Copyright ⓒ 2010, The Microbiological Society of Korea
남태평양에 서식하는 두 종의 해면 Hyrtios sp.와 Callyspongia sp.의 공생세균 군집의 다양성
박진숙
*
한남대학교 생명공학과Bacterial Community Diversity Associated with
Two Marine Sponges from the South Pacific Ocean based on 16S rDNA-DGGE analysis
Jin-Sook Park
Department of Biotechnology, Hannam University, Deajeon 305-811, Republic of Korea (Received September 10, 2010/Accepted September 28, 2010)
The bacterial community structure associated with two marine sponges, Hyrtios sp. 604 and Callyspongia sp. 612 collected from the South Pacific Ocean were analyzed by 16S rDNA-denaturing gradient gel electrophoresis (DGGE). The phylogenetic analysis showed that the bacterial community associated with Hyrtios sp. 604 contained diverse bacterial groups such as Chloroflexi, Firmicutes, Cyanobacteria, Alphaproteobacteria, Gammaproteobacteria, Actinobacteria, and Acidobacteria. Callyspongia sp. 612 harbored Chloroflexi, Cyanobacteria, Alphaproteobacteria, and Gammaproteobacteria. Hyrtios sp.
604 belonging to genus Hyrtios known to produce natural products showed greater bacterial diversity than Callyspongia sp. 612. Phylum Actinobacteria was shown to be one of dominant bacterial groups in Hyrtios sp. 604. Although the same phyla of bacteria were found in both sponge species, the sponge- associated predominant bacterial groups differed between the two sponges with different chemical characteristics from the same geographical location. Uncultured bacteria represented over 90% of the bacteria diversity present in all bacterial communities of the sponges.
Keywords: 16S rDNA, bacterial community, DGGE, marine sponge, sponge-associated bacteria
해면은 오랜 진화적 역사를 갖는 다세포 여과섭식 무척추동 물로써 지금까지 약 8,000여 종이 기재되어 있으며 해양 및 담 수에 서식하는 것으로 알려져 있다. 이 중 열대지역의 해양에 가장 많은 종이 분포하며, 반면 담수에는 1% 이하의 해면 만 이 서식하는 것으로 알려져 있다(19). 해면은 Calcarea, Demo- spongiae, Hexactinellida, 3개의 주요 강으로 나뉘며 가장 많 은 해면 종들이 Demospongiae 강에 속하는 것으로 보고되어 있다(17). 해면은 여과섭식 동물로써 주변의 해수 혹은 담수의 여과를 통하여 먹이원을 공급받으며, 이때 미생물과의 공생을 통하여 영양원을 얻거나 골편의 안정화, 노폐물의 가공을 수행 하고 혹은 미생물 감염에 저항하기 위하여 많은 종류의 이차 대사산물을 생산하는 것으로 알려져 있다. 해양생태계에서 해 면의 서식에 중요한 역할을 하는 이러한 이차 대사산물에는 항 바이러스, 항세균, 항암물질, 세포독성, 항염증성 물질 등 의학
* For correspondence. E-mail: [email protected]; Tel: +82-42-629-8771;
Fax: +82-42-629-8769
적으로 중요한 물질들이 포함된다. 실제 해양으로부터 유래하 는 생물의약품 중 대부분이 무척추동물로부터 유래하며 그 중 해면으로부터 가장 많은 종류의 생리활성물질이 생산되는 것 으로 알려져 있다(19). 이에 따라 새로운 생리활성물질을 해면 으로부터 추출하기 위하여 많은 연구들이 지속되어오고 있으 며 많은 종류의 생리활성물질들이 보고되고 있다(12, 19). 또한 이러한 생리활성물질들이 공생하는 미생물에 의해 생산된다는 보고도 증가하고 있다(11). 해면 공생미생물에 의해 생리활성 물질의 생산이 가능하다면 해면 종 자원의 고갈을 방지할 수 있으며 보다 안정적으로 생리 활성 물질을 생산할 수 있을 것 이다. 공생 미생물의 산업적 이용을 위해서는 우선 해면 공생 미생물의 군집구조의 분석과 숙주 특이성 등 공생관계의 안정 성 측면에서 연구가 필요하며, 특히 해면 공생세균 군집의 다 양성에 관한 연구는 해면 공생세균의 특성 및 생태적 기능의 규명, 그리고 해면 공생 세균의 산업적 응용에 있어 매우 중요 하다(16). 이에 따라 해면 공생미생물의 다양성 및 군집구조에
관한 연구가 매우 증가하고 있는 추세이다(8, 10, 13). 해면 공 생미생물 중 가장 많은 비율을 차지하는 미생물은 세균으로, 해면 공생세균은 해면 생체량의 40% 이상에 달하는 것으로 알 려져 있다(2). 따라서 해면 공생세균의 군집구조의 분석은 해 면에 공생하는 전체 미생물의 군집을 평가하는데 있어 매우 중 요하다(15).
