishment of functional afferent and efferent connections
전기생리적 특성까지 흑색질 신경세포여야 한다.
둘째 Parkinson 병 동물 모델에서 임상적 호전이 검증되어야 한다. 셋째 인간 조가비핵(putamen)에 장기간 생존하기 위해서는 적어도 100,000개 이상 의 도파민 신경세포가 이식되어야 한다. 넷째 이식 된 도파민 신경세포는 주위 줄무늬체와 치밀한 연 결이 이루어져야 하고, 다섯째 신경 회로의 기능이 정상적으로 이루어져야 한다.
현재까지 도파민 신경세포는 쥐 또는 원숭이 배 아줄기세포와 쥐 또는 인간 태아에서 기원한 신경 줄기세포에서 획득되었고, 성체 뇌 또는 다른 조직 에서 분리된 적은 없다34). 최근에는 인간 배아줄기 세포에서도 도파민 신경세포를 유도할 수 있다47). 아직은 모든 조건을 만족하는 세포는 없으나, 쥐 배아줄기세포에서 가장 희망적인 연구 결과가 보고 되고 있고, 실제 임상 적용은 쥐 배아줄기세포에서 의 연구 결과를 응용한 인간 배아줄기세포에 의해 이루어져야 할 것이다48). 배아줄기세포의 특정 신 경 전구체로의 분화는 총론에서 이미 언급한 바 있 다. 분화된 세포는 도파민 신경세포를 포함하여 다 양한 세포가 섞여 있기 때문에, 실제 임상 적용 시 에는 fluorescence-activated cell sorting(FACS) 또는 동종 재조합(homologous recombination)에 의한 트랜스제닉(transgenic) 배아줄기세포를 만들 어 도파민 신경세포만을 선택하여야 한다49). 성체 뇌의 내부 줄기세포로부터 새로운 도파민 신경세포 를 유도하기 위한 연구는 외부 이식에 비해 아직 초보적인 단계이다50).
배아줄기세포를 다루는 기법이 발전함에 따라 많은 수의 도파민 신경세포를 한꺼번에 만들 수
있지만47, 48), 임상 적용을 위해서는 다음 연구들이
진전되어야 한다34). 첫째 Parkinson 병 환자 중에 세포 치료의 적응증이 확립되어야 한다. 도파민 신 경세포의 손실만이 증상의 주요 원인이고 L-dopa 약물에 대한 반응이 좋은 환자가 대상이 될 수 있 을 것이다. 둘째 이식의 위치, 방법, 이식 세포 수 가 일정해야 하고 기존의 태아 조직 이식과 달리 줄기세포를 도파민 신경세포로 분화하는 과정에서 유전자 조작을 통해 이식 후 생존율, 분화, 이동,
기능의 향상이 부작용 없이 가능한 지도 연구되어 야 한다. 셋째 면역 억제가 필요한 지 또는 면역 거부 반응을 극복할 방법이 연구되어야 한다. 체세 포핵 이식에 의한 환자 맞춤형 줄기세포가 대안이 될 수 있겠지만 현재까지 가능하다는 증거는 없다.
넷째 세포 이식에 따른 부작용이 충분히 검증되어 야 한다. 특히 기대가 되는 배아줄기세포에서 분화 된 세포는 실험실에서 일부 분화가 진행된 전구체 이기 때문에 기형종 발생의 우려가 줄었지만 그럼 에도 불구하고 결국 인간에게 인간 배아줄기세포가 이식될 것이므로 동종 이식에 의해 증가하는 기형 종 발생의 위험이 완전히 해결된 것은 아니다.
2. 뇌경색
뇌경색은 Parkinson 병과는 달리 여러 종류의 신경세포와 아교 세포가 함께 손상된다. 다양한 줄 기세포들이 뇌경색 동물 실험에 이용되어 이식된 세포가 확인되었고 행동이 호전됨을 확인하였으며, 인간에서도 기저핵 경색이 발생한 환자들에게 NT- 2 기형암종(teratocarcinoma)에서 유래한 신경세포 를 이식하여 임상적 호전과 함께 기저핵에 대사가 증가하였음을 영상 검사로 확인하였다51). 골수에서 유래한 세포 특히 골수 간세포를 이용한 뇌경색 치료 또는 탯줄 혈액에서 획득한 줄기세포의 이식 은 실제 임상 시험이 이루어지고 있다52). 하지만 이러한 긍정적인 발표에도 불구하고 세포 치료 후 뇌경색이 임상적으로 호전되는 정도는 불완전하며, 해결되어야 할 과제들이 많이 있다.
