Vibrio vulnificus is opportunistic pathogen naturally occurring in coastal area that can cause acute sepsis. In this study, 61 isolates were investigated the characteristics of strains such as pathogenicity and antibiotic susceptibility in the west coastal area of Gyeonggi-do from 2016 to 2020. Most strains showed proteolytic activity and hemolytic activity. Polymerase Chain Reaction (PCR) method was used to detect virulence genes such as vvhA (100%), glnA (100%), pilA (31.1%), viuB (42.6%), wza (42.6%), and rtxA (42.6%). The results of antibiotic sus- ceptibility tests to 27 different antibiotics presented 86.9~
100% resistance to 5 antibiotics including penicillin and 1.6~
55.7% intermediate resistance to 5 antibiotics including cefoxitin. Our findings revealed that Vibrio vulnificus isolated from environment may serve as potential human pathogen and one of strains showed intermediate resistance to chloramphenicol.
These findings from this study were judged to be useful guidelines for prevention and antibiotic treatment to Vibrio vulnificus sepsis.
Keywords: Vibrio vulnificus, antibiotic resistance, virulence genes
비브리오 패혈증균(Vibrio vulnificus)은 호염성, 운동성, 그 람음성 간균으로서(Janda et al., 1988) 비브리오콜레라균(Vibrio
cholerae), 장염비브리오균(Vibrio paraheamolyticus)과 함께 해양 환경에 광범위하게 분포하고 있다(Daniels, 2011). 비브 리오 패혈증은 비브리오 패혈증균 감염으로 발병하며 특히 만 성 간 질환 환자 또는 면역 저하 환자에게서 심각한 패혈증을 일으키는 것으로 알려져 있다(Shapiro et al., 1998). 국내에서 는 1982년 첫 비브리오 패혈증 환자가 보고되었으며(Goo et al., 1982), 최근 5년간 발생 현황을 보면 2015년 37명, 2016년 56명, 2017년 46명, 2018년 47명, 2019년 42명으로 매년 전국 적으로 50건 내외의 발생이 보고되고 있다(Fig. 1) (KDCA, 2020). 비브리오 패혈증으로 인한 치사율은 최소 21.4% (2016 년)에서 최대 52.2% (2017년)로 높게 나타나 보건위생상 주의 가 필요한 질환임을 알 수 있다(Fig. 1).
비브리오 패혈증균은 병원성 발현을 위해 hemolysin, RTX toxin 등을 생성하는데, 다양한 유전자가 병원성 조절에 관여 한다(Bang et al., 1999). 적혈구를 용혈시켜 균주의 철 사용을 돕는 hemolysin은 vvhA에 의해 조절되며(Zhang and Austin, 2005), RTX toxin은 rtxA이 관여하여 세포 괴사를 유발한다 (Kim et al., 2008). Vulnibactin은 비브리오 균주의 주요 독성 기전인 숙주로부터 철 획득에 관여하는 단백질로, viuB에 의해 조절된다(Ratledge and Dover, 2000). 이 밖에도 균주 외막의 지질 단백질 형성에 관여하는 wza (Collins and Derrick, 2007), 숙주의 상피세포에 균이 부착하도록 돕는 pilA (Paranjpye and Strom, 2005), 암모니아 대사효소인 Glutamine synthetase를 조절하는 glnA (Pesole et al., 1995) 등의 병원성과 관련된 유전 자가 알려져 있다.