일반적으로 자연계의 미생물 중 실제 배양 가능한 종류는 전체 자연계 미생물 군집의 1% 이하인 것으로 알려져 있어 (12), 미생물의 군집구조의 분석에는 배양방법(culture-depen- dent)보다 비배양(culture-independent)에 근거하는 분자생물학 적 방법들이 많이 이용되고 있다. 즉 자연계로부터 배양을 거 치지 않고 직접 DNA를 분리하고, 이로부터 현재 미생물 분류 의 토대가 되고 있는 16S ribosomal RNA 유전자(16S rDNA) 를 확보하여 시료 내 종 다양성을 연구하는 방법들이 이용되고 있다. 이러한 방법에는 DGGE (denaturing gradient gel electrophoresis), FISH (fluorescence in situ hybrydization), ARDRA (amplified rDNA restriction analysis) 등이 있으며, 특히, 16S rDNA 염기서열 분석에 기초한 해면 공생세균의 군 집구조 분석에서 기존의 배양법에 의해 규명되지 못하였던 대 규모의 배양되지 않은 세균의 다양성이 밝혀지고 있다(2, 5).
본 연구는 남태평양에 위치한 미크로네시아의 축(Chuuk)에 서 2010년 2월에 채집한 해양 해면 Hyrtios sp. 604와 Callyspongia sp. 612의 공생 세균의 주요 군집구조를 비배양 법(culture-independent methods)에 기초한 16S rDNA-DGGE (denaturing gradient gel electrophoresis) 방법을 이용하여 비 교⋅분석하였다. 본 연구에서 다루고 있는 Hyrtios sp. 604와 Callyspongia sp. 612는 모두 Demospongiae에 속하는 해면으 로 Hyrtios의 경우, Callyspongia와 달리 공생하는 미생물들에 의해 면역억제제, 항암제 항진균제 등 다양한 생리활성물질들 을 생산하는 것으로 보고되어 있다(6, 20). 본 연구에서는 생화 학적 특성이 다른 두 종의 열대 해양 해면이 동일 지역에 서식 하는 경우에 있어 Hyrtios와 Callyspongia, 공생세균 군집 구조 의 차이를 파악하고자 하였다.
재료 및 방법
해면의 채집
열대의 해양 해면 Hyrtios sp. 604와 Callyspongia sp. 612 에 공생하는 세균의 다양성 조사를 위하여 2010년 2월, 남태평 양에 위치한 마이크로네시아의 축 주에서 스쿠버 다이빙을 이 용하여 25 m 깊이의 바다에서 채집하였다. 채집한 해면은 멸 균된 인공해수(ASW)로 3회 세척 후 -20°C에서 동결하여 운반 하였다.
Total genomic DNA 추출
채집된 해면은 -70°C에서 동결 후 동결건조기에서 -50°C, 0.033 M bar 기압으로 14시간 건조하였다. 동결 건조된 해면 은 멸균된 막자 사발에 넣고 액체질소를 부은 후 분쇄하였으며 CTAB 방법을 이용하여 total genomic DNA를 추출하였다(2).
1.5 ml tube의 100 μl선까지 분쇄된 시료를 넣고 CTAB buffer 800 μl을 첨가하여 섞은 후 동결(-70°C)과 융해(60°C)를 각각 10분씩 5회 반복 한 후, 50 μl proteinase K (10 mg/ml, Bio- sesang, Korea)와 50 μl lysozyme (50 mg/ml, Sigma, USA)을 첨가하여 37°C에서 2시간 처리하였다. 이어 20% SDS를 0.1 ml을 첨가한 후 65°C에서 1시간 처리하고 0.6배의 Phenol:
Chroloform: Isoamylalcohol (25:24:1, Sigma, USA)을 넣고 12,000 rpm에서 5분간 원심 분리하여 상등액을 수거하고 이 과정을 2회 반복하였다. 수거된 상등액에 0.54배의 isopropanol 을 첨가하여 냉동고에 15분간 방치 후 12,000 rpm에서 10분간 원심 분리한 후 상등액은 버리고 침전물을 얻었다. 여기에 70% 에탄올 500 μl를 넣어 세척하고 같은 조건으로 원심 분리 하여 상등액을 제거한 후 공기 중에서 건조하고 50 μl의 증류 수에 녹여 DGGE를 위한 PCR 반응의 주형으로 사용하였다.
