155
16S rDNA-RFLP에 의한 Spirastrella abata와 Spirastrella panis 해면에 서식하는 배양가능한 공생세균 군집의 비교
조현희·박진숙*
한남대학교 생명공학과
계통적으로근연하며지리적분포가유사한두종의
Spirastrella
속의해양해면, S. panis
와S. abata
의배양가능한 공생세균군집구조를16S rDNA-RFLP
방법에의해분석하였다.
공생세균의배양은해면추출물3%
를포함하는MA
배지를사용하였다.
증폭된16S rDNA
의RFLP (restriction fragment length polymorphism)
분석을위한제한 효소로Hae III
와Msp I
을이용하였으며,
그결과24
개의RFLP type
을구별할수있었다.
각패턴별로1~5
개의분리균주를선별하여부분염기서열분석결과
,
알려진세균종과98.4%
이상의유사도를나타내었으며2
종의Spirastrella
해면으로부터 분리된 세균들은 모두Alphaproteobacteria , Gammaproteobacteria , Firmicutes , Actinobacteria 4
개의강(class)
에포함되었다. Alphaproteobacteria
는S. abata
에서39.3%, S. panis
에서47.6%
가관 찰되어두해면에서우점하는세균군집이었다. Gammaproteobacteria
의경우S. abata
에서38.5%
로관찰된반면S. panis
에서1.6%
의아주적은비율로관찰되었다.
또한Bacillus (phylum Firmicutes )
종은S. abata
에서9.7%
를 나타낸반면, S. panis
에서는44.3%
의분포를나타내었다. Planococcus maritimus (8.1%, phylum Firmicutes )
와Psychrobacter nivimari s (28.9%, phylum Gammaproteobacteria )
는S. abata
에서만관찰되어이들은S. abata
에특이 적인세균종임을알수있었다.
같은장소에서식하는계통적으로근연한두종의해면에서공생세균의군집구 조는차이가 큰것으로 나타났다.
Key words
□16S rDNA, community, culturable bacteria, marine sponge, RFLP, Spirastrella , sponge-associated bacteria
해양무척추동물은세포내혹은세포외에많은미생물을포함
하는것으로알려져있다
(12).
특히여과섭식하는해면은 세균,
고세균
,
시아노박테리아,
녹조류,
홍조류,
규조류등많은미생물 을포함하고있으며, Demosponges
종의mesohyl
은매우다양한생태적미소환경
(micro-environment)
을제공하여(22),
해면 생체량의
40%
이상에 달하는 세균을함유하기도 하는것으로 알려져 있으며
,
이러한 해면은bacteriosponges
혹은high-microbial abundance sponges (8)
라불리운다.
해면에서식하는공생미생물은해면의표면
,
혹은해면세포내,
혹은mesohyl
에서식하며일 시적인혹은영구적인공생관계를유지하는것으로알려져있다.
공생하는 미생물은
Pseudomonas denitificans
와 같이 여러 종의 해면에서우점종으로 발견되는경우(5, 14)
도있으나,
외부의환 경변화에도불구하고숙주해면과의공생관계가영구적인세균종이있다는보고
(6, 24)
도있다.
세균은여과섭식자인해면동물의먹이원이기도하지만
,
해면의면역반응에중요한역할을하기도한다
(21).
해면동물의소화과정과면역반응에저항하며성공적으로공생하는세균은이차대사산물의생합성에관련되어있 어생리활성물질의원생산자로지목되고있다
(7).
많은신규생리활성물질이 해면으로부터 분리되고있으며
(3, 4, 10, 17, 18),
본연구에서 다루고 있는
Spirastrella
로부터 항암제(1, 12),
콜 레스테롤합성저해제(19),
항생제(2)
등의생리활성물질들이생산 되는것으로보고되고있다.
이러한해면의생리활성물질들이공생하는 세균에의해생산되는것으로 추정되면서해면공생세균 의다양성및군집구조에관한연구가활발히수행되고있다
.
자연계미생물의대부분이실험실배양이불가능하므로최근자연 생태계의 군집구조를 밝히는데 분자적 방법
(DGGE, FISH,
rDNA
등)
들이적용되고있다.
그러나자연환경에서의미생물의생태적 역할의측정을위해서는 미생물의 배양과특성 파악이 이루어져야만가능하며
(20),
해면의배양없이천연물생산을위 한물질탐색과대량생산,
혹은2
차대사산물의생산력을분석하 기위해서는배양가능한해면공생세균에관한연구가필수적이다
(11).
따라서 해면공생세균의다양성을밝히는 데있어배양법에근거한연구도활발히이루어지고있다
(10, 13, 28, 30).
본연구에서는 다양한 천연물이 생산되고 있는 것으로 알려진
Spirastrella
속의 해면 중계통적으로 근연하며 지리적 분포가유사한것으로 알려진
S. abata
와S. panis
를우리나라의제주도로부터채집하여배양가능한
(culturable)
종속영양세균의공생세균군집구조를
16S rDNA (16S rRNA gene)
를PCR
에 의해증 폭하여RFLP (restriction fragment length polymorphism)
방법에 의해분석하였다.