경기 서해안에서 분리한 비브리오 패혈증균의 특성 연구
송지원* ・ 정일형 ・ 박신희 ・ 윤수정 ・ 허은선 ・ 김영숙 경기도보건환경연구원 수인성질환팀
Characterization of Vibrio vulnificus isolated from the west coastal area of Gyeonggi-do
Ji-Won Song* , Il-Hyung Jeong, Sin-Hee Park, Su-Jung Yun, Eun-Seon Hur, and Young-Sug Kim
Team of Waterborne Disease, Gyeonggi Province Institute of Health and Environment, Suwon 16381, Republic of Korea
(Received July 27, 2021; Revised September 8, 2021; Accepted September 9, 2021)
*For correspondence. E-mail: [email protected];
Tel.: +82-31-250-2585; Fax: +82-31-250-2527
최근 항생제 내성 균주가 의료계와 농·축·수산업계의 항생 제 오남용으로 인하여 환경으로 유입되고 있으며(Cabello et al., 2013), 식품 등을 통해 인체에 직접 전달되거나 인체 병원 균에 항생제 내성 유전자를 전달하는 방식으로 보건위생에 위 협이 되고 있다(Duran and Marshall, 2005). 비브리오균은 해 양 환경에 상재하며 오염된 해산물을 섭취하거나 상처가 오염 된 바닷물에 노출될 시 감염되는 것으로 알려져 있어(Kim and Rhee, 2003) 최근 비브리오균의 항생제 내성에 관한 다양한 연 구가 이루어지고 있다(Faja et al., 2019; Mohamad et al., 2019;
Valáriková et al., 2020). 비브리오 패혈증은 병의 진행속도가 매우 빠르고 병증 진행 후 48시간 이내 치사율이 50%에 이르 는 치명적 질환이므로 신속하고 적절한 항생제 투여가 매우 중요하며(Kim and Hong, 2012), 비브리오 패혈증의 치료에 적용되는 항생제로는 3세대 세팔로스포린계, 테트라사이클 린계, 플루오로퀴놀론계 또는 클로람페니콜계 항생제 등이 있 다(French et al., 1989).
재료 및 방법
균주 분리 및 특성 분류
2016년부터 2020년까지 서해안의 다섯 지역에서 분리한 비브리오 패혈증균 61균주를 사용하였다. 실험 대상으로 사 용된 균주의 월별 및 채취장소별 분포는 Table 1과 같다.
해수 500 ml를 여과한 여과막(0.45 µm) 또는 갯벌을 Alkaline 펩톤수(pH 8.6)에 접종하여 37°C에서 24시간 증균배양하였다.
증균액을 Thiosulfate Citrate Bile-salt Sucrose (TCBS, HAN- GANG) 배지에 도말하여 녹색 집락을 확인 후 CHROMagar- Vibrio (CHROMagar
TM) 배지에서 청록색을 보이는 집락을 선택 하여 1% NaCl이 첨가된 Tryptic Soy Agar (TSA, HANGANG) 배지에 계대하여 분리하였다. 최종적으로 VITEK2 (VITEK GN card, bioMérieux) 장비를 이용하여 생화학적 동정을 실시
하였다. 분리한 균주의 생물형 확인을 위해 인돌생성여부 확 인실험(Indole test, Kisan Bio)을 실시하였다.
배양 검사
각 균주의 용혈능 및 단백질 분해능 확인을 위해 배양검사를 실시하였다. 각 균주에 대한 용혈능 검사(Hemolysis assay)는 1%
NaCl을 첨가한 혈액 배지(5% Sheep blood agar, HANGANG)에 시험 균주를 접종하여 37°C에서 24시간 배양 후 용혈환의 생 성 유무로 용혈능을 판독하였다. 각 균주에 대한 단백질 분해 능 검사(Proteolysis assay)는 탈지유 고체배지(Skim Milk Agar, BD)를 제조하여 실시하였다. 탈지유 수용액과 한천 용액을 각 각 고압 멸균(121°C, 15분) 한 뒤 각각 최종농도가 1%, 1.5%가 되게 1:1로 조합하여 실온에서 굳혀서 사용하였다. 배지에 균 주를 접종하여 37°C에서 3일간 배양 후 균주 주변에 투명한 환 생성 유무로 단백질 분해능을 판독하였다.
