한국어병학회지 제22 권 제3 호(2009) J. Fish Pathol., 22(3)
:
275 ~ 282 (2009)275
Edwardsiella tarda
인위감염에 의한 넙치, Paralichthys olivaceus의 가계별 내성변이
김현철∙손새봄∙정종근∙이정호
�∙최혜승
�∙노재구∙박철지∙민병화∙김종현
��∙김경길
���∙명정인 국립수산과학원 육종연구센터,
�양식환경연구소,
��양식관리과,
���생명공학과
Family variation in survival of olive flounder Paralichthys olivaceus after Edwardsiella tarda infection in challenge tests
Hyun Chul Kim, Sae Bom Sohn, Jong Geun Jeong, Jeong Ho Lee�, Hye Sung Choi�, Jae Koo Noh, Choul-Ji Park, Byung Hwa Min, Jong-Hyun Kim��, Kyung-Kil Kim���and Jeong In Myeong
Genetics and Breeding Research Center, NFRDI, Geoje 656-842, Korea
�Aquaculture Environment Institute, NFRDI, Tongyeong 650-943, Korea
��Biotechnology Research Division, NFRDI, Busan, 619-902, Korea
���Aquaculture Management Division, NFRDI, Busan, 619-902, Korea
In this study, effect of mating group, mating dam and mating sire was analyzed using olive flounder Par- alichthys olivaceus challenged with Edwardsiella tarda. A challenge test was conducted using 1,708 olive flounder fingerlings obtained through 4×4 factorial cross from parents selected based on resistant degree.
Regarding the means of resistant index, survival rate and days of survival by each mating group, the group(RR) produced by mating among resistant parents were 17.98, 13.77%, and 18.36 days, respectively;
the group(SS) produced by mating among susceptible parents were 12.46, 2.71%, and 12.40 days, respec- tively. This study gives an indication that challenge test results may be successfully used as selection criteria for disease resistance to E. tarda in olive flounder, P. olivaceus.
Key words : Olive flounder, Paralichthys olivaceus, Edwardsiella tarda, Selective breeding, Disease Resis- tance, Challenge test
우리나라의 넙치 양식은 1990년대 초반부터 동 해 및 남해안과 제주도 연안에서 발달하기 시작 하였다. 뿐만 아니라 고도의 양식기술이 개발되고 있으며 시설의 규모화가 진전되어 지금까지 주요 양식 산업으로 자리 잡고 있다. 이와 같은 넙치 양식 산업의 급속한 발전으로 양식넙치의 생산량 은 2008년을 기준으로 약 4만 7천 톤으로 전체 어류 양식 생산량의 3.4%에 불과하나, 생산 금액 은 4,083억 원으로 전체 양식 생산 금액의 26.8%
를 차지하게 되었다 (통계청, 「어업생산통계」 ).
그러나 최근에 이르러 잦은 질병의 발생과 이
로 인한 집단 폐사, 치료를 위한 항생제의 과다 한 사용 등은 그 자체 비용 뿐 아니라 궁극적으 로 환경오염과 소비자들의 건강에 위해를 가할 수 있는 요소로 언급되는 등 양식 어류의 불신 으로 이어져 양식 산업 경쟁력 약화의 주요 원 인이 되고 있다. 이에 따라 넙치 양식의 안정적 인 생산을 위한 대안으로 우수한 형질을 지닌 개체를 선발하여 이들의 교배를 거듭하는 전통 적인 선발 육종 방식이 연구되어지고 있다. 이 는 양질의 어미 확보 및 관리와 더불어 우수한 품종을 개발하여 장기적이고 체계적인 육종 연
�Corresponding Author : Jeong Ho Lee, Tel : 055-633-1272 Fax : 055-633-0891, E-mail : [email protected]
9...
구로써 오랫동안 이에 대한 연구가 이루어지고 있으며, 이를 활용하여 질병에 대한 저항성이 향상된 개체의 생산 가능성을 제시하였다 (Cipriano and Heartwell, 1986; Ehlinger 1964, 1977; Embody and Hayford, 1925; Gjedrem and Aulstad, 1974; Gjedrem, 1983; Standal and Gjerde, 1987). 최근 대서양 연어 (Gjedrem and Gj∅en, 1995;
∅deg rd et al., 2006), 무지개 송어 (Hen- ryon et al., 2002, 2005), 잉어 (Das Mahapatra et
al., 2008),대서양 대구 (Kettunen and Fjalestad, 2006)에 대한 연구는 질병 내성의 향상에 대한 실현 가능성을 한층 더 높여주었다.