*To whom correspondence should be addressed.
Tel: 82-42-629-8771, Fax: 82-42-629-8769
E-mail: [email protected]
재료 및 방법
해면의 채집 및 세균의 분리해면은
2007
년9
월28
일제주도모슬포항에서스쿠버다이빙을 이용하여
25 m
깊이의 바다에서 채집하였으며,
채집된Spirastrella abata
와Spirastrella panis
는해면조직이 공기와접촉되는 것을방지하기 위하여 해수가 포함된
plastic bags
에 옮겨4
oC
에서운반하였다.
해면조각을멸균된인공해수(ASW)
로3
회 세척후,
해면의 안쪽을1 cm
3 크기로잘라 인공해수3 ml
넣어균질화시킨 다음
10 min
간초음파 처리하였다.
각각10
-4까지 순차 희석한 후 각각의 해면 추출물3%
를 가한MA
배지(marine agar 2216, Difco, USA)
에100
µl
씩 도말하여30
oC
에서2
일간배양하였다.
각각의해면추출액은Wichels
등(27)
의방법 에따라준비하였다.
Spirastrella abata
에서303
개, Spirastrella panis
에서317
개의 균주를 분리하였다.
분리된 균주들의순수 분리를 위하여 동일 한천배지에서계대배양하여단일콜로니를얻었다.
두종의해 면에서 분리한 균주 중, Spirastrella abata
에서135
균주, Spirastrella panis
에서124
개의 균주를 무작위로 선별하여 총348
개의균주를이용해16S rDNA
의RFLP
분석을수행하였다.
DNA 추출 및 16S rDNA의 PCR 증폭
염색체
DNA
는 분리된 세균 균주의 각colony
로부터gDNA Isolation kit (Promega, USA)
를사용하여 분리하였으며 분리된DNA
는PCR
반응의주형으로사용하였다. 16S rDNA
의증폭에 는27f (AGA GTT TGA TCC TGG CTC AG)
와1492r (TAC GGY TAC CTT GTT ACG AC)
의primer
쌍을 사용하였다. PCR
반응은100 ng
의 주형DNA
와5U e-Taq polymerase (Solgent, Korea), 10 mM dNTP, 10
×e-Taq buffer, 10 pmol primer
를최종 반응량이50
µl
가되도록 혼합하여 수행하였다. GeneAmp PCR system 2700 thermal cycler (Applied Biosystems, Version 2.0, USA)
를이용하여94
oC
에서3
분간초기 변성시킨 후, 94
oC
에서40
초간 변성, 55
oC
에서40
초간 냉각, 72
oC
에서1
분간신장,
이과정을30 cycle
반복수행한후최종 적으로72
oC
에서10
분간신장시켰다.
증폭된DNA
의확인을위해서
PCR
반응액3
µl
를취하여1% agarose gel (Bio-Rad, USA)
을 이용하여Mupid-ex (ADVANCE, Japan)
로100 V, 25
분간1
×TAE buffer (40 mM Tris-acetate, 1 mM EDTA, pH 8.0)
에서전기 영동 하였다.
전기영동후, EtBr (ethidium bromide, 50 ng/ml)
에10
분간 염색하여Gel Logic 200 (Kodak, USA)
을이용하여UV
하에서약
1.5 kb
단편을확인하였다.
RFLP analysis
PCR
산물의RFLP
분석을위해증폭된1.5 kb
의DNA
단편에2
종의 제한효소Hae III (TaKaRa, Japan)
와Msp I (TaKaRa)
을사용하였다
. PCR
산물에 각각의제한 효소를 첨가하여37
oC
에서4
시간 반응시켰다.
반응물은3% agarose gel (Bio-Rad, USA)
을 사용하여1
×TAE buffer (40 mM Tris-acetate, 1 mM EDTA, pH
Fig. 1. The RFLP fingerprinting types of 16S rDNA amplification products of bacterial strains isolated from the marine sponges, S . abata digested with the restriction enzyme Hae III (A) and Msp I (B).
aindicates the strain number of bacterial isolates. M is 1 kb molecular weight marker (bp).
The lane number corresponds to the RFLP fingerprinting type listed in Table 1.
8.0)
로100 V, 30
분간전기 영동한후EtBr (50 ng/ml)
로염색하 여Gel Logic 200 (Kodak)
을이용하여UV
하에서관찰하여각 균주의밴드유형을확인하였다.
염기서열 분석 및 계통수 작성
16S rDNA-RFLP fingerprinting type
에 따라 각각의RFLP
type
별로1~5
개의 분리 균주들을 선택하여 총46
균주의 부분염기 서열
(900 bp
이상)
을 분석하였다.