16S rDNA의 DGGE-PCR 증폭
GC clamp가 추가된 341f와 518r primer쌍을 사용하여 16S rRNA gene의 V3 영역을 증폭하였다. PCR 반응은 GeneAmp PCR system 2700 thermal cycler (Applied Biosystems, Version 2.0, USA)를 이용하였다. PCR 반응 혼합물의 조성은 5 μl의 10× reaction buffer, 1 μl의 dNTPs, 5 unit/μl Taq polymerase (Solgent, Korea), 각각 10 pmol primer, 그리고 30 ng의 시료 DNA를 1 μl 첨가하여 최종부피가 50 μl가 되도 록 하였다. PCR 조건은 94°C에서 3분간 초기 변성시킨 후, 94°C에서 40초간 변성, 65°C (1 cycle당 0.5°C touch down) 에서 40초간 냉각, 72°C에서 1분간 신장, 이 과정을 30 cycle 반복 수행한 후 최종적으로 72°C에서 10분간 신장시켰다. 증 폭된 DNA의 확인을 위해서 PCR 반응액 2 μl를 취하여 2%
agarose gel (Bio-Rad, USA)을 이용하여 Mupid-ex (ADVANCE, Japan)로 100 V에서 25분간 1× TAE buffer (40 mM Tris, 20 mM acetic acid, 50 mM EDTA, pH 8.0)로 전기영동 하였다.
전기영동 후 Ethidium Bromide (50 ng/ml)로 염색하여 Gel Logic 200 (Kodak, USA)을 이용, 자외선 조사로 확인하였다.
증폭된 DNA의 크기를 확인하기 위한 marker로는 100 bp ladder (Intron, Korea)를 사용하였다.
DGGE 전기영동
DGGE는 Bio-Rad Dcode system (Bio-Rad, USA)을 이용 하여 수행하였다(12). 증폭된 PCR 산물은 bis acrylamide (Bio-Rad)를 포함한 10% polyacrylamide를 이용하여 35%에 서 65%의 농도 구배 조성(7 M urea, 40% formamide)으로 전 기영동을 수행하였다. Polyacrylamide gel은 크기 20×13 (W×H cm), 두께 1 mm로 작성하고 PCR 시료는 2× loading dye와 혼 합하여 40 μl의 시료를 loading하였다. 1× TAE buffer를 사용 하여 60°C, 30 V에서 30분간 안정화한 후 60 V로 전압을 올 려 16시간 전기영동을 수행하였다. 전기영동 후 DGGE gel은 Ethidium bromide로 1시간 염색 후 Gel Logic 200 (Kodak, USA)을 이용, 자외선 조사로 확인하였다.
H C
Fig. 1. DGGE banding patterns of amplified 16S rDNA obtained from Hyrtios sp. 604 (H) and Callyspongia sp. 612 (C).
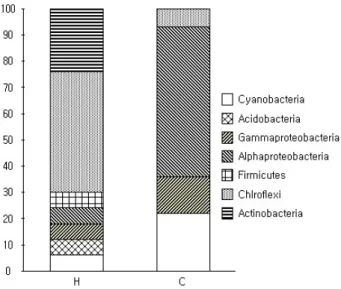
Fig. 2. Comparison of bacterial community in Hyrtios sp. 604 (H) and Callyspongia sp. 612 (C) based on 16S rDNA-V3.