독성 유전자 검출 검사
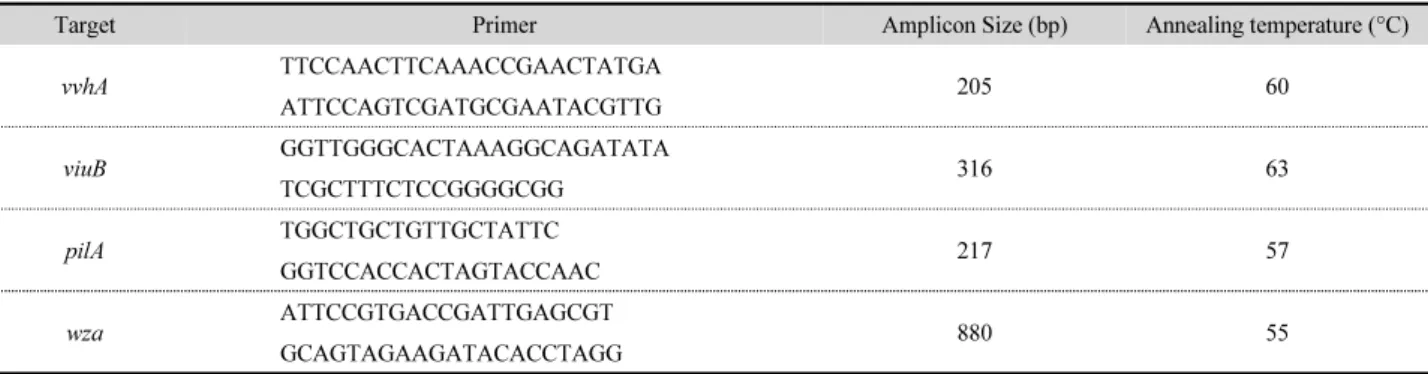
배양된 균주에서 QIAamp DNA Mini Kit (QIAGEN)를 사 용하여 제조사의 프로토콜에 따라 DNA를 추출한 뒤 Conven- tional-PCR법을 사용하여 독성유전자(vvhA, viuB, pilA, wza) 검출시험을 실시하였다. AccuPower HotStart PCR PreMx kit (Bioneer)를 사용하였으며, 각 primer 1 µl (1 ng/µl)와 template DNA 2 µl를 첨가하여 ProFlex 96 well (Life technologies) 기 기를 사용하여 PCR을 수행하였다. PCR 반응 조건은 94°C에 서 3분, 94°C에서 30초 denaturation, 각 primer에 맞는 온도에 서 30초 annealing, 72°C에서 30초 extension 과정을 30회 반복 하였으며, 최종 72°C에서 1분간 elongation 한 뒤 최종적으로 전기영동(QIAxcel, QIAGEN)을 통해 증폭산물을 확인하였
Fig. 1. Incidence of Vibrio vulnificus sepsis for 5 years in Korea.
Table 1. Monthly and areal distribution of Vibrio vulnificus isolated from
seawater and mud
Sampling area Source
of strains
Month
Total
4 5 6 7 8 9
Gimpo Seawater 3 5 3 - - 3 14
Mud - - - - 1 1 2
Siheung Seawater - 1 4 - 1 - 6
Mud - 1 - - 2 6 9
Ansan Seawater 1 - 2 2 - 1 6
Mud - - - - - - -
Hwaseong Seawater - - 4 - 2 1 7
Mud - - - - - - -
Pyeongtaek Seawater 3 3 - 1 1 1 9
Mud 1 - 1 4 2 8
다. 각 프라이머 서열 및 정보는 다음과 같다(Table 2). Real- time PCR 법을 사용하여 glnA와 rtxA 검출시험을 실시하였다.
PowerChek
TMVibrio vulnificus Real-time PCR Kit II (Kogene) 를 사용하였으며 제조사 프로토콜에 따라 실험을 실시하였다.
항생제 감수성 검사
항생제 감수성 검사를 위해 27종의 항생제에 대하여 액체배 지 미량희석법(Broth microdilution) 또는 VITEK2 AST-N169 test를 실시하였다. 비브리오 패혈증균 집락을 1% Nacl을 첨 가한 Muller Hinton Broth (MHB, Thermo)에 McFarland 탁도 0.5가 되도록 현탁한 뒤, TREK MIC plate (Sensititre MIC plate, Thermo)에 50 µl씩 접종하여 37°C에서 24시간 배양 후 각 항생제의 최소억제농도(MIC)를 판독하여 Clinical and Laboratory Standards Institute (CLSI) 기준에 따라 항생제 감 수성을 판정하였다. VITEK2 AST-N169 test (bioMérieux)는 제조사 프로토콜에 따라 실시하였다.
결과 및 고찰
균주 특성 및 배양 검사 결과
본 연구에 사용된 균주의 생화학적 동정 결과 해수 및 갯벌 에서 분리한 61균주 모두 생화학적으로 Vibrio vulnificus로 판 정되었다. 월별 검출 시기를 살펴보면, 해수 온도가 올라가기 시작하는 4월부터 발견되어 9월까지 해양 환경에서 검출되었 다(Table 1). 비브리오 패혈증균은 주로 해수 온도 18도 이상 에서 활발히 증식하는 것으로 알려져 있어 우리나라에서는 주 로 초여름부터 늦가을까지 발견되고 있으나 세계적으로는 기 후 변화 등의 원인으로 점점 그 분포 영역과 발견 시기가 확대 되고 있다(Baker-Austin and Oliver, 2018).