증폭된PCR
산물은High Pure PCR Product Purification kit (Roche, IN)
를이용하여정제하였으며
, 27f primer
로ABI PRISM 3100 automated sequencer (PE Applied Biosystems, CA)
를 이용하여 염기 서열을 결정하였다
.
결정된 염기서열은NCBI (the National Center for Biotechnology Information)
에 등록된 염기서열을 대상으로Blast search
를 수행하였다.
각 염기서열의alignment
는CLUSTAL W (23)
를이용하여정렬하였고계통분석은PHYDIT
3.5 package (2)
를 이용하여neighbor-joining method
에 의해phylogenetic tree
를 추론하였다. 1,000
회 반복bootstrap
분석에 의해계통수를확인하였다.
결 과
RFLP 분석
두종의
Spirastrella
속의해면, S. panis
와S. abata
의배양가 능한 공생세균 군집구조를16S rDNA
의PCR-RFLP
방법에의 해분석하였다.
마쇄한해면조직액을초음파처리하고해면추출 액을첨가한배지에배양하였을경우,
그렇지않았을때와비교 하여 훨씬 많은수의 공생세균이 관찰되었으므로(
자료미제시)
이방법에따라공생세균을분리하여군집분석에이용하였다
.
두종의해면에서순수분리된총
620
개균주중무작위로선별된,
총
259
균주에대하여두종의제한효소를사용하여16S rDNA
의
RFLP
분석을 수행한 결과, 24
개의 서로 다른RFLP type
이 구분 되었다(Fig. 1 and Table 1). Hae III
에의해15 type (Table 1
의아라비아숫자), Msp I
에의해16
개의type (Table 1
의알파 벳)
을얻었으며이를근거로분석한 결과,
총24
개의서로다른RFLP type
을얻을수있었다.
염기서열 분석 및 계통수 작성
RFLP
분석에 의해24
개의type
으로 나뉜, 2
종의Spirastrella
해면으로부터분리된 세균들에대하여 각
RFLP type
별로1~5
개의분리균주를선별하여부분염기서열을분석하였으며
,
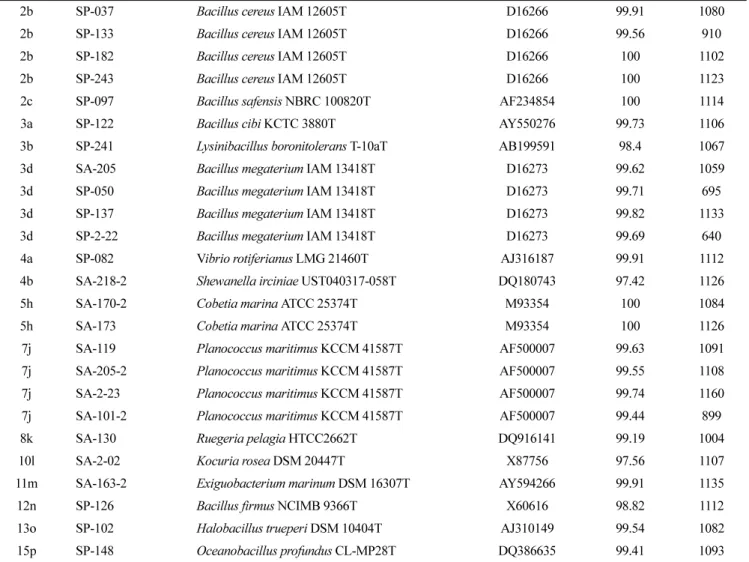
결정Table 1. RFLP fingerprinting types and sequence similarities of 16S rDNA from bacterial strains isolated from the marine sponges Spirastrella abata (SA) and Spirastrella panis (SP)
RFLP type
aStrain
bClosest species Accession no. Similarity (%) Length (bp)
1a SA-007 Pseudovibrio denitrificans JCM 12308T AY486423 99.63 1095
1a SA-2-14 Pseudovibrio denitrificans JCM 12308T AY486423 99.81 1077
1a SP-061 Pseudovibrio denitrificans JCM 12308T AY486423 99.82 1085
1a SP-064 Pseudovibrio denitrificans JCM 12308T AY486423 99.82 1097
1a SP-086 Pseudovibrio denitrificans JCM 12308T AY486423 99.74 777
1b SA-002 Psychrobacter nivimaris DSM 16093T AJ313425 99.37 1112
1b SA-031 Psychrobacter nivimaris DSM 16093T AJ313425 99.36 1093
1b SA-174 Psychrobacter nivimaris DSM 16093T AJ313425 99.36 1099
1c SA-107 Pseudovibrio denitrificans JCM 12308T AY486423 99.45 1085
1c SP-050 Pseudovibrio denitrificans JCM 12308T AY486423 98.76 888
1c SP-2-18 Pseudovibrio denitrificans JCM 12308T AY486423 98.74 1111
1d SA-2-07 Psychrobacter pacificensis IFO 16279T AB016057 99.81 1085
1d SP-052 Psychrobacter pacificensis IFO 16279T AB016057 99.55 670
1e SP-2-07 Pseudovibrio ascidiaceicola DSM 16392T AB175663 100 1098
1f SA-120 Psychrobacter celer KCTC 12313T AY842259 98.4 1123
1g SP-2-36 P seudovibrio japonicus WSF2T AB246748 99.91 1107
1i SP-234 Terribacillus halophilus 002-051T AB243849 100 1077
2a SP-185 Bacillus aerophilus JCM 13347T AJ831844 99.82 1084
2a SP-206 Bacillus aerophilus JCM 13347T AJ831844 99.82 1096
2a SP-2-06 Bacillus aerophilus JCM 13347T AJ831844 99.81 1074
2b SP-025 Bacillus cereus IAM 12605T D16266 100 332
된 부분 염기서열은
NCBI
에등록하였다(Table 1).