염기서열 분석 및 계통학적 분석
DGGE로부터 확인된 각각의 band는 적출하여 50 μl의 증류 수로 DNA를 추출한 후 이것을 PCR 반응의 주형으로 GC가 제거된 341f와 518r primer 쌍을 사용하여 DGGE-PCR과 동 일한 조건에서 PCR을 수행하였다. PCR 증폭 산물은 PCR- purification Kit (Solgent, Korea)를 이용하여 정제한 후 염기 서열 분석(Macrogen, Korea)을 의뢰하였다. 염기서열은 DNAstar program을 이용하여 편집하였으며 National Center for Bio- technology Information (NCBI)의 Basic Local Alignment Search Tool (BLAST)을 이용하여 염기서열의 상동성을 검색 하였다. 계통수는 Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0의 neighbor-joining 방법을 이용 하였고, 1,000회 반복 bootstrap 분석에 의해 계통수의 안정성 을 조사하였다.
결과 및 고찰
DGGE band 양상
미크로네시아 축(Chuuk)의 동일 지역에 서식하는 두 종의 해양 해면 Hyrtios sp. 604와 Callyspongia sp. 612의 해면 공 생세균의 계통학적 다양성을 알아보기 위하여 비배양 지문법 인 16S rDNA의 PCR-DGGE를 수행하였다. 해면 시료로부터 추출한 gDNA를 주형으로 16S rRNA gene의 V3 영역을 증폭 한 결과, 194 bp의 예상된 크기의 PCR 산물을 얻었다. 두 해 면 종의 공생세균의 DGGE band 패턴 분석 결과, 해면 Hyrtios sp. 604에서 17개, Callyspongia sp. 612에서 14개의 band를 확인할 수 있었으며 Hyrtios sp. 604와 Callyspongia sp. 612의 주요 DGGE band들은 각각의 해면에 특징적인 것
으로 나타났다(Fig. 1). 두 해면 종의 주요 DGGE band들이 나 타내는 세균 군집 다양성을 파악하기 위하여 16S rDNA의 V3 영역의 부분 염기서열을 분석하였다.
염기서열 분석 및 계통학적 분석
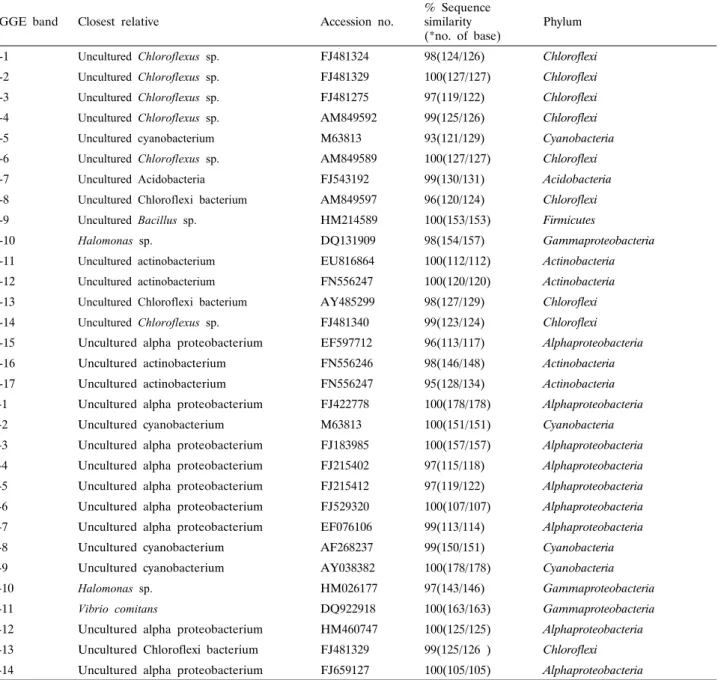
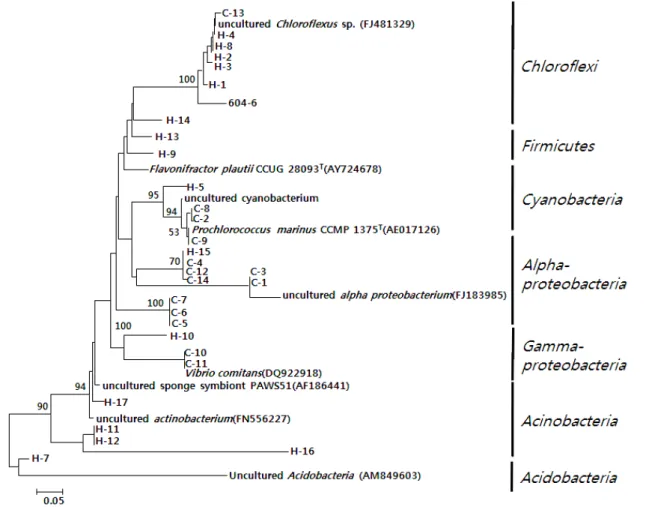
세균 군집의 다양성을 분석하기 위하여 DGGE band로부터 DNA를 추출하여 341f와 518r을 이용하여 재증폭하여 염기서 열을 분석하였다. 적출된 DGGE band에 대하여 염기 서열을 결정한 결과, Table 1에 보는 바와 같이 Hyrtios sp. 604에서 17개, Callyspongia sp. 612에서 14개, 총 31개의 주요 band들 에 대하여 부분 염기서열을 확인할 수 있었다. 이 서열들에 대 하여 Blast search를 통하여 기존의 보고된 서열들과 상동성 (similarity)을 검색하였다. 각각의 DGGE band에서 유래한 염 기서열을 이용하여 군집구조를 분석하고(Fig. 2), 계통수를 작 성한 결과는 Fig. 3과 같다.
