Kor. J. Appl. Microbiol. Bioeng.
Vol.15 , No.1 , 29-33(1987)
Molecular Cloning of CMCase Gene from an Alkalophilic
Bαcillus Sp. in Escherichia coli
Yu , Ju-Hyun* , In-Soo Kong , Sung-Uk Kim and Jin-Man Kim Deþt. 0/ Food E I/ gi l/ eeri l/ g. YO I1Sι Uniν . 134 SJziη choη -doμg. Serdae /JI oo Jl-J( II.
S‘ι0111 120 , J( orea
Bacillus
속 세균으로부터CMCase
유전자의 대장균에의 형질전환유주현* 공인수, 김성욱,
김진만연세대학교
공과대학식품공학과
A new cellulase gene from Bacillus sp. strain N-4 has been cloned in Escherichia coli with pBR 322. A new plasmid named pYBC 107 was isolated from the E. co /i transformants that produced carboxymethyl cellulase (CMCase). The cloned gene was contained within a 5.5 kb Hind III DNA fragment of the chromosomal DNA part of the plasmid pYBC 107. The CMCase produced ty E. co /i (pYBC 107) located mainly (60%) within the cell wall and showed optimum activity and stability at pH 8.0.
Bacilli elaborate a wide variety of εxtracellular
enzymes , including celluase ( l). The production of these enzyme has attracted much attention because of the interesting question raised as to mechanisms of genetic control and secretion in Gram positive bacte ria. In addition 、 the bioconversion of cellulosic malerial into useful products has becomc a promising fìeld of biotec- nology research during the past serveral years (2 ) . Cellulase genes have been cloned from E5- cherichia adecarboxy!a La (3) , C/ o5lridium Iher- moce//um (4 , 5) , Ce//u!omona5 jìmi (6) , Ther- momono5pora sp. (7) and 8aci//u5 5ubli!i5 (8 ) . Horikoshi et a!. isolated a 8aci/!u5 sp. strain N -4 (A TCC 21833 ) , that produces high levels of an alkaline cellulase complex which consists of at least four isozymes (9 , 10 ) . Two genes for these isoenzymes have been cloned in E. co!i with plasmid vector pBR 322 ( 11) .
In this paper we report the cloning of a new cellulase gene from Bacillu5 sp. strain N- 4 in E.
co /i. To analyze the control mechanisms for thε genes coding for the multicomponεnts of the cel-
ωlase we have attempted to clone.
Materials and κlethods
Bacterial strains and plasmids
8aci//U5 sp. strain N
•4 (ATTCC 21833 ) 、、 as used 1 ‘ or the isolation 01 ‘ lhc cellulase gcne' ‘ E.
co/i MB 1000 ( rι mι , /ac , Irp , pyrF ) (1 2 ) and the plasmid pBR 322 (1 3 ) 、\'cre uscd as lhe cloning host and vector、 respeclively.
Media
E. co/i was grown with shaking at 3 7' C in Luria broth (l Og Difco Bacto lryptone. 5g Ditco yeast extract . 5g NaCl in 11iter ‘ pH 7.2 ) . Anti- biotics used in the selection ofplasmid were added 10 the following fìnal concentralions ( μ g/ m/ ) : ampicillin (Ap) 50 , tetracycline (Tc) 15. The cellulase seleCli\e medium (CSM) conlained 10g Difco Bacto peprone ‘ 19 Bacto yeast eXlraCl. 6g Na 2 HP0 4 , 3g KH 2 P0 4 , O.5g NaCl. 19 NH 4 Cl , 5g CMC per liler of deionized 、\'ater (pH 7. 2).