염기서열이 분석된 총46
개의 분리 균주는 모두 기존에 보고된 세균 종과98.4%
이상의 유사도를나타내었다.
염기서열이분석된 종들의동정결과와이들의
RFLP type
을근거로하여처음RFLP
분석 에쓰여 졌던259
개의 균주들을 분석하고이들의 분석 결과에 근거하여군집의 차이를 분석하였다.
이 결과는Table 2
와Fig.
2
에나타내었다.
두종의해면으로부터분리된
,
배양가능한공생세균은 모두Alphaproteobacteria , Gammaproteobacteria , Firmicutes , Actinobacteria 4
개의 강(class)
에 포함되었다. Table 2
에서 보는 바와 같이Alphaproteobacteria
는S. abata
에서39.3%, S. panis
에서47.6%
가관 찰되어 두 해면에서 우점하는 세균 군집이었다.
특히, Alphaproteobacteria
에서Pseudovibrio
종이두해면모두에서우점 종을차지하였다. Gammaproteobacteria
의경우S. panis
에서1.6%
의 아주적은비율로관찰된반면S. abata
에서38.5%
로관찰되었다.
그중에서
S. panis
에서관찰되지않았던Psychrobacter nivimaris
는S. abata
에서28.9%
로가장많은수를차지하여이종은S. abata
에 특이적인종임을알수있었다. Firmicutes
의경우, S . abata
에서는21.5%, S . panis
에서는49.2%
를차지하였다. S . abata
에서는Bacillus
속세균종들은
9.7%
로매우적은분포를나타낸반면, S . panis
해 면 종에서는Bacillus aerophilus , B. cereus , B. megaterium
등Bacillus
속세균종들이44.3%
를차지하여Bacillus
속세균종들이 가장우점하는세균종이었다. Planococcus maritimus
는S. panis
에서관찰되지않았으며
S . abata
에서만 발견되었다.
그구성비율은8.1%
를차지하여단일종으로는Firmicutes
에포함되는종에서가장 우점하는세균종이었다.
고 찰
24
개의RFLP type
별로1~5
개씩중복하여16S rDNA
의부분 염기서열(900 bp
이상)
을 분석한 결과 동일한RFLP type
은같은 종으로 동정되어
(Table 1), 2
종의 제한 효소에 의한RFLP
Table 1. RFLP fingerprinting types and sequence similarities of 16S rDNA from bacterial strains isolated from the marine sponges Spirastrella abata (SA) and Spirastrella panis (SP) (continued)
2b SP-037 Bacillus cereus IAM 12605T D16266 99.91 1080
2b SP-133 Bacillus cereus IAM 12605T D16266 99.56 910
2b SP-182 Bacillus cereus IAM 12605T D16266 100 1102
2b SP-243 Bacillus cereus IAM 12605T D16266 100 1123
2c SP-097 Bacillus safensis NBRC 100820T AF234854 100 1114
3a SP-122 Bacillus cibi KCTC 3880T AY550276 99.73 1106
3b SP-241 Lysinibacillus boronitolerans T-10aT AB199591 98.4 1067
3d SA-205 Bacillus megaterium IAM 13418T D16273 99.62 1059
3d SP-050 Bacillus megaterium IAM 13418T D16273 99.71 695
3d SP-137 Bacillus megaterium IAM 13418T D16273 99.82 1133
3d SP-2-22 Bacillus megaterium IAM 13418T D16273 99.69 640
4a SP-082 V ibrio rotiferianus LMG 21460T AJ316187 99.91 1112
4b SA-218-2 Shewanella irciniae UST040317-058T DQ180743 97.42 1126
5h SA-170-2 Cobetia marina ATCC 25374T M93354 100 1084
5h SA-173 Cobetia marina ATCC 25374T M93354 100 1126
7j SA-119 Planococcus maritimus KCCM 41587T AF500007 99.63 1091
7j SA-205-2 Planococcus maritimus KCCM 41587T AF500007 99.55 1108
7j SA-2-23 Planococcus maritimus KCCM 41587T AF500007 99.74 1160
7j SA-101-2 Planococcus maritimus KCCM 41587T AF500007 99.44 899
8k SA-130 Ruegeria pelagia HTCC2662T DQ916141 99.19 1004
10l SA-2-02 Kocuria rosea DSM 20447T X87756 97.56 1107
11m SA-163-2 Exiguobacterium marinum DSM 16307T AY594266 99.91 1135
12n SP-126 Bacillus firmus NCIMB 9366T X60616 98.82 1112
13o SP-102 Halobacillus trueperi DSM 10404T AJ310149 99.54 1082
15p SP-148 Oceanobacillus profundus CL-MP28T DQ386635 99.41 1093
a
Arabian number and alphabetic abbreviation present RFLP types with the restriction enzymes Hae III and Msp I, respectively
b
Strains used in phylogenetic tree constructed
type
은종을구별하기위한간단하고도안정적인수단임을확인할수있었다
.