두 해면의 DGGE band로부터 결정된 서열들은 기존의 알려 진 서열들과 94%에서 100%의 상동성을 나타내었다. DGGE band의 패턴은 다양하였으나, band로부터 밝혀진 대부분의 서 열들은 배양되지 않은 세균 클론(uncultured bacterial clone)들 과 높은 상동성을 나타내었다. 일반적으로 자연계의 미생물 중 실제 배양 가능한 종은 전체 자연계 미생물 군집의 1% 이하인 것으로 알려진 바와 같이 본 연구에서 조사된 해면에 존재하는 세균 군집 역시 배양되지 않는 세균 군집이 높은 비율로 존재 함을 알 수 있었다.
Hyrtios sp. 604의 주요 공생세균 그룹은 Proteobacteria (Alphaproteobacteria, Gammaproteobacteria) Acidobacteria, Actinobacteria, Chloroflexi, Cyanobacteria, Firmicute의 6개 의 문에 속하는 다양한 세균 그룹을 나타내었다. Hyrtios sp.
604의 주요 우점 공생세균 그룹은 uncultured Chloroflexi와 Actinobacteria로 밝혀졌으며, 기존의 서열과 96%에서 100%
의 상동성을 나타내었다.
Table 1. Phylogenetic affiliation of re-amplified denaturing gradient gel electrophoresis (DGGE) bands derived from the sponges
DGGE band Closest relative Accession no. % Sequence
similarity
(*no. of base) Phylum
H-1 Uncultured Chloroflexus sp. FJ481324 98(124/126) Chloroflexi
H-2 Uncultured Chloroflexus sp. FJ481329 100(127/127) Chloroflexi
H-3 Uncultured Chloroflexus sp. FJ481275 97(119/122) Chloroflexi
H-4 Uncultured Chloroflexus sp. AM849592 99(125/126) Chloroflexi
H-5 Uncultured cyanobacterium M63813 93(121/129) Cyanobacteria
H-6 Uncultured Chloroflexus sp. AM849589 100(127/127) Chloroflexi
H-7 Uncultured Acidobacteria FJ543192 99(130/131) Acidobacteria
H-8 Uncultured Chloroflexi bacterium AM849597 96(120/124) Chloroflexi
H-9 Uncultured Bacillus sp. HM214589 100(153/153) Firmicutes
H-10 Halomonas sp. DQ131909 98(154/157) Gammaproteobacteria
H-11 Uncultured actinobacterium EU816864 100(112/112) Actinobacteria
H-12 Uncultured actinobacterium FN556247 100(120/120) Actinobacteria
H-13 Uncultured Chloroflexi bacterium AY485299 98(127/129) Chloroflexi
H-14 Uncultured Chloroflexus sp. FJ481340 99(123/124) Chloroflexi
H-15 Uncultured alpha proteobacterium EF597712 96(113/117) Alphaproteobacteria
H-16 Uncultured actinobacterium FN556246 98(146/148) Actinobacteria
H-17 Uncultured actinobacterium FN556247 95(128/134) Actinobacteria
C-1 Uncultured alpha proteobacterium FJ422778 100(178/178) Alphaproteobacteria
C-2 Uncultured cyanobacterium M63813 100(151/151) Cyanobacteria
C-3 Uncultured alpha proteobacterium FJ183985 100(157/157) Alphaproteobacteria C-4 Uncultured alpha proteobacterium FJ215402 97(115/118) Alphaproteobacteria C-5 Uncultured alpha proteobacterium FJ215412 97(119/122) Alphaproteobacteria C-6 Uncultured alpha proteobacterium FJ529320 100(107/107) Alphaproteobacteria C-7 Uncultured alpha proteobacterium EF076106 99(113/114) Alphaproteobacteria
C-8 Uncultured cyanobacterium AF268237 99(150/151) Cyanobacteria
C-9 Uncultured cyanobacterium AY038382 100(178/178) Cyanobacteria
C-10 Halomonas sp. HM026177 97(143/146) Gammaproteobacteria
C-11 Vibrio comitans DQ922918 100(163/163) Gammaproteobacteria
C-12 Uncultured alpha proteobacterium HM460747 100(125/125) Alphaproteobacteria C-13 Uncultured Chloroflexi bacterium FJ481329 99(125/126 ) Chloroflexi C-14 Uncultured alpha proteobacterium FJ659127 100(105/105) Alphaproteobacteria
* The numbers in parentheses are the part of the total bases used to calculate the levels of sequences similarity. H, Hyrtios sp. 604 ; C, Callyspongia sp.