비브리오 패혈증균의 생물형은 균주의 생화학적 특성에 따
라 3가지로 분류된다. I형은 인체감염의 대부분을 차지하며, II 형은 뱀장어에 감염을 일으키는 것으로 알려져 있다. III형은 가 장 최근에 발견되었으며 I형과 II형의 특징을 교차하여 나타내 며 이스라엘에서만 발생이 보고되었다(Jones and Oliver, 2009).
본 연구에서는 61개의 균주 모두 인돌양성(Indole positive)인 생물형 I형으로 확인되었다.
배양검사 결과 용혈능은 61개 균주(100%)에서, 단백질분 해능은 59개 균주(96.7%)에서 확인되었다. 비브리오 패혈증 균은 hemolysin을 생성하여 숙주 세포를 파괴시켜 혈관 확장, 부종 등의 각종 염증 반응을 유발하고 적혈구를 파괴시키는 용혈 작용을 일으켜 체내 손상을 일으킨다(Jones and Oliver, 2009; Mahmoud and Chien, 2018). 또한 비브리오 패혈증균이 생성하는 단백질 분해효소는 숙주에 침투 시 단백질을 분해하 거나, 균주의 병원성 인자의 전처리 과정에 관여함으로서 균 주의 병원성에 영향을 미친다(Shinoda and Miyoshi, 2011). 이 로 보아 해안 환경에서 분리한 균주 중 대부분이 잠재적으로 병원성을 가지고 있음을 유추할 수 있다.
독성 유전자 검출 현황
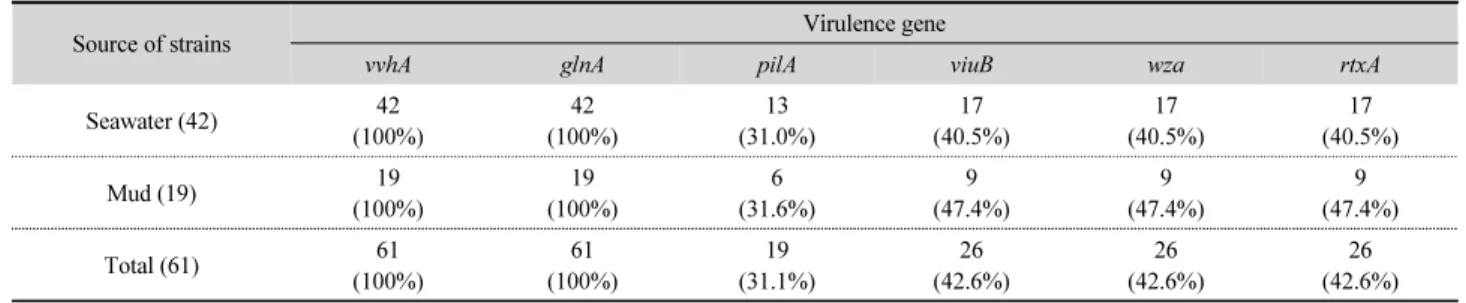
61개 균주에 대한 각 독성유전자의 검출결과, vvhA와 glnA는 61개 균주(100%)에서 검출되었고, pilA는 19개 균주(31.1%), viuB, wza, rtxA는 각각 26개 균주에서(42.6%) 검출되었다 (Table 3). 분리원에 따른 검출 현황을 살펴보면, pilA는 해수 유래 및 갯벌 분리 균주의 검출율이 비슷한 양상을 보였으나, viuB, wza 및 rtxA는 해수 분리 균주(40.5%)보다 갯벌 분리 균 주에서(47.4%) 높게 나타났다. 이는 viuB 유전자가 해수보다 갯벌에서 높은 검출율을 나타낸다고 보고된 바(Oh et al., 2007) 와 비슷하였다. rtxA는 비브리오 패혈증균의 독성 발현에 관여 하는 주요 유전자로 알려져 있는데(Lee et al., 2008), 본 연구에 서 확인된 환경 분리 균주에서의 검출률은(42.6%) 2008년 보 고된 임상 분리 균주(76.0%)와 해수 분리 균주(76.5%) (Hong
Table 2. Primer list for conventional-PCR
Target Primer Amplicon Size (bp) Annealing temperature (°C)
vvhA TTCCAACTTCAAACCGAACTATGA
205 60
ATTCCAGTCGATGCGAATACGTTG
viuB GGTTGGGCACTAAAGGCAGATATA
316 63
TCGCTTTCTCCGGGGCGG
pilA TGGCTGCTGTTGCTATTC
217 57
GGTCCACCACTAGTACCAAC
wza ATTCCGTGACCGATTGAGCGT
880 55
GCAGTAGAAGATACACCTAGG
et al., 2018)보다 다소 낮은 검출률을 보였다.