문
(phylum)
수준에서두해면종의 공생세균은동일하게4
개의 강에 속하였으며
, S. abata
에서는Alphaproteobacteria (39.3%)
와Gammaproteobacteria (38.5%)
가 우점하는 반면, S.
panis
의 경우Firmicutes (49.2%)
와Alphaproteobacteria (47.6%)
가우점하였다
.
즉,
두종의해면에서공통된우점세균군집은Alphaproteobacteria
이었다.
또한Alphaproteobacteria
중단일종 으로가장우점하는세균은Pseudovibrio denitrificans
로S. panis
에서
42.7%, S. abata
에서37.8%,
각각분포하는것으로나타났다
.
이는Muscholl-Silberhorn
등(14)
에 의한 지중해에 서식하는해면
10
종에대한배양가능한 공생세균의다양성연구결과에 서Alphaproteobacteria
가 우점세균 군집이라는 것과 일치하는 결과였으며,
또한지중해 해면10
종중6
종의해면에서우점하 는세균종이Pseudovibrio denitrificans
라는연구결과매우유사 한결과였다.
남지나해로부터채집한4
종의해면에서배양가능한 주요우점세균군집에Alphaproteobacteria
가포함되며(13), 4
종의
Dictyoceratid
해면의 배양 가능한 주요 세균 군집이Alphaproteobacteria
라는연구결과(16)
와도일치하는것이다.
그 밖에Enticknap
등(5)
에의한연구 결과에의하면 서로다른해양
(
미국의Florida
와Maryland,
그리고Indonesia
및Jamaica
등)
에서식하는
7
종의해면으로부터분리한,
배양가능한공생세균군집에서 우점세균 군집이 모두
Alphaproteobacteria
이며 또한 우점세균종이Pseudovibrio denitrificans
라는결과와도일치하는 것이었다.
Gammaproteobacteria
의경우S. panis
에서는1.6%
밖에분포하 지 않으나, S. abata
에서는38.5%
로 나타나Alphaproteobacteria
에이어 우점세균 군집으로 나타났다
. Gammaproteobacteria
중Psychrobacter nivimaris
는가장 높은 분포율(28.9%)
을나타내는 세균종이나오직S. abata
해면에서만발견되어S. abata
에특이 적인세균종임을알수있었다.
Bacillus
속세균종의경우S . panis
해면에서는44.3%
를차지 하여 가장 우점하는 세균 종인데비해, S . abata
에서는9.7%
로매우 적은 분포를 나타내었다
.
반면Firmicutes
종 중Planococcus maritimus
는가장높은비율(8.1%)
로관찰되는세균종이었으나
S . abata
에서만관찰되었다.
그외분포비율은적으 나Exiguobacterium marinum , Ruegeria pelagia , Shewanella irciniae , Cobetia marina , Psychrobacter celer
역시S. abata
에서 만 관찰되었으며Bacillus cibi , B. firmus , Halobacillus trueperi , Lysinibacillus boronitolerans , Pseudovibrio ascidiaceicola
종은S.
panis
에서만관찰되었다.
본연구에 사용한
Spirastrella
두종의 해면은 계통적유연관계가가까우며
,
지리적분포가유사한것으로알려진해면으로써 제주도의모슬포항의같은장소에서채집된것으로써,
공생세균의군집은 문
(phylum)
수준에서Alphaproteobacteria
라는공통의 세균군집을 갖는것으로 나타났으나(Fig. 2),
세균종의구성에있어서는종특이적인분포를나타내었다
.
특히
Planococcus maritimus
와Psychrobacter nivimari s
의경우S. abata
에서만관찰되어이들은S. abata
에특이적인세균종임 을알수있었다. Turque
등(25)
의연구에서도해면종들은서로 공유하는 세균 그룹이외에도 해면 종에 특이적인세균 종들이 분포함을보고하고있다.