8aci!!U5 sp. strain N-4 、vas grown in a medium
Key words
‘recombinant DN A. transfor l11 ation. plas l11 id pBR 322 , gene expression. cellulase cO l11 plex
* Corresponding author
29
30 Kor. J. Appl. Microbiol. Bioeng.
conlaining 10g starch , 5g Bacto yeast extrac l, 5g Na acetate. 1 mM Na 2 EDT A , pH 7. 8) (20) and
p o lypeptone、 19 K 2 HP0 4 , O. 2g MgS0 4 • 7 H 2 0 DNA bands were visualized by slaining with eth- and 10g Na 2 C0 3 per liter of deionized water idium bromide and photographcd with a short (pH 10. 0) . Sodium carbonate was sterilized se- - wa、' c length UV transilluminato r. Hind III en-
perately. donuclease clea\'agc of DN A was used as molecu-
DNA isolation
Bacillus sp. strain N - 4 cells wcre harvested at the end of the exponential groι.th phase. Bacterial chromosomal DNA was prepared by the method of Saito and Miura (14) . Plasmid pBR 322 was purified by the method of Tanaka and 씨l eisblum
( 15). The rapid preparation of recombinant plasmids was performed by the miniscreening method (16).
DNA c\ oning
Bacillus sp. DNA (3 μg) and the 、'ector pBR 322 ( 1 μ g) were cleaved with HindIII and then trealed 、、 ith bacterial alkaline phosphatase (1 7) , mixed , and ligated . The ligated mixture was introduced into E. co/i MB1000 (18) and trans- formants were sclccted on LB plate containing 50 μg/ m/ ot ampicillin. The presenc얻 of cloned DN A was verified by transfering colonies onto LB plate containing 15 μ g / m/ of tetracycline . Clones expressing CMCase activity were detected by replica-plating the bacterial colonies onto CSM , as described above. The plates were incubat- ed at 3 7' C for 16 to 24 h. They were then treated for 1 h with chloroform in desiccator jar and air -dried for 30 min prior to covering the surface with 8 m/ of sterile top agar in 10 mM Tris-HCl (pH 8.0) containing 0.5 mg/m/ of lysozyme (Sigma). The plates were incubated at 37
0C for 16-24 h to allow CMCase diffusion. The surfaces of the plates were then flooded with an O. 1%
aqueous solution of congo-red (19) for 30 min Re f1 00ding with 1 M NaCl for 15 min was per- formed to remove unbound dye molecules. The formation of visible halos around the colonies indicates the production of CMCase.
Gel electrophoresis
DNA moecules were seperated on an 0.8%
agarose gel in running buffer (40 mM Tris, 5 mM
lar size reCerence.
Assays of CMCase
For the assay of CMCase , 0.4 m/ of enzyme solution was mixed with 1 m/ of 0.2 M phos- phate buCCer(pH8.0). After 10min oCincuba- tion at 3 7' C , 1 m/ of dinitrosalicylic acid ( 21) solution was added , the solution was well mixed , and the mixture was heated in a boiling water bath for 5 min. The absorbance oC the sample was then measured at 540 nm. One unit of enzyme activity is defined as the amount of enzyme which liberates 0.1 mg reducing sugar , expressed as maltose per min , under the above conditions.
Localization of CMCase in E. coli
Osmotic shock was performed was described by Neu and Heppel (22) . E. co/i cells harbouring pBR322 and pYBC107 werecultured in LB media Cor 24 hrs at 3 7' C. After centriCugation 、
the cells were washed twice in a halfvolume oCthe with 10 mM Tris-HCl buCfer (pH 8.0) and
resuspεndεd in the same volume of 25% sucrose.
The suspεnsionνas shaken for 10 min al room lemperature in the presence oC 1 mM EDT A. The mixture was centrifuged for 10 min al 8, OOOg in
Fig.l. Screening of E. coli Mtll000 contain- ing recombinant CMCase gene by the congo-red staining method.
A: CMCase clone screening after Ap' , Tc
Sselec t \O n.