Edwardsiella tarda
는 많은 담수어나 해산어에 질병을 일으키는 에드워드증 (edwardsiellosis)의 원인균으로 장내 세균과에 속하는 그람 음성 세 균이다 (Thune et al., 1993). 우리나라의 주요 양 식 산업 대상 어종인 넙치에서 보고된 바 있으 며 (Nakatsugawa, 1983), 주요 증상으로 복부팽 만과 탈장, 간과 신장의 농양 형성 등 전신적 증 상을 나타내며 많은 폐사와 함께 양식 산업에 큰 경제적 손실을 발생시키고 있다. Ewing et al.
(1965)에 의해 E. tarda가 어류 병원체로 보고된 이후 병원체의 감염 기작, siderophores, cell adhe- sion factors, cell invasion activity, two types of haemolysin와 같은 잠재적 병원성 인자, 항생제, 백신에 대한 연구가 활발히 진행되고 있다 (Mo- hanty and Sahoo, 2007). 그러나 명확한 메카니즘 이나 치료 대책이 마련되고 있지 않아 이에 대 한 다른 방향으로의 연구가 시급한 실정이다.
본 연구는 넙치 양식에 있어 질병에 내성을
지니는 우수한 품종을 개발하기 위한 품종 개량 의 기초 연구로 넙치 질병 내성 검증 가계를 대 상으로 E. tarda를 인위 감염시켜 내성지수, 생존 율 및 생존일수를 조사함으로써 넙치의 질병내 성에 대한 가계별 내성변이를 검증하여 양식되 고 있는 넙치의 유전적 개량을 위한 기초 자료 를 제공하기 위해 실시하였다.
재료 및 방법
시험 균주
시험 균주는 경남지역의 양식생물질병역학조 사에서 간충혈, 신장비대, 복수 증상을 나타내는 넙치에서 분리된 E. tarda를 사용하였다. 배양은 1% mannitol 이 첨가된 Salmonella-Shigella (SS, Difco) 배지에서 25℃, 48시간 배양후 흑색 colony를 1.5% NaCl 첨가한 Tryptic soy agar (TSA, Difco) 에 도말하여 27℃, 24시간 배양 후 인위 감염 실험에 사용하였다.
시험어
시험어로 사용한 넙치는 국립수산과학원 육종 연구센터에서 2008년 부화시켜 생산한 넙치 종 묘를 이용하였다. 에드워드 질병 내성 검증을 위하여 2007년에 실시한 육종넙치 핵집단 2세 대의 인위 감염 결과와 친자 확인 결과를 기초 로 가계별 내성지수를 계산하여 이를 근거로 내 성 가계와 민감 가계를 선정하였다(Table 1). 질 병 내성 가계와 민감 가계 중 성숙 정도와 인공
Table 1. Edwardsiellosis resistivity of parents used in family production
Dam Sire Number of survival Number of total Resistant index Survival rate(%) Days of survival
D1 S1 3 22 32.82 13.64 32.8
D2 S2 3 50 21.00 6.00 26.0
D3 S3 0 36 8.72 0.00 10.2
D4 S4 0 19 7.74 0.00 7.7
(D1: Resistant dam 1, D2: Resistant dam 2, D3: Susceptible dam 1, D4: Susceptible dam 2, S1: Resistant sire 1, S2: Resistant sire 2, S3: Susceptible sire 1, S4: Susceptible sire 2)
수정 가능 정도에 따라 각각 2가계를 최종 선정 하여 4 × 4 팩토리얼 디자인에 의해 16가계를 생산하였다(Table 2).