해면Rhopaloeides odorabile , 44
개체에 관한배양가능한공생세균에관한연구결과,
계절이나지리적분포에 관계없이 모든 개체에서
Alphaproteobacterium
(designated NW001)
이우점종이었으며(26),
이공생세균이 없는경우그해면의개체 수가감소하거나질병을 일으킨다는보고
(5)
를하여해면종에특이적인공생세균종이분포함을보고하였는데
,
본 연구에서도Psychrobacter nivimaris
와Planococcus maritimus
와같이Spirastrella
해면종에따른숙주특이성을갖Table 2. Relative abundance of species in major classes deduced from 16S rDNA sequences of bacteria associated with marine sponges, S.
abata and S. panis
Class S. abata S. panis
Species
Alphaproteobacteria 39.3
a47.6
Pseudovibrio ascidiaceicola 4.0
Pseudovibrio denitrificans 37.8 42.7
Pseudovibrio japonicus 0.8 0.9
Ruegeria pelagia 0.7
Gammaproteobacteria 38.5 1.6
Shewanella irciniae 1.5
Cobetia marina 2.2
Psychrobacter celer 3.7
Psychrobacter nivimaris 28.9
Psychrobacter pacificensis 0.7 0.8
Vibrio rotiferianus 1.5 0.8
Firmicutes 21.5 49.2
Bacillus aerophilus 3.0 16.9
Bacillus cereus 1.5 10.5
Bacillus cibi 0.8
Bacillus firmus 0.8
Bacillus megaterium 1.5 9.7
Bacillus safensis 3.7 5.6
Exiguobacterium marinum 2.3
Halobacillus trueperi 0.8
Lysinibacillus boronitolerans 1.6
Oceanobacillus profundus 0.7 0.8
Terribacillus halophilus 0.8 1.7
Planococcus maritimus 8.1
Actinobacteria 0.7* 1.6
Kocuria rosea 0.7 1.6
a
The number is percent (%) of each species in total bacterial commu-
nity
는공생세균들이존재하는것으로나타났다
. Zhang
등은5
종의해면으로부터분리한
,
배양가능한Actinobacteria
의다양성에관 한연구(29)
에서해면종에따라서로다른종구성을나타내는 것으로 보고하였으며, Ridley
등(16),
역시4
종의Dictyoceratid
해면에서
Alphaproteobacteria
는공통의우점종이지만지리적분 포와무관하게해면종에따라숙주특이적인세균군집이분포 함을보고하여본연구결과와유사한결과를나타내고있다.
한편
, Lafi
등(11)
은Great Barrier reef
로부터 채집한Pseudoceratina clavata
와Rhabdastella globostellata
의 해면에서배양 가능한세균의 군집을 조사한 결과
, Alphaproteobacteria
와Firmicutes
가 포함되어 있으나, Alphaproteobacteria
와Firmicutes
의 종구성은 속
(genus)
수준에서도서로 유사하다는,
본 연구결과와는 상이한 연구 결과를 보고하였다
. Hentscel
등(8)
의DGGE
와rpo gene
을이용한 비배양법에 의한연구 결과에서도유연관계가 멀며 지리적 분포가 서로 다른 두 종의 해면
Aplisina aerophoba
와Athenella swinhoei
의 공생미생물 군집이 거의 유사하다는연구 결과를 보고하여 해면공생세균은 숙주 특이성나타낸다는연구결과들과는상이한 결과를보고하였다.
Fig. 2. Neighbor-joining phylogenetic tree from analysis of 16S rDNA sequences (>900 bp) of isolated bacterial strains associated with S. abata
(SA) and S. panis (SP) . Numbers above branches indicate bootstrap values of neighbor-joining analysis (>50%) from 1,000 replicates. The scale
bar represents 0.1 substitutions per nucleotide position.
따라서공생세균의숙주특이성에관한연구는현재까지보고된 연구결과로는일관성을찾기어려우며
,
앞에서기술한바와같 이해면종에따라공생세균종의구성이매우 다른경우와그 와달리공생세균의종구성이매우유사하다는 상반된결과들 이보고되고 있어 해면공생세균의 숙주특이성에 관한 고찰은 다양한지리적분포를갖는많은해면종을대상으로이에관한 연구가더욱수행되어야할것으로생각된다.
Alphaproteobacteria
의경우서로다른다양한해면으로부터높 은빈도로발견되는배양가능한공생세균으로밝혀지고있는데,
이는각각의숙주해면과공진화해온것이아니며
,
진화과정에 서비교적늦게해면과 공생관계를구성한 것으로추정되고있다
(5).
이와같이해면에 따른공생세균의군집구조를 규명함으로써세균과해면의공생관계에따른생태적기능의분석및두 생물종간의진화적고찰이가능하리라생각된다
.