612.
DGGE band H-1, H-2, H-4, H-6 그리고 H-14는 uncultured Chloroflexus sp. (FJ481324, FJ481329, FJ481340)와 98%에 서 100%의 상동성을 나타내었으며 H-3과 H-8, 그리고 H-13 은 uncultured Chloroflexi bacterium (DQ661857, AM849597, AY485299)과 96%에서 100%의 상동성을 나타내었다. DGGE band H-11, H-12, H-16 그리고 H-17은 uncultured actino- bacterium (GQ412829, FN556227, FN556246, AJ347072)과 98%에서 100%의 상동성을 나타내었다. DGGE band H-5는 Uncultured cyanobacterium (M63813)과 94%, H-7은 Uncultured Acidobacteria (AM849603)와 95%, H-10은 Halomonas halophila (GU001901)와 96%, 그리고 H-15는 Uncultured alphaproteo-
bacterium (FJ183985)과 96%의 상동성을 각각 나타내었다.
Callyspongia sp. 612의 주요 공생세균은 Proteobacteria (Alphaproteobacteria, Gammaproteobacteria), Chloroflexi, Cyano- bacteria의 3개의 문에 속하는 세균 그룹으로 나타났으며, 기존 의 서열과 94%에서 100%의 상동성을 보였다. Callyspongia sp. 612의 주요 우점하는 공생세균 그룹은 Alphaproteobacteria 로 나타났다.
DGGE band C-1, C-3 C-4, C-5 C-6, C-7 C-12 그리고 C-14는 uncultured Alphaproteobacteria (FJ422778, GQ337224, FJ422768, GQ342231, FN435566, FJ800262, HM128528, HM036451, FN435363)와 각각 96% 이상의 상동성을 나타내
Fig. 3. Phylogenetic tree based on 16S rDNA sequences retrieved from the DGGE bands of the sponges, Hyrtios sp. 604 and Callyspongia sp.
612 and Numbers above branches indicate bootstrap values of neighbor-joining analysis (>50%) from 1,000 replicates. The scale bar represents 0.05 substitution per nucleotide position.
었으며, C-2, C-8과 C-9는 uncultured cyanobacterium (GU461222, HM128528, GQ480671)과 94%에서 97%의 상동성을 나타내 었다. C-10은 Halomonas sp. (HM026177)와 98%, C-11은 Vibrio comitans (DQ922918)와 98%, C-13은 Uncultured Chloroflexi (EF076104)과 98%의 상동성을 각각 나타내었다 (Table 1).
16S rRNA gene 염기서열로부터 계통학적 다양성을 분석한 결과(Fig. 2), 두 종의 해면에서 나타난 전체 세균 군집 구조는 Hyrtios sp. 604에서 Chloroflexi, Actinobacteria를 포함하여 6 개의 문, Callyspongia sp. 612의 경우, 가장 일반적인 해면 공 생세균 그룹으로 알려진 Proteobacteria (Alpha-, Gamma-)를 포함하여 3개의 문(phylum)에 속하는 세균 군집 구조를 나타 내었다. 중국 남지나해의 Selletta tenui와 Halicondria sp. 해면 의 경우 본 연구와 동일한 DGGE 방법에 의해 공생세균 다양 성을 조사한 결과 Proteobacteria 하나의 세균 문만이 나타나 는 것으로 보고하고 있어(9) 이 결과와 비교하여 볼 때 본 연 구에 사용한 두 해면 공생세균의 군집은 매우 다양한 것으로 파악된다.