최근에는 외부 이식에 의한 방법이 아니라 내부 줄기세포의 자가 회복 기능을 유도하는 연구가 대 안으로 떠오르고 있다. 최근 쥐를 이용한 동물 실 험에서 뇌경색으로 인해 뇌실하부에 존재하는 신경 줄기세포 생성이 촉진되고 손상된 줄무늬체로 이동 하여 다양한 신경세포로 분화하는 것이 관찰되었 다. 하지만 대부분의 세포가 수주 내 80% 이상 사 멸하여 결국 0.2% 정도의 세포 대체 효과밖에 없 었다53). 이 후 신경세포 생성을 촉진시키기 위해 fibroblast growth factor-2(FGF-2), epidermal growth factor(EGF), stem cell factor, eryth-
ropoietin, brain-derived neurotrophic factor (BDNF), caspase inhibitors, 항염증 약물이 연구 되고 있다34). 대뇌 피질에서는 새로운 신경세포 형 성에 대한 보고가 거의 없는데, 이는 대뇌 피질에 존재할 가능성이 있는 신경 줄기세포로 부터 신경 세포가 생성되거나 뇌실하부에 존재하는 세포의 이 동을 촉발할 수 있는 신호 부족을 예상해 볼 수 있다54).
이식한 대부분의 세포가 생체 내에서 생존한다 고 하여도 많은 종류의 세포가 한꺼번에 손상되는 뇌경색의 경우 성체 신경 줄기세포 또는 신경세포 전구체로 분화된 배아줄기세포가 이식된 후 다양한 세포로 자연히 분화해줄 것인지, 더불어 주위 신경 조직과 정상적인 회로를 형성하고 간질 발작과 같 은 부작용 없이 정상적인 기능만을 할 것인지는 아직 밝혀지지 않은 세포 발달 기전과 함께 연구 가 진행되어야 할 것이다. 더불어 세포 치료의 효 과를 극대화하기 위해서는 세포 이식과 함께 내부 신경 줄기세포의 신경세포 생성을 촉발하고 더불어 경색 부위에 혈액 공급을 위한 혈관 재생 요소들 을 같이 투여함으로서 신경세포 생성을 증가시킬 수 있다55).
배아줄기세포에서 유래한 신경 전구체와 관련된 연구는 성체 신경 줄기세포에 비해 드문데, 성체 줄기세포에 비해 줄기세포로서의 잠재력이 월등함 에도 불구하고 윤리적 논쟁, 기형종 발생의 한계가 극복되지 못한 측면이 있다35). 하지만 소아에서의 뇌경색은 성인과는 달리 보다 더 광범위한 손상일 경우가 많아 이식을 위한 세포로서 배아줄기세포를 이용한 세포 치료에 비중을 두어야 할 것으로 생 각한다. 최근 저자들은 청소년 주령의 광범위 뇌경 색을 유발한 흰쥐에게 쥐 배아줄기세포에서 분화시 킨 신경 전구체를 직접 주입함으로서 긍정적인 결 과를 얻은 바 있다22).
3. 백질이영양증 및 유전 대사 질환
백질이영양증에서 줄기세포는 손상된 수초(my- elin)를 재생할 수 있는 희소돌기아교세포 또는 신 경집(Schwann) 세포로 분화 되어야 한다. 신경세
포로 분화시키는 것과는 달리 아교세포로의 분화는 초보 단계이다42). 광범위한 수초 재생을 위한 세포 의 기본 조건으로는, 일정 기간 전구체 상태에서 세포 분열이 왕성해야 하고, 손상된 백질 구석구석 으로의 이동성이 뛰어나야 하며 궁극적으로 수초를 충분하게 만들어 내야 한다42). 지금까지 연구 결과, 개와 돼지에서 분리된 아교구(oligosphere)는 만족 스럽지만, 인간 뇌에서 분리된 아교구의 세포 분열 능은 만족스럽지 않다56-58). 태아 뇌에서 분리한 아 교구을 이용한 동물 실험은 긍정적인 결과를 보였 으나59), 인간 태아 뇌의 공급은 한계가 있고 윤리 적인 논쟁을 피할 수 없다. 쥐 배아줄기세포에서 희소돌기아교 세포로의 분화를 유도하여 충분한 수 의 세포를 얻었고 동물 실험에서 기능 호전과 함 께 상당한 수초화를 관찰하였지만60), 인간 배아줄 기세포는 최근까지도 희소돌기세포로의 분화가 매 우 힘들었다. 하지만 2006년 Nistor 등61)이 황색구 (yellow sphere) 과정을 거치면서 신경 전구체의 80% 이상을 희소돌기아교 전구체로 분화 유도하 였다.