배양가능한공생세균의다양성에관한연구에서세균의순수 분리
,
특히해면 종에 따른고도의 해면 특이적인(host-specific)
세균종일경우
,
분리가 매우어려운것으로 알려져있으며(14),
분리를위한 배지와배양조건등에따라세균 종의구성이매
우달라지며
(13, 14, 29),
또한숙주와유사한영양조건을맞추더라도숙주내의환경을 재현하기가어렵고또공존하는다른미 생물과의상호작용 하에서만생장 가능한 것들도 있어
(9)
단일 균주로순수분리해내는방법만으로는실제배양가능한공생세균의군집구조의파악에는많은한계가있다
. MA
배지에해면추출물을첨가한 배지를 이용하는 경우
,
보다 다양한 공생세균군집이나타나지만영양농도가높은배지의경우해면추출물의
영향이거의없는것으로보고되고있어
(26),
현재까지의알려진배지를사용한 순수분리에 의한배양방법으로는배양가능한 공생세균중극히일부의세균군집만이파악가능하다
.
이러한 배양상의한계를극복하기위해다양한조성의배지를이용하여,
순수분리가아닌혼합배양에의해세균군집의구조를파악하 려는시도
(14)
도이루어지고있으나공생세균군집의농화배양을위한배지로우선해면추출물이나기타유사물질을이용하는방 안을포함하여배양조건에관한 더많은연구가 필요할것으로
생각된다
.
나아가metagenome
기법 혹은 분자생물학적 방법에의한연구결과와의비교도유용할것으로사료된다
. 감사의 말
이논문은
2005
년정부(
교육인적자원부)
의재원으로한국학술진흥재단의 지원을 받아 수행된 연구이며
(KRF-2005-204- C00070),
이에감사드립니다.
참고문헌
1. Alam, N., W. Wang, J.G. Hong, C.O. Lee, K.S. Im, and J.H. Jung.
2002. Cytotoxic sphingosine 4-Sulfates from the sponge Spiras- trella abata . J . Nat . Prod . 65, 944-945.
2. Chun, J. 1995. Computer-assisted classification and identification of actinomycetes. Ph.D. Thesis, Univ. Newcastle, Newcastle upon
Tyne, UK.
3. Dai
,J., Y. Liu, Y.D. Zhou, and D.G. Nagle. 2007. Hypoxia-selec- tive antitumor agents: norsesterterpene peroxides from the marine sponge Diacarnus levii preferentially suppress the growth of tumor cells under hypoxic conditions. J. Nat. Prod. 70, 130-133.
4. Dalisay, D.S. and T. Molinski. 2009. Structure elucidation at the nanomole scale. 2. Hemi-phorboxazole A from Phorbas sp. Org.
Lett. 11, 1967-1970.
5. Enticknap, J.J., M. Kelly, O. Peraud, and R.T. Hill. 2006. Charac- terization of a culturable alphaproteobacterial symbiont common to many marine sponges and dvidence for vertical transmission via sponge Larvae. Appl . Environ . Microbiol . 72, 3724-3732.
6. Friedrich, A.B., J. Hacker, I. Fischer, P. Proksch, and U. Hentschel.
2001. Temporal variations of the microbial community associated with the Mediterranean sponge Aplysina aerophoba . FEMS Microbiol . Ecol . 38, 105-113.
7. Guangyi, W. 2006. Diversity and biotechnological potential of the sponge-associated microbial consortia. J. Ind. Microbiol. Biotech- nol . 33, 545-551.
8. Hentschel, U., J. Hopke, M. Horn, A.B. Friedrich, M. Wagner, J.
Hacker, and B.S. Moore. 2002. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl . Environ . Microbiol . 68, 4431-4440.
9. Kaeberlein, T., K. Lewis, and S.S. Epstein. 2002. Isolating “ uncul- tivable ” microorganisms in pure culture in a simulated natural environment. Science 296, 1127-1129.
10. Kennedy, J., P. Baker, C. Piper, P.D. Cotter, M. Walsh, M.J. Mooij, M.B. Bourke, M.C. Rea, P.M. O’Connor, R.P. Ross, C. Hill, F.
O’Gara, J.R. Marchesi, and A.D.W. Dobson. 2009. Isolation and analysis of bacteria with antimicrobial activities from the marine sponge Haliclona simulans collected from Irish Waters. Mar. Bio- technol . 11, 384-396.
11. Lafi, F.F., M.J. Garson, and J.A. Fuerst. 2005. Culturable bacterial symbionts isolated from two distinct sponge species ( Pseudocera- tina clavata and Rhabdastrella globostellata ) from the great bar- rier reef display similar phylogenetic diversity. Microb. Ecol . 50, 213-220.
12. Levina, E.V., A.I. Kalinovsky, P.V. Andriyashenko, P.S. Dmi- trenok, D.L. Aminin, and V.A. Stonik. 2005. Phrygiasterol, a cyto- toxic cyclopropane-containing polyhydroxysteroid, and related compounds from the pacific starfish Hippasteria phrygiana . J . Nat . Prod . 68, 1541-1544.