한편 두 종의 해면에서 공통되는 공생세균 그룹은 Proteo-
bacteria (Alpha-, Gamma-), Chloroflexi, Cyanobacteria로 나 타났으며, 이 세균 그룹들은 16S rRNA gene의 분석에 의해 지금까지 밝혀진 해면공생세균 15개 문(phylum) 중에서 가장 전형적인 세균으로 알려져 있는(4, 12) 그룹이다. 두 해면 종은 공통적으로 위의 3문의 공생세균 그룹을 포함하고 있으나 Hyrtios sp. 604 해면의 경우, 공통의 세균 그룹 이외에 Acido- bacteria, Actinobacteria, Firmicutes의 세균 그룹을 더 포함하 였다. 또한 공생세균 중 우점하는 세균그룹에 있어 Hyrtios sp.
604의 경우 Chloroflexi와 Actinobacteria, Callyspongia sp.
612의 경우 Alphaproteobacteria와 Cyanobacteria가 주요 그 룹으로 관찰되어 두 종의 해면은 서로 다른 공생세균 군집구조 를 갖는 것으로 나타났으며(Figs. 2, 3) 이는 해면 종에 따라 세균 군집 구조가 서로 다르다는 연구 결과들(8, 9, 18)과 일치 하는 결과로써 해면 공생세균의 군집 구조는 숙주 특이적(host- specific)인 것으로 판단된다.
한편 동일 종의 해면일지라도 온대와 열대의 지리적 분포에 따라 세균군집이 다르거나(16), 혹은 계통적 유연관계가 멀며 지리적 분포가 다른 해면에서도 유사한 세균군집을 갖는다는 (7) 보고 이외에도, 공생세균에 따라 소수의 해면 종에 선택적
으로 공생하거나 혹은 어떤 공생세균은 숙주 특이성이 비교적 미약하여 다양한 해면 종에 공생하는 것들도 알려져 있어(15) 숙주 특이성에 관한 근거는 아직 명확히 밝혀져 있지 않다. 해 면동물과 공생하는 세균의 기원은 여과섭식 시 주변 환경으로 부터 유래한 미생물의 선택적 흡착에 의하거나 혹은 생식세포 를 통한 공생세균의 수직적 전달에 의한 것으로(14) 공생세균 의 숙주 특이성 혹은 숙주 특이적 공생세균의 규명은 해면의 종류 및 분포에 따른 생태학적 특성뿐 만 아니라 발생학적 측 면에서도 연구가 이루어져야 할 것으로 사료된다.
본 연구에 이용된 Hyrtios sp. 604는 다양한 이차대사산물을 생산하는 것으로 알려진 Hyrtios 속 해면으로써 Callyspongia sp. 612에 비해 훨씬 높은 공생세균 다양성을 나타내었다(Fig.
2). 이는 이차대사산물을 생산하는 것으로 보고된 바(1) 있는 Hyrtios erectus의 경우 Amphimedon sp.와의 비교 연구에서 동일한 지역에 서식하는 해면 종이지만 두 종에서 나타나는 세 균 군집이 서로 다르고 Amphimedon sp.에 비해 세균 군집의 다양성이 더 높다는 보고(13)와 일치하는 결과이다. 또한 Hyrtios sp. 604의 공생세균 그룹에서 Callyspongia sp. 612에 서 나타나지 않은 Actinobacteria가 우점 세균그룹으로 나타났 다. 이 결과는 지금까지 밝혀진 배양(culture- dependent) 혹은 비배양(culture-independent) 법에 의해 밝혀진 해면 공생세균 군집에서 Alphaproteobacteria와 Gammaproteobacteria 일반 적으로 우점하는 것으로 보고되어 있으나(1, 7), 임상적으로 중 요한 대사산물을 생산하는 해면의 공생세균 그룹에서는 Actino- bacteria의 비율이 높은 것으로 밝혀진 연구 결과들과(3, 19) 유사한 결과이다.