줄기세포 치료의 가장 유력한 후보가 될 백질이 영양증은 Krabbe 병, Pelizaeus-Merzbacher 병, 부신백질이영양증과 이염색성 백질이영양증(meta- chromatic leukodystrophy) 등이다42). Krabbe 병 의 경우 globoid 세포라고 알려진 대식세포에 의해 줄기세포가 방해받을 수 있고62), 부신백질이영양증 역시 장쇄 지방산에 의한 염증이 줄기세포에 손상 을 줄 우려가 있다63). Pelizaeus-Merzbacher 병은 염증은 없으나 별아교세포 신경아교증(astrocytic gliosis)이 줄기세포의 이동에 방해가 될 가능성이 있다42). 하지만 이상의 질환은 주로 백질에 병변이 국한되어 있고, 각 질환에 대한 동물 모델이 있다 는 점에서 줄기세포 치료의 우선 대상이 되고 있 다62).
소아에서 발생하는 유전 대사 질환 중 신경계를 침범하는 질환이 많다. 유전자 치료가 관심을 끌었 으나 임상적 성과가 미미하여 세포 치료의 가능성 에 초점이 모아지고 있다64, 65). 하지만 기본적으로 유전 대사 질환은 전신 질환인 만큼 신경계에만
초점을 맞출 수가 없다는 한계가 있다65). 그럼에도 불구하고 Lee 등66)이 보고한 Sanhoff 쥐 모델에서 의 성체 신경 줄기세포와 쥐 배아줄기세포에서 분 화시킨 신경 전구체 이식 실험은 적어도 신경계 병변에 대한 줄기세포치료의 가능성을 보여준 실험 이다. 뇌경색에서도 언급하였듯이 대사 질환에서의 줄기세포의 기능은 단지 손상된 신경세포의 대체뿐 만 아니라, 수초의 형성, 염증 억제, 독성 물질 제 거 및 대사 질환에서 결핍되어있는 효소를 제공하 는 기능을 함께 수행함을 실험적으로 증명하였다 (Fig. 4).
결 론
줄기세포 치료는 특별한 치료 방법이 없는 소아 신경 질환에 여전히 희망적인 분야라고 할 수 있 다. 줄기세포와 관련된 연구 분야는 줄기세포의 획 득에서부터, 배양, 분화, 전 임상 연구과 임상 연구 에 이르기까지 매우 다양하며, 인간에서 실제 임상 적용되기까지 해결되어야 할 많은 과제들이 남아있 다. 이에 동물 실험에서 안전성과 효과가 담보된 임상 시험도 적극적으로 진행되어야겠지만, 분자생 물학, 유전학, 정상 발달과 관련된 기본적인 연구 들이 동물 실험과 함께 내실있게 진행되어야 할 것이다. 최근 보여주기식 임상 연구와 성과 위주의
줄기세포 연구 풍토로 인해 있어서는 안될 논문 조작 사건까지 발생하여 국내 줄기세포 연구가 어 려운 여건에 처해있지만 전화위복의 기회로 삼아 국내 소아신경 학자들의 많은 관심과 참여를 기대 해본다.
References
1) Sell S. Stem cells : What are they? Where do they come from? Why are they here? When do they go wrong? Where are they going?
In : Sell S, editor. Stem cells handbook. 1st ed. Totowa : Humana Press Co, 2004:1-18.
2) Kirschstein R. The stem cell. In : Kirschstein R, editor. Stem cells : scientific progress and future research directions. 1st ed. Maryland : National Institutes of Health, 2001:1-4.
3) Freshney RI. Primary culture. In : Freshney RI, editor. Culture of animal cells : A manual of basic technique. 4th ed. Toronto : Wiley- Liss Co, 2000:149-76.
4) Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse em- bryos. Nature 1981;292:154-6.
5) Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, et al. Embryonic stem cell lines derived from human blastocysts. Science 1998;282:1145-7.
6) Oh SK, Kim HS, Ahn HJ, Seol HW, Kim Fig. 4. Multimodal therapeutic concepts of stem cells in Sand-
hoffs disease. Cited from Lee66).