13. Li, Z., L. He, and X. Miao. 2008. Cultivable bacterial community from South China Sea sponge as revealed by DGGE fingerprinting and 16S rDNA phylogenetic analysis. Curr. Microbiol . 55, 465- 14. Muscholl-Silberhorn, A., V. Thiel, and J.F. Imhoff. 2008. Abun- 472.
dance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microb . Ecol . 55, 94-106.
15. Park, S.H., K.K. Kwon, D.S. Lee, and H.K. Lee. 2002. Morpho- logical diversity of marine microorganisms on different media. J.
Microbiol. 40, 161-165.
16. Ridley, C.P., D.J. Faulkner, and M.G. Haygood. 2005. Investiga- tion of Oscillatoria spongeliae -dominated bacterial communities in four dictyoceratid sponges. Appl . Environ . Microbiol . 71, 7366- 7375.
17. Selvin, J. 2009. Exploring the antagonistic producer Streptomyces
MSI051: Implications of polyketide synthase gene type II and a
ubiquitous defense enzyme phospholipase A2 in the host sponge
Dendrilla nigra . Curr. Microbiol. 58, 459-463.
18. Selvin, J. and A.P. Lipton. 2004. Biopotentials of secondary metabolites isolated from marine sponges. Hydrobiologia 513, 231-234.
19. Shin, B.A., Y.R. Kim, I.S. Lee, C.K. Sung, J. Hong, C.J. Sim, K.S.
Im, and J.H. Jung. 1999. Lyso-PAF analogues and lysophosphati- dylcholines from the marine sponge Spirastrella abata as inhibi- tors of cholesterol biosynthesis. J . Nat . Prod . 62, 1554-1557.
20. Tamaki, H., Y. Sekiguchi, S. Hanada, K. Nakamura, N. Nomura, M. Matsumura, and Y. Kamagata. 2005. Comparative analysis of bacterial diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques.
Appl . Environ . Microbiol . 71, 2162-2169.
21. Thakur, N.L. and A.C. Anil. 2000. Antibacterial activity of the sponge Ircinia ramosa : Importance of its surface-associated bacte- ria. J. Chem. Ecol . 26, 57-71.
22. Thiel, V., S.C. Neulinger, T. Staufenberger, R. Schmaljohann, and J.F. Imhoff. 2007. Spatial distribution of sponge-associated bacte- ria in the Mediterranean sponge Tethya aurantium . FEMS Micro- biol. Ecol . 59, 47-63.
23. Thompson, J.D., D.G. Higgins, and T.J. Gibson. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap pen- alties and weight matrix choice. Nucleic Acids Res . 22, 4673-4680.
24. Thomson, C., M. Horn, W. Wagner, U. Hentschel, and P. Proksch.
2003. Monitoring microbial diversity and natural products profiles of the sponge Aplysina cavernicola following trasplantation. Mar .
Biol . 142, 685-692.
25. Turque, A.S., A.M. Cardoso, C.B. Silveira, R.P. Vieira, F.A.D. Fre- itas, R.M. Albano, A.M. Gonzalez, R. Paranhos, G. Muricy, and O.B. Martins. 2008. Bacterial communities of the marine sponges Hymeniacidon heliophila and Polymastia janeirensis and their environment in Rio de Janeiro, Brazil. Mar . Biol . 155, 135-146.
26. Webster, N.S., K.J. Wilson, L.L. Blackall, and R.T. Hill. 2001.
Phylogenetic diversity of bacteria associated with the marine sponge Rhopaloeides odorabile . Appl . Environ . Microbiol . 67, 434-444.
27. Wichels, A., S. Wurtz, H. Dopke, C. Schutt, and G. Gerdts. 2006.
Bacterial diversity in the breadcrumb sponge Halichondria pani- cea (Pallas). Microbiol . Ecol . 56, 102-118.
28. Zhang, H., Y.K. Lee, W. Zhang, and H.K. Lee. 2006. Culturable actinobacteria from the marine sponge Hymeniacidon perleve : iso- lation and phylogenetic diversity by 16S rRNA gene-RFLP anal- ysis. Antonie van Leeuwenhoek 90, 159-169.
29. Zhang, H., W. Zhang, Y. Jin, M. Jin, and X. Yu. 2008. A compar- ative study on the phylogenetic diversity of culturable actinobac- teria isolated from five marine sponge species. Antonie van Leeuwenhoek . 93, 241-248.
30. Zhu, P., Q. Li, and G. Wang. 2008. Unique microbial signatures of the alien hawaiian marine sponge Suberites zeteki . Microb . Ecol . 55, 406-414.
(Received May 27, 2009/Accepted June 22, 2009)
ABSTRACT: Comparative Analysis of the Community of Culturable Bacteria Associated with Sponges,
Spirastrella abata