본 연구에서 남태평양의 미크로네시아의 축에 서식하는 두 종의 해면을 같은 시기에 동일 지역에서 채집하여 DGGE fingerprint를 이용하여 공생세균 군집의 다양성을 살펴본 결과, Hyrtios sp. 604와 Callyspongia sp. 612의 공생세균 군집은 Alphaproteobacteria, Gammaproteobacteria, Chloroflexi, 그리 고 Cyanobacteria를 공통의 세균 군집으로 포함하였으며, 이차 대사산물을 생산하는 것으로 알려진 Hyrtios 속의 해면 Hyrtios sp. 604는 공통의 공생세균 그룹 외에 Actinobacteria, Acidobacteria, Firmicutes를 포함하여 훨씬 다양한 세균 종을 함유하는 것으로 나타났으며 Actinbacteria를 우점 세균 그룹 중의 하나로 포함하였다. 이는 동일지역에 서식하는 두 해면 종에서 서로 다른 공생세균 군집을 나타내어 숙주 특이적 분포 를 보이는 것으로 해면 종에 따른 해양 환경에서의 생태적 기 능의 결과에 의한 것으로 생각된다.
공생세균 군집 구조의 파악은 해면으로부터 이차대사산물을 안정적으로 생산하기 위한 선행 연구로써 다양한 해면을 대상 으로 활발히 이루어지고 있으나 특히 숙주 특이성에 관한 연구 는 해면 종에 따른 이차대사산물의 특성과 숙주 특이적 공생세 균과의 상호 연관성을 파악하는데 중요하다.
본 연구에서 대상으로 한 Hyrtios 속 해면의 경우 이차대사 산물의 생산자로 알려져 있어 이차대사산물을 생산하는 것으 로 알려진 바 없는 Callyspongia sp. 612와의 공생세균 군집구 조의 비교는 공생세균의 산업적 이용의 기초자료로 활용할 수
있을 것으로 기대된다. 지금까지의 연구 결과로는 숙주 특이성 뿐만 아니라 지역적 특이성에 관한 보고들도 이루어 지고 있어 (2, 12) 숙주 특이성에 관하여는 더 많은 연구 결과의 축적이 필요하며, 해면 공생세균의 다양성 및 숙주 특이성 외에 공생 세균의 전이 모델, 숙주와의 상호작용 등, 광범위한 연구가 이 루어져야 할 것이다. 또한 공생 세균의 다양성에 관한 연구에 서 가장 많이 이용되는 16S rRNA PCR-DGGE 방법은 비교적 신속하게 해면 공생세균의 군집구조를 분석할 수 있는 효과적 인 방법이나 DNA 추출 방법에 따라 우점 그룹이 달라진다는 연구 결과도 제시되어 있어(4) 연구 방법에 따른 차이를 고려 하면서 해면 공생세균의 다양성 및 숙주 특이성 등의 연구가 수행되어야 할 것으로 사료된다.
적요
남태평양에 서식하는 두 종의 해면 Hyrtios sp. 604와 Callyspongia sp. 612로부터 16S rDNA DGGE 방법을 이용하 여 공생세균 군집의 다양성을 분석하였다. DGGE band로부터 염기서열 분석 결과, Hytrios sp. 604의 공생세균은 Alpha- proteobacteria, Gammaproteobacteria, Acidobacteria, Actino- bacteria, Chloroflexi, Cyanobacteria, Firmicutes로 나타났으 며, Callyspongia sp. 612의 공생세균 그룹은 Alphaproteo- bacteria, Gammaproteobacteria, Chloroflexi, Cyanobacteria로 나타났다. 풍부한 이차대사산물의 생산자로 보고된 Hyrtios 해 면 속의 Hyrtios sp. 604는 Callyspongia sp. 612에 비해 더 다 양한 공생세균 군집을 나타내었으며 주요 공생세균 군집으로 Actionobacteria가 포함되었다. 동일 지역에 서식하나 화학적 특성이 다른 두 해면 종의 세균 군집은 서로 다른 것으로 나타 났다. 발견된 공생세균의 염기서열은 90% 이상이 uncultured clone들과 높은 상동성을 나타내었다.
감사의 말
이 논문은 2009년 정부(국토해양부)의 재원으로 한국해양수 산기술진흥원의 지원을 받아 수행된 연구(PJT200096)이며, 이 에 감사드립니다.
참고문헌