자치어는 동일한 환경에서 사육하기 위해 각 가계당 3 ㎖ 수정란을 FRP 5톤 사육수조에 혼합 하여 사육하였다. 먹이로는 부화 직후 소량의 클 로렐라를 사육수에 혼합하였고, 부화 후 3일령부 터는 로티퍼를 공급하였으며, 성장함에 따라 알 테미아와 배합사료를 공급하였다. 사육수의 관 리는 배합사료 공급 이전에는 지수식으로, 그 이 후부터는 일부 환수와 유수식으로 관리하였다.
사육기간 동안의 수온은 17.6~20.4℃, 염분은 30.7~34.8 psu, DO는 6.0~7.9 ㎎/ℓ 범위였다.
인위 감염 시험
시험 균주에 대한 넙치의 폐사율을 조사하기 위하여 부화 후 118일령까지 사육한 넙치 종묘
를 이용하였으며, 인위 감염시 체장은 10.5~15.2
㎝ 범위였다. 인위감염 시험은 총 1,708마리의 넙치 종묘를 동일 수조에서 4×10
6cfu/㎖ 농도 의 시험균주로 2.5시간동안 침지하여 실시하였 다. 수온은 24 ± 0.5℃로 유지하였으며, 폐사율 은 1일에 4회씩 34일간 기록하였다. 폐사어는 폐사 원인을 확인하기 위해 폐사 즉시 해부하고 간, 신장과 비장 조직을 무균적으로 절취하여 Salmonella-Shigella (SS, Difco) 배지에 도말한 후 E. tarda를 재분리하였다. 인위 감염 후 친자 확인된 개체수는 1,705마리였으며, 가계별 개체 수는 Table 3과 같다.
친자 확인
친자확인의 시료는 계측형질 측정시 각 개체 의 넙치 가슴지느러미 조직을 절취한 다음 phe- nol extraction 방법으로 DNA를 추출하여 multi-
Table 2. Family production for Edwardsiellosis resistivity verification♀ Resistant dam Susceptible dam
♂ D1 D2 D3 D4
Resistant sire S1 E11 E21 E31 E41
S2 E12 E22 E32 E42
Susceptible sire S3 E13 E23 E33 E43
S4 E14 E24 E34 E44
: RR : SR : RS : SS
Table 3. Number of offspring identified for each of the families generated in the full factorial cross Dam
D1 D2 D3 D4 Observed total
per sire
Sire S1 100 95 75 129 399
S2 40 107 222 138 507
S3 102 54 79 55 290
S4 117 59 242 91 509
Observed total
per dam 359 315 618 413 1,705
plex PCR 반응에 사용하였다. 유전자서열분석기 (ABI 3100)를 이용하여 8개 유전좌위에 대한 각 개체의 유전자형을 결정하였으며, 친자확인은 멘델의 유전법칙에 위배되는 친어를 배제시키 는 방법(Exclusion method)으로 친어 및 자손의 유전자형을 분석하여 추정된 부모 조합을 교배 지침과 비교하는 방법으로 실시하였다.
통계학적 분석
교배 그룹별, 교배 암컷별, 교배 수컷별 내성 지수(Resistant index), 생존율(Survival rate), 생존 일수(Days of survival)를 조사하였으며, 내성지 수, 생존율 및 생존일수는 아래의 공식을 이용
하였다. 조사 형질을 대상으로 교배그룹, 교배암 컷, 교배수컷의 효과를 분석하기 위해 SAS Package (Ver. 8.2)를 이용하여 ANOVA-test를 실 시하였으며, 평균치간의 유의성 검정은 Duncan's multiple range test를 실시하여 귀무가설을 유의 수준 5%로 각각 검정하였다.