YY, Park YB, et al. Derivation and charac- terization of new human embryonic stem cell lines : SNUhES1, SNUhES2, and SNUhES3.
Stem Cells 2005;23:211-9.
7) Heins N, Englund MC, Sjoblom C, Dahl U, Tonning A, Bergh C, et al. Derivation, char- acterization, and differentiation of human em- bryonic stem cells. Stem Cells 2004;22:367-76.
8) Kim HS, Oh SK, Park YB, Ahn HJ, Sung KC, Kang MJ, et al. Methods for derivation of human embryonic stem cells. Stem Cells 2005;23:1228-33.
9) Doss MX, Koehler CI, Gissel C, Hescheler J, Sachinidis A. Embryonic stem cells : a prom- ising tool for cell replacement therapy. J Cell Mol Med 2004;8:465-73.
10) Smith AG, Heath JK, Donaldson DD, Wong GG, Moreau J, Stahl M, et al. Inhibition of pluripotential embryonic stem cell differentia- tion by purified polypeptides. Nature 1988;336:
688-90.
11) Williams RL, Hilton DJ, Pease S, Willson TA, Stewart CL, Gearing DP, et al. Myeloid leukaemia inhibitory factor maintains the de- velopmental potential of embryonic stem cells.
Nature 1988;336:684-7.
12) James D, Levine AJ, Besser D, Hemmati-Bri- vanlou A. TGFbeta/activin/nodal signaling is necessary for the maintenance of pluripotency in human embryonic stem cells. Development 2005;132:1273-82.
13) Xu RH, Peck RM, Li DS, Feng X, Ludwig T, Thomson JA. Basic FGF and suppression of BMP signaling sustain undifferentiated prolif- eration of human ES cells. Nat Methods 2005;2:185-90.
14) Rosler ES, Fisk GJ, Ares X, Irving J, Miura T, Rao MS, et al. Long-term culture of hu- man embryonic stem cells in feeder-free con- ditions. Dev Dyn 2004;229:259-74.
15) Oh SK, Kim HS, Park YB, Seol HW, Kim YY, Cho MS, et al. Methods for expansion of human embryonic stem cells. Stem Cells 2005;23:605-9.
16) Brimble SN, Zeng X, Weiler DA, Luo Y, Liu Y, Lyons IG, et al. Karyotypic stability, geno- typing, differentiation, feeder-free maintenance, and gene expression sampling in three human embryonic stem cell lines derived prior to
August 9, 2001. Stem Cells Dev 2004;13:585- 97.
17) Keller GM. In vitro differentiation of embry- onic stem cells. Curr Opin Cell Biol 1995;7:
862-9.
18) Lee SH, Lumelsky N, Studer L, Auerbach JM, McKay RD. Efficient generation of mid- brain and hindbrain neurons from mouse em- bryonic stem cells. Nat Biotechnol 2000;18:
675-9.
19) Kawasaki H, Mizuseki K, Nishikawa S, Ka- neko S, Kuwana Y, Nakanishi S, et al. Induc- tion of midbrain dopaminergic neurons from ES cells by stromal cell-derived inducing ac- tivity. Neuron 2000;28:31-40.
20) Kim DW, Chung S, Hwang M, Ferree A, Tsai HC, Park JJ, et al. Stromal cell-derived inducing activity, Nurr1 and signaling mole- cules synergistically induce dopaminergic neu- rons from mouse embryonic stem cells. Stem Cells. 2006 (In press)
21) Perrier AL, Tabar V, Barberi T, Rubio ME, Bruses J, Topf N, et al. Derivation of mid- brain dopamine neurons from human embry- onic stem cells. Proc Natl Acad Sci USA 2004;101:12543-8.
22) Kang HC. Behavioral improvement after transplantation of neural precursors derived from embryonic stem cells into globally is- chemic brain of adolescent rats(dissertation).
Seoul, Yonsei Med J 2006.
23) Conti L, Pollard SM, Gorba T, Reitano E, Toselli M, Biella G, et al. Niche-independent symmetrical self-renewal of a mammalian tissue stem cell. PLoS Biol 2005;3:e283.
24) Ben-Hur T, Idelson M, Khaner H, Pera M, Reinhartz E, Itzik A, et al. Transplantation of human embryonic stem cell-derived neural progenitors improves behavioral deficit in parkinsonian rats. Stem Cells 2004;22:1246-55.