B: Screening by retransformation of CMCase
hybrid plasmid
Vol.15 , No.l ,
-23.13
- 9.42 - 6.68 - 4.36
31
treated with bactcrial alkaline phosphalasc. The ligalion mixtures \\'crc u 、cd LO lransform E. coli MB 1000: 1. 5X 10 3 Apr lransl ‘ ormanlS pcr μg oftotal DNA were obtained. After replica plating on plalcs containing ampicillin and tClracycline almOSl 31% 01 ‘ thc clonc'、 \\'crc Tcκ indicaling inscrtion ofa foreign DNA fragmen l. Rccombinant clones were lransferred 1O CSM plates in order lo search for CMCase positive IransformanlS. Of 5 , 400 Apr ‘ Tc
Sclones only onc colony 、 ho\\'cd a halo after ly、 is on lhc CSM platc ‘ as dcscri bed i n Materials and Melhods (Fig. l). The hybrid plι-
2.32 Ill id dcsignaled pYBC 107 \\a\ used in rClran 、-
D E
2.03 formation 01 ‘ the original E ωli MB 1000 slrain lO 、 erify lhe prescnce 0 1' CMCase gcnc on lhc pla、Ill id. All 01 ‘ lhe transformant、 이lo\.\'cd a halo
\\'hen these transformants were grown on CSM plates supplemenled with ampicillin
A B C
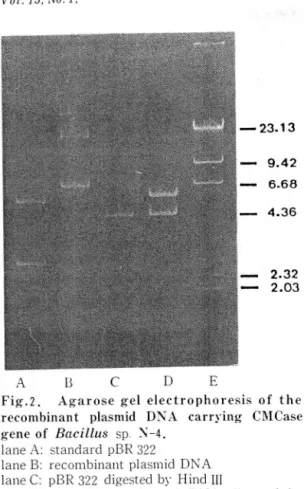
Fig.2. Agarose gel electrophoresis of the recombinant plasmid DNA carrying C l\1 Case gene of Bacillus sp. :'\-4.
lane A: standard pBR 322
lane B: recombinant plasmid DNA lane c: pßR 322 digested by Hind 1II
lane D: recombinant plasmid DNA digested by Hind III
lane E: À DNA digested by Hind l! l as a molec lI
lar \\'eight Ill arker (kb)
a cold room. The 、 upernalanl fluid \\'as rcm O\ cd and lhc \‘ cll drained pellel \\as rapidly mixed \\ith a 、 이 ume ofco ld \\'ater equal lO that ofthe original
、 이 umc 01、 lhe su、 penSIOn ‘ The suspension 、\'as
mi\cd in an icc balh on a termcd the cold 、vater
、、 a 、h ‘ \\'a , remo、 ed. The cells \\ere then suspe nded in lhe samc 、 olumc 01' 10 mM Tris- HCl (pH 7.
5 ) , an aliquot \\ “ sonicalcd at 20 MH z for 5
ml n
‘and intracellular CMCase acti\ity \\'a 、 mea sured. The periplasmic CMCase appeared in the cold 、、 ater 、、 a~、 h
RESULTS
C1 0ning of CMCase gene
The Hind 1Il digested chromosomal DNA of Baci l! u5 sp. slrain N-4 \、 as ligated 、I' ith plasmid pBR322 DNA clea 、 ed \l' ilh Hind 1II and then
Characterization of pYBC 107
Plasmid pYBC 107 which had been iso lated from E. coli cellκ was digested \\'ith Hind lII and analyzed on agarose gel (Fig.2 J. Thc 9.9 kb plasmid pYBC 107 、.\'as composed 01' a 5.5 kb Hind lIl insc rt and 4.4 kb plasmid pBR322.
Localization of Cκ1Case in E. coli (pYBC 107 ) The CMCase produccd by E. co!i(pYBC 107 ) cclls 、vas found a 、 intracellular enzyme ‘ ncarly 60% of the lO lal CMCase aClivily \\'as localizcd in the intraccllular fraClion
,A about 25% 01' the total enzyme rcmain ed in thc peripla 、 m anJ rc、 t 0 1' that (12 ‘’ 0) c
,\crctcd into thc mcdium (Table 1)
Table 1. Distribution of CMCase activity in E.
co /i l\1 B 1000 a containing plasmid DNA.
plasmid CMCase activit y b in following fraction Ext ra cellular Periplasmic Cellular
(% ) (%) (%)
pYBC107 100 (12. 3 ) 210 (25. 3 ) 510 (6 1. 4 )
pBR322 0 0 0
a; E. co !i strains 、.vere aerobically gro\\'n in L -broth for 24 h at 3TC.
b; C l\ lCase activity (at pH 8. 0) v.as expressed as
miliunits per mililiter of the broth
Kor. J. Appl. Microbiol. Bioeng.