결 과
교배 그룹의 효과
교배에 의해 생산된 그룹에 대한 내성지수를 조사한 결과(Table 4, Fig. 1), 내성 친어간의 교배 (RR), 내성 암컷과 민감 수컷간의 교배(RS), 민 감 암컷과 내성 암컷간의 교배(SR) 및 민감 친 어간의 교배(SS)의 내성지수가 각각 17.98±
1.62, 15.36±0.90, 14.81±0.69, 12.46±0.41로 나
Fig. 1. Survival rates of olive flounder challenged withE. tarda by mating group
(RR: Resistant dam × resistant sire, RS: Resistant dam × susceptible sire, SR: Susceptible dam × resistant sire, SS:
Susceptible dam × susceptible sire)
Table 4. Resistant index, survival rates, days of survival of olive flounder challenged with E. tarda by mating group
Mating group Resistant index Survival rate(%) Days of survival
RR 17.98±1.62a 13.77±5.85a 18.36±0.51a
RS 15.36±0.90b 3.66±1.40b 15.33±0.42b
SR 14.81±0.69bc 7.32±3.46ab 14.56±0.35b
SS 12.46±0.41c 2.71±0.74b 12.40±0.35c
a, b, c,: different letters in the same column indicate significant differences(α=0.05, a>b>c)
(RR: Resistant dam × resistant sire, RS: Resistant dam × susceptible sire, SR: Susceptible dam × resistant sire, SS: Sus- ceptible dam × susceptible sire)
타나 내성 친어간의 교배에 의해 생산된 그룹이 다른 그룹에 비해 유의적으로 내성지수가 높은 것으로 나타났으며, 민감 친어간의 교배에 의해 생산된 그룹이 유의적으로 내성지수가 낮은 것 으로 나타났다(p<0.05).
생존율의 경우, RR, RS, SR, SS가 각각 13.77
±5.85%, 3.66±1.40%, 7.32±3.46%, 2.71±
0.74%로 나타나 RR 그룹이 RS, SR, SS보다 각 각 10.1%, 6.5%, 11.1% 생존율이 높은 것으로 나 타났다(p<0.05).
생존일수의 경우, RR, RS, SR, SS가 각각 18.36
±0.51일, 15.33±0.42 일, 14.56±0.35일, 12.40
±0.35일로 나타나 동일 인위 감염 조건하에서 RR 그룹이 RS, SR, SS보다 각각 3.0 일, 3.8 일, 6.0 일을 더 생존한 것으로 나타났다(p<0.05).
교배 암컷의 효과
교배에 사용된 암컷에 대한 내성지수를 조사 한 결과(Table 5, Fig. 2), 내성 암컷 D1, D2가 각 각 15.66±0.66, 17.67±1.86으로 나타났으며, 민 감 암컷 D3, D4는 각각 14.13±0.87, 13.15±
0.81 로 나타나 내성 암컷 D2가 민감 수컷 D3, D4보다 유의적으로 내성지수가 높은 것으로 나 타났다(p<0.05).
생존율의 경우, D1, D2, D3, D4가 각각 6.43±
2.52%, 11.00±6.58%, 7.78±3.22%, 2.25±0.81%
로 나타나 내성 암컷 D2의 생존율이 가장 높은 것으로 나타났으나 유의적인 차이는 보이지 않 았다.
생존일수의 경우, D1, D2, D3, D4가 각각 15.63±0.44일, 18.28±0.50일, 13.71±0.50일, 13.38±0.35일로 나타나 동일 인위 감염 조건하 에서 내성 암컷 D2이 D3, D4보다 각각 4.6일, 4.9일을 더 생존한 것으로 나타났다(p<0.05).