25) Amit M, Carpenter MK, Inokuma MS, Chiu CP, Harris CP, Waknitz MA, et al. Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative poten- tial for prolonged periods of culture. Dev Biol 2000;227:271-8.
26) Eiges R, Schuldiner M, Drukker M, Yanuka O, Itskovitz-Eldor J, Benvenisty N. Establish- ment of human embryonic stem cell-trans-
fected clones carrying a marker for undif- ferentiated cells. Curr Biol 2001;11:514-8.
27) Ma Y, Ramezani A, Lewis R, Hawley RG, Thomson JA. High-level sustained transgene expression in human embryonic stem cells using lentiviral vectors. Stem Cells 2003;21:
111-7.
28) Kirschstein R. The adult stem cell. In : Kirs- chstein R, editor. Stem cells : scientific prog- ress and future research directions. 1st ed.
Maryland : National Institutes of Health, 2001:
23-42.
29) Becker AJ, McCullough EA, Till JE. Cytol- ogical demonstration of the clonal nature of spleen colonies derived from transplanted mouse marrow cells. Nature 1963;197:452-4.
30) Panicker M, Rao M. Stem cells and neuro- genesis. In : Marshak DR, Gardner DK, Gott- lieb D, editors. Stem cells. 1st ed. New York : Cold Spring Harbor Laboratory Press, 2001:
399-438.
31) Brazelton TR, Rossi FM, Keshet GI, Blau HM. From marrow to brain : expression of neuronal phenotypes in adult mice. Science 2000;290:1775-9.
32) Panicker M, Rao M. Mesenchymal stem cells of human adult bone marrow. In : Marshak DR, Gardner DK, Gottlieb D, editors. Stem cells. 1st ed. New York : Cold Spring Harbor Laboratory Press, 2001:349-74.
33) Anderson DJ, Gage FH, Weissman IL. Can stem cells cross lineage boundaries? Nat Med 2001;7:393-5.
34) Lindvall O, Kokaia Z, Martinez-Serrano A.
Stem cell therapy for human neurodegenera- tive disorders-how to make it work. Nat Med 2004;10:42-50.
35) Erdo F, Buhrle C, Blunk J, Hoehn M, Xia Y, Fleischmann B, et al. Host-dependent tumori- genesis of embryonic stem cell transplantation in experimental stroke. J Cereb Blood Flow Metab 2003;23:780-5.
36) Rodda SJ, Kavanagh SJ, Rathjen J, Rathjen PD. Embryonic stem cell differentiation and the analysis of mammalian development. Int J Dev Biol 2002;46:449-58.
37) Cheng T. Cell cycle inhibitors in normal and tumor stem cells. Oncogene 2004;23:7256-66.
38) Davila JC, Cezar GG, Thiede M, Strom S,
Miki T, Trosko J. Use and application of stem cells in toxicology. Toxicol Sci 2004;79:
214-23.
39) O'Shea KS. Neural differentiation of embry- onic stem cells. In : Zigova T, Sanberg PR, Sanchez-Ramos JR, editors. Neural stem cells : methods and protocols. 1st ed. Totowa : Humana Press Co, 2002:3-14.
40) Moon SY, Park YB, Kim DS, Oh SK, Kim DW. Generation, culture, and differentiation of human embryonic stem cells for therapeutic applications. Mol Ther 2006;13:5-14.
41) Kim HT, Kim IS, Lim SE, Lee IS, Park KI.
Gene and cell replacement via neural stem cells. Yonsei Med J 2004;45:32-40.
42) Duncan ID. Oligodendrocytes and stem cell transplantation: their potential in the treat- ment of leukoencephalopathies. J Inherit Me- tab Dis 2005;28:357-68.
43) Lindvall O, Hagell P. Clinical observations after neural transplantation in Parkinson's disease. Prog Brain Res 2000;127:299-320.
44) Polgar S, Morris ME, Reilly S, Bilney B, Sanberg PR. Reconstructive neurosurgery for Parkinson's disease : a systematic review and preliminary meta-analysis. Brain Res Bull 2003;60:1-24.
45) Freed CR, Greene PE, Breeze RE, Tsai WY, DuMouchel W, Kao R, et al. Transplantation of embryonic dopamine neurons for severe Parkinson's disease. N Engl J Med 2001;344:
710-9.