100 ‘-
껴‘따 4
1 ‘、 호
r
뇨
→。
。./ ‘
γY
/
•
’ ’
’ ’
’ ’
’
·AU
M
( X )
칼;
;
]얘
@
-넓I잉여
>
E. co /i 32
pH
Fig.4. Effect of pH on the CMCase stability of E. coli MB 1000 containing plasmid DN A pYBC 107 and Bacillus sp. N-4 .
• - . : the intracellular CMCase activity of E coli MB 1000(p YBC 107) .
。 o the extracellular CMCase
Bαα lllls sp. N -4.
10
activitv of 8
6 4
0
DISCUSSION
Alkalophilic Baci//us sp. strain N-4 secrets at least four types of cellulase into the culture medium (1 0). Cellulase is not a single enzyme , but an enzyme complex consisti ng of endo-1 , 4- β
- glucanase , exo - l, 4- ß - gl ucanase and cellobiase We have found CMCase , possessing endo
glucanase activity , present in cell extract of E.
co /i carrying plasmid pYBC 107. This was demon strated by the formation of yellow halos in the congo
•red staining test
On the DNA
levεLlhe CMCase gene ofplasmid P YBC 107 reported here was apparently different from the CMCase genes of plasmid pN K 1 and pNK 2 constructed by Horikoshi el al. (11) which was derived from alkalophilic Bac i/l us sp. strain N - 4 , the same organism we used as donor strain in this study
The distribution of the CMCase in E. coli transformant cells was also different from that of transformants obtained by Horikoshi et al. (11).
Form these we may conclude that we have c1 0ned new gene coding for one of the cellulase complex.
\ \ ]
、
~
‘ τ) a、‘ ~ξî----。、
Properties of Cκ1Case produced by (pYBC 107)
The E. coli (pYBC 107) cells were grown overnight in LB supplemented with ampicillin Cells were concentrated 10 - fold in sonication buffer (1 0 mM Tris-HCl , pH 7. 4 , 5 mM MgCI 2 , 50 μgjml Ribonuclease A) and the cells were ruptured by sonication. The extract was centrifuged at 25 , 000 g in a Hitachi rotor RPR 20 for 30 min. Supernatants were collected and assayed for CMCase activity at various pH ranges from 4 to 10. The maximum activity of the CMCase appeared at pH between 8.0 and 10.0 ( Fig.3 1.
The stability of the enzyme was tested by in- cubating the enzyme in buffer solution having various pH values at 60
0C for 10 min before activity was measured. The enzyme was stable at pH between 7.0 and 8.0(Fig.4 1.
p
~
〉、
| ’ -
':;: 50' r- /
~
/
~ /
걷 /
여 I d /
100
activity of 10 pH
Fig.3. Effect of pH on the CMCase activities of E. coli MB 1000 containing plasmid DNA p YBC 107 and Bacillus sp. N -4 .
• - . : the intracellular CMCase activity of E.
coli MB 1000(p YBC 10 7).
0-- 0 : the extracellular CMCase
Bαα llus sp. N -4 ,
8 6
0 4
Vol.15 , No. 1 ,
요 。 t
--,
적 어 도
4
종류의cellulase
를 생 산하는 것으로 알 려 진alkalophilic
Bacillμssp. strain
N ←4로부터vector plasmid DNA pBR
322을 이 용하여Horiko.
shi
등이cloning
한CMCase
유전자와는 다른 새로 운CMCase
유전자플E. coli
에cloning
하였다.제조된 재조합 plasmid 는
5.5
kb 의 삽입크기를 가 지고 있였으며pYBC
107로 병멍하였다.pYBC 107
를 포함하고 있는E. coli
가 생 산한CMCase
의60%
01 상이 세포내에 존재하였다. 재조합plasmid
에 의하여 생산된 CMCase 는 알칼리 영역에서 높은 활성을 나타내었£며, 대체로 넓은 범위에서50%
이상의 안정성을 보여주었다.