교배 수컷의 효과
교배에 사용된 수컷에 대한 내성지수를 조사 한 결과(Table 6, Fig. 3), 내성 수컷 S1, S2가 각 각 17.58±1.68, 15.21±0.99로 나타났으며, 민감 수컷 D3, D4는 각각 14.97±1.12, 12.85±0.60으 로 나타나 내성 수컷 S1이 민감 수컷 S3, S4보 다 유의적으로 내성지수가 높은 것으로 나타났
Table 5. Resistant index, survival rates, days of survival of olive flounder challenged with E. tarda by mating dam
Mating dam Resistant index Survival rate(%) Days of survival
D1 15.66±0.66ab 6.43±2.52 15.63±0.44b
D2 17.67±1.86a 11.00±6.58 18.28±0.50a
D3 14.13±0.87b 7.78±3.22 13.71±0.50c
D4 13.15±0.81b 2.25±0.81 13.38±0.35c
a, b, c,: different letters in the same column indicate significant differences(α=0.05, a>b>c) (D1: Resistant dam 1, D2: Resistant dam 2, D3: Susceptible dam 1, D4: Susceptible dam 2)
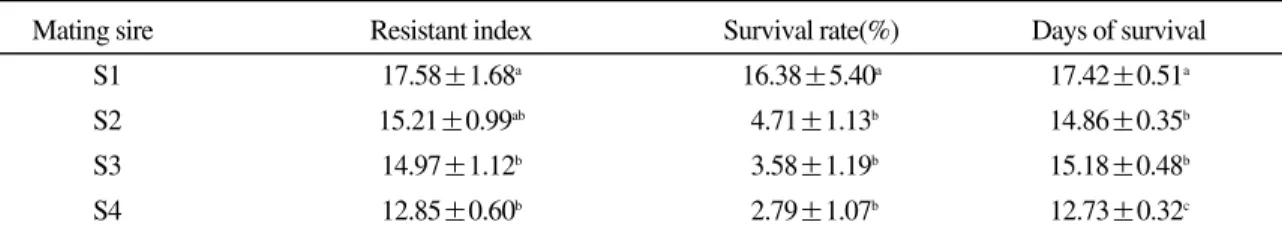
Table 6. Resistant index, survival rates, days of survival of olive flounder challenged with E. tarda by mating sire
Mating sire Resistant index Survival rate(%) Days of survival
S1 17.58±1.68a 16.38±5.40a 17.42±0.51a
S2 15.21±0.99ab 4.71±1.13b 14.86±0.35b
S3 14.97±1.12b 3.58±1.19b 15.18±0.48b
S4 12.85±0.60b 2.79±1.07b 12.73±0.32c
a, b, c,: different letters in the same column indicate significant differences(α= 0.05, a>b>c) (S1: Resistant sire 1, S2: Resistant sire 2, S3: Susceptible sire 1, S4: Susceptible sire 2)
다(p<0.05).
생존율의 경우, S1, S2, S3, S4가 각각 16.38±
5.40%, 4.71 ±1.13%, 3.58±1.19%, 2.79±1.07%
로 나타나 내성 수컷 S1의 생존율이 민감 수컷 S3, S4 보다 각각 12.8%, 13.6% 높은 것으로 나 타났다(p<0.05).
생존일수의 경우, S1, S2, S3, S4가 각각 17.42
±0.51일, 14.86±0.35일, 15.18±0.48일, 12.73±
0.32일로 나타나 동일 인위 감염 조건하에서 내 성 수컷 S1이 S3, S4보다 각각 2.2 일, 4.7일을 더 생존한 것으로 나타났다(p<0.05).
고 찰
선발 육종은 수산생물의 유전적 개량을 위한 가장 일반적이고 효과적인 방법으로 이용되고 있으며, 특히 어류 및 패류에서 선발 육종 및 육 종효율 예측을 목적으로 중요 경제형질들에 대 한 양적 유전학적 평가가 적용되어지고 있다(Ar- gue et al., 2002; Gjerde et al., 2004; Zheng et al., 2006).
본 연구는 넙치 양식에 있어 질병에 내성을 지니는 우수한 품종을 개발하기 위한 품종 개량 의 기초 연구로 실시되었다. 에드워드 질병 내
성 검증을 위하여 2007년에 실시한 육종넙치 핵 집단 2세대의 인위 감염 결과와 친자 확인 결과 를 기초로 가계별 내성지수를 계산하였으며, 이 를 근거로 내성 가계와 민감 가계를 선정하였 다. 질병 내성 가계와 민감 가계 각각 2가계, 암∙수 각각 4마리를 선정하여 4 × 4 팩토리얼 디자인에 의해 16가계를 생산하였으며, 동일 수 조에서 부화후 118일까지 사육한 후 E. tarda에 인위적으로 감염시켜 매일 발생하는 폐사 개체 에 대하여 친자확인을 실시하였다. 친자확인 결 과를 기초로 내성지수, 생존율, 생존일수에 영향 을 미치는 교배 그룹, 교배 암컷, 교배 수컷의 효과를 분석하였다.