46) Olanow CW, Goetz CG, Kordower JH, Stoessl AJ, Sossi V, Brin MF, et al. A double-blind controlled trial of bilateral fetal nigral trans- plantation in Parkinson's disease. Ann Neurol 2003;54:403-14.
47) Lacovitti L, Stull ND, Jin H. Differentiation of human dopamine neurons from an embry- onic carcinomal stem cell line. Brain Res 2001;9:99-104.
48) Chung S, Sonntag KC, Andersson T, Bjork- lund LM, Park JJ, Kim DW, et al. Genetic engineering of mouse embryonic stem cells by Nurr 1 enhances differentiation and matu- ration into dopaminergic neurons. Eur J Neu- rosci 2002;16:1829-38.
49) Zwaka TP, Thomson JA. Homologous recom- bination in human embryonic stem cells. Nat
Biotechnol 2003;21:319-21.
50) Zhao M, Momma S, Delfani K, Carlen M, Cassidy RM, Johansson CB, et al. Evidence for neurogenesis in the adult mammalian sub- stantia nigra. Proc Natl Acad Sci USA 2003;
100:7925-30.
51) Kondziolka D, Wechsler L, Goldstein S, Melt- zer C, Thulborn KR, Gebel J, et al. Trans- plantation of cultured human neuronal cells for patients with stroke. Neurology 2000;55:
565-9.
52) Bang OY, Lee JS, Lee PH, Lee G. Autolo- gous mesenchymal stem cell transplantation in stroke patients. Ann Neurol 2005;57:874-82.
53) Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. Neuronal replacement from endog- enous precursors in the adult brain after stroke. Nat Med 2002;8:963-70.
54) Magavi SS, Leavitt BR, Macklis JD. Induc- tion of neurogenesis in the neocortex of adult mice. Nature 2000;405:951-5.
55) Palmer TD, Willhoite AR, Gage FH. Vascular niche for adult hippocampal neurogenesis. J Comp Neurol 2000;425:479-94.
56) Zhang SC, Lipsitz D, Duncan ID. Self-renew- ing canine oligodendroglial progenitor expand- ed as oligospheres. J Neurosci Res 1998;54:
181-90.
57) Zhang SC, Lundberg C, Lipsitz D, O'Connor LT, Duncan ID. Generation of oligodendroglial progenitors from neural stem cells. J Neuro- cytol 1998;27:475-89.
58) Smith PM, Blakemore WF. Porcine neural progenitors require commitment to the oligo- dendrocyte lineage prior to transplantation in order to achieve significant remyelination of demyelinated lesions in the adult CNS. Eur J Neurosci 2000;12:2414-24.
59) Windrem MS, Nunes MC, Rashbaum WK,
Schwartz TH, Goodman RA, McKhann G 2nd, et al. Fetal and adult human oligoden- drocyte progenitor cell isolates myelinate the congenitally dysmyelinated brain. Nat Med 2004;10:93-7.
60) Brustle O, Jones KN, Learish RD, Karram K, Choudhary K, Wiestler OD, et al. Embryonic stem cell-derived glial precursors : a source of myelinating transplants. Science 1999;285:754- 6.
61) Nistor GI, Totoiu MO, Haque N, Carpenter MK, Keirstead HS. Human embryonic stem cells differentiation into oligodendrocytes in high purity and myelinate after spinal cord transplantation. Glia 2005;49:385-96.
62) Suzuki K, Taniike M. Murine model of genet- ic demyelinating disease : the twitcher mouse.
Microsc Res Tech 1995;32:204-14.
63) Powers JM, Liu Y, Moser AB, Moser HW.
The inflammatory myelinopathy of adreno- leukodystrophy : cells, effector molecules, and pathogenic implications. J Neuropathol Exp Neurol 1992;51:630-43.
64) Stein CS, Ghodsi A, Derksen T, Lavidson BL. Systemic and central nervous system correction of lysosomal storage in mucopoly- saccharidosis type VII mice. J Virol 1999;73:
3424-9.
65) Moser HW. Adrenoleukodystrophy : phenotype, genetics, pathogenesis and therapy. Brain 1997;120:1485-508.
66) Lee JP. editor. The multiple actions of neural stem cells may collaborate to improve func- tion & extend life in metabolic neurodegen- erative disorders : evidence from Sanhoff dis- ease mice. Proceedings of the 2005 Seoul Symposium on Stem Cell Research; 2005 Aug 25-26; Seoul. Seoul : Stem Cell Research Cen- ter, 2005.