내성 암컷 2마리와 내성 수컷 2마리간의 교배 에 의해 생산된 교배그룹이 내성지수, 생존율 및 생존일수 모든 조사형질에서 다른 교배그룹 보다 유의적으로 내성이 높은 것으로 나타났으 며, 민감 친어간의 교배를 통해 생산된 교배그 룹은 유의적으로 질병 내성이 낮은 것으로 나타 났다. 이와 같은 결과는 2007년에 실시한 핵집 단의 인위 감염 실험 결과를 재현하는 것으로 에드워드 내성관련 형질도 일반적인 성장 형질 과 마찬가지로 선발 대상형질로 이용이 가능할 것으로 판단된다.
Fig. 3. Survival rates of olive flounder challenged with E. tarda by mating sire
(S1: Resistant sire 1, S2: Resistant sire 2, S3: Susceptible sire 1, S4: Susceptible sire 2)
Fig. 2. Survival rates of olive flounder challenged with E. tarda by mating dam
(D1: Resistant dam 1, D2: Resistant dam 2, D3: Susceptible dam 1, D4: Susceptible dam 2)
교배에 사용된 친어 개체중 내성 암컷 D2를 이용하여 생산된 후대의 모든 에드워드 내성관 련 형질에서 다른 교배 암컷에 비해 내성이 높 은 것으로 나타났으며, 반대로 교배 암컷 D4를 이용하여 생산된 후대의 에드워드 내성은 낮은 것으로 나타났다. 내성 수컷인 S1을 이용하여 생산된 후대의 모든 에드워드 내성관련 형질에 서 다른 교배 수컷에 비해 내성이 높은 것으로 나타났으며, 반대로 교배 수컷 S4를 이용하여 생산된 후대의 에드워드 내성은 낮은 것으로 나 타났다. 따라서 2007년도 핵집단의 인위감염 실 험을 통해 선발된 D1, D2, S1, S2 친어 가운데서 도 각 친어의 개체별 내성 정도에서 차이가 나 타나는 결과는 가계별 질병 내성 정도를 분석하 여 가계 선발하는 것도 중요하지만, 팩토리얼 교배로 후대 생산을 통한 개체 선발도 병행하는 것이 바람직하다는 것을 나타낸다.
이상의 결과에서 동일한 환경 조건에서 에드 워드 질병 감염 실험에 의한 질병 내성 정도를 확인한 결과, 가계간, 개체간 내성 정도의 차이 가 존재하는 것을 확인하였다. 이는 질병 내성 관련 형질도 유전적인 변이가 존재하여 선발 대 상 형질로 적합하다고 판단되며, 보다 정확한 선발을 위해 각 형질별로 유전력을 기초로한 육 종가를 추정하여 이를 기준으로 선발하는 것이 바람직할 것으로 판단된다.
요 약
본 연구는 에드워드 내성 정도를 기준으로 선 발된 넙치 친어를 이용하여 4×4 펙토리얼 교 배를 통해 얻어진 16가계 넙치의 에드워드 인위 감염 결과를 이용하여 교배 그룹, 교배 암컷, 교 배 수컷의 효과를 분석하였다. 내성 친어간의 교배에 의해 생산된 교배구(RR)의 내성 지수, 생존율, 생존 일수는 각각 17.98, 13.77%, 18.36 일로 나타났으나, 민감 친어간의 교배에 의해 생산된 교배구(SS)는 각각 12.46, 2.71%, 12.40일 로 나타났다. 이상의 결과에서, 에드워드 질병
내성 형질에 대한 가계간, 개체간 내성 정도의 차이가 존재하는 것을 확인하였고, 인위 감염 결과를 이용하여 질병 저항성에 대한 선발이 가 능할 것으로 판단된다.
감사의 글
이 연구는 국립수산과학원(육종기술개발, RP- 2009-AQ-091)의 지원에 의하여 연구 되었습니다.
참 고 문 헌
Argue, B.J., Arce, S.M., Lotz, J.M. and Moss, S.M.:
Selective breeding of Pacific white shrimp (Litopenaeus vannamei) for growth and re- sistance to Taura Syndrome Virus. Aquacul- ture, 204:447-460, 2002.
Cipriano, R.C. and Heartwell, C.M.: Susceptibility of salmonids to furunculosis: differences be- tween serum and mucus responses against
Aeromonas salmonicida. Trans. Am. Fish.Soc., 115:83-88, 1986.
Das Mahapatra, K., Gjerde, B., Sahoo, P.K., Saha, J.N., Barat, A., Sahoo, M., Mohanty, B.R.,
∅
deg rd, J., Rye, M. and Salte, R.: Genetic variations in survival of rohu carp (Labeo
rohita, Hamilton) after Aeromonas hy- drophila infection in challenge tests., Aqua-culture, 279:29-34, 2008.
Ehlinger, N.F.: Selective breeding of trout for resis- tant to furunculosis., N. Y. Fish Game J., 11:78-90, 1964.
Ehlinger, N.F.: Selective breeding of trout for resis- tant to furunculosis., N. Y. Fish Game J., 24:25-36, 1977.
Embody, G.C. and Hayford, C.O.: The advantage
of rearing brook trout fingerlings from se-
lected breeders., Trans. Am. Fish. Soc.,
55:135-142, 1925.
Ewing, W.H., McWhorter, A.C., Escobaar, M.R.
and Lubin, A.H.: Edwardsiella, a new genus of Enterobacteriaceae based on a new species., Int. Bull. Bacteriol. Nomencl. Tax- on., 15:33-38, 1965.
Gjedrem, T. and Aulstad, D.: Selection experiment with salmon. I. Differences in resistance to vibrio disease of salmon parr (Salmo salar)., Aquaculture, 3:51-59, 1974.
Gjedrem, T.: Genetic variation in quantitative traits and selective breeding in fish and shellfish., Aquaculture, 33: 51-72, 1983.
Gjedrem, T. and Gj ∅en, H.M.: Genetic variation in susceptibility of Atlantic salmon, Salmo
salar L. to furunculosis., Aquaculture, 97:1-6, 1995.
Gjerde, B., Terjesen, B.F., Barr, Y., Lein, I. and Thorland, I.: Genetic variation for juvenile growth and survival in Atlantic cod (Gadus
morhua). Aquaculture, 236:167-177, 2004.Henryon, M., Berg, P., Olesen, N.J., Kj ær, T.E., Slierendrecht, W.J., Jokumsen, A. and Lund, I.: Selective breeding provides and approach to increase resistance of rainbow trout (On-
chorhynchus mykiss) to the disease, entericredmouth disease, rainbow trou fry syn- drome, and viral haemorrhagic septicaemia., Aquaculture, 250:621-636, 2005.
Henryon, M., Jokumsen, A., Berg, P., Lund, I., Ped- ersen, P.B., Olensen, N.J. and Slierendrecht, W.J.: Genetic variation for growth rate, feed conversion efficiency, and disease resistance exists within a farmed population of rainbow
trout., Aquaculture, 209:59-76, 2002.
Kettunen, A. and Fjalestad, K.: Resistance to vib- riosis in Atlantic cod (Gadus morhua L.) : First challenge test results. Aquaculture, 258:263-269, 2006.
Mohanty, B.R. and Sahoo, P.K: Edwardsiellosis in fish: a brief review., J. Biosci., 32:1331- 1344, 2007.
Nakatsugawa, T.: Edwardsiella tarda isolated from cultured young flounders., Fish Pathol., 18:99-101, 1983.
∅
deg rd, J., Olesem, I., Gjerde, B. and Klemets- dal, G.: Evaluation of statistical models for genetic analysis of challenge test data on fu- runculosis resistance in Atlantic salmon (Salmo salar): prediction of field survival., Aquaculture, 259:116-123, 2006.
Standal, M. and Gjerde, B.: Genetic variation in survival of Atlantic salmon during the sea rearing period., Aquaculture, 66:197-207, 1987.
Thune, R.L., Stanley, L.A. and Cooper, R.K.:
Pathogenesis of Gram-negative bacterial in- fections in warm water fish., Annu. Rev.
Fish Dis., 3:37-68, 1993.
Zheng, H., Zhang, G., Liu, X., and Guo, X.; Sus- tained response to selection in an introduced population of the hermaphroditic bay scallop
Argopecten irradians irradians Lamarck(1819). Aquaculture, 255:579-585, 2006.
Manuscript Received : November 19, 2009 Revised : December 15, 2009 Accepted : December 21, 2009