61개의 균주 중 55개 균주에서 최소 3개 이상의 유전자가 검출되었다(Table 4). 6개 독성 유전자가 모두 검출된 균주는 3개(4.9%)였는데, 이는 2016년 평택 갯벌, 2016년 안산 해수, 2018년 김포 해수에서 분리된 균주로 확인되었다. 비브리오 패혈증균은 인체 감염 및 병원성 발현에 관여하는 다양한 독 성인자를 가지는데, 균주 표면의 지질다당질, 편모(flagella), 섬모(pili) (Horseman and Surani, 2011), 철 획득 기전에 관여 하는 외독소(exotoxin), 용혈소(hemolysin) 등이 알려져 있다 (Gulig et al., 2005; Jones and Oliver, 2009). 이러한 결과를 종 합하여 볼 때, 경기 서해안 지역의 해양 환경에서 분리된 균주 모두 2개 이상의 독성유전자를 보유하고 있어 잠재적인 병원 성을 가지고 있음을 유추할 수 있으며 이에 따라 경기 서해안 지역에서 해수를 통한 창상감염 및 어패류 섭취를 통한 감염 에 주의해야 할 것으로 판단된다.
항생제 감수성 분석
61개 균주에 대하여 항생제 감수성 실험을 한 결과, Amoxi- cillin (아목시실린, ≤ 8)을 포함한 19개 항생제에 대하여 모든 균주가 감수성을 나타내었다(Table 5). 그 뒤를 이어 Chloram- phenicol (클로람페니콜, ≤ 8, 98.4%), Gentamicin (겐타마이 신, ≤ 4, 93.4%), Erythromycin (에리스로마이신, ≤ 0.5, 44.3%),
Tylosin tartrate (틸로신 타르타르산염, ≤ 1, 8.2%) 순으로 감 수성을 나타내었다. Penicillin (페니실린, ≥ 0.06)과 Cefazolin (세파졸린, ≥ 8)에는 모든 균주가 내성을 나타내었고 Cefoxitin (세폭시틴, ≥ 32, 98.4%), 틸로신 타르타르산염(≥ 4, 91.8%), Clindamycin (클린다마이신, ≥ 4, 86.9%) 순으로 나타났다.
가장 높은 중등도 내성을 보인 항생제는 에리스로마이신(0.5
< MIC < 8, 55.7%)이었으며, 클린다마이신(0.5 < MIC < 4, 13.1%), 겐타마이신(4 < MIC < 16, 6.6%), 세폭시틴(8 < MIC
< 32, 1.6%) 및 클로람페니콜(8 < MIC < 32, 1.6%) 순으로 중 등도 내성이 확인되었다.
비브리오 패혈증의 치료에는 흔히 3세대 세팔로스포린, 테 트라사이클린계, 플루오르퀴놀론계 및 클로람페니콜 약제가 사용되는데(Chuang et al., 1997; Han et al., 2007), 본 실험에 서는 이들 약제에 대한 내성은 거의 확인되지 않아 환경 균주 에 의한 직접감염 시 현 치료법이 유용할 것으로 판단되나, 1개 균주에서 클로람페니콜에대한 중등도 내성이 확인되었다. 항 생제에 대해 중등도 내성을 나타내는 경우 치료에 적용시 약 제의 효과가 충분히 나타나지 않을 수 있어 고용량의 투여가 이루어지게 되고(Rodloff et al., 2008), 이는 항생제 내성 균주 의 출현을 야기시킬 수 있다고 알려져 있으므로(Hamdani et al., 2020) 주의가 필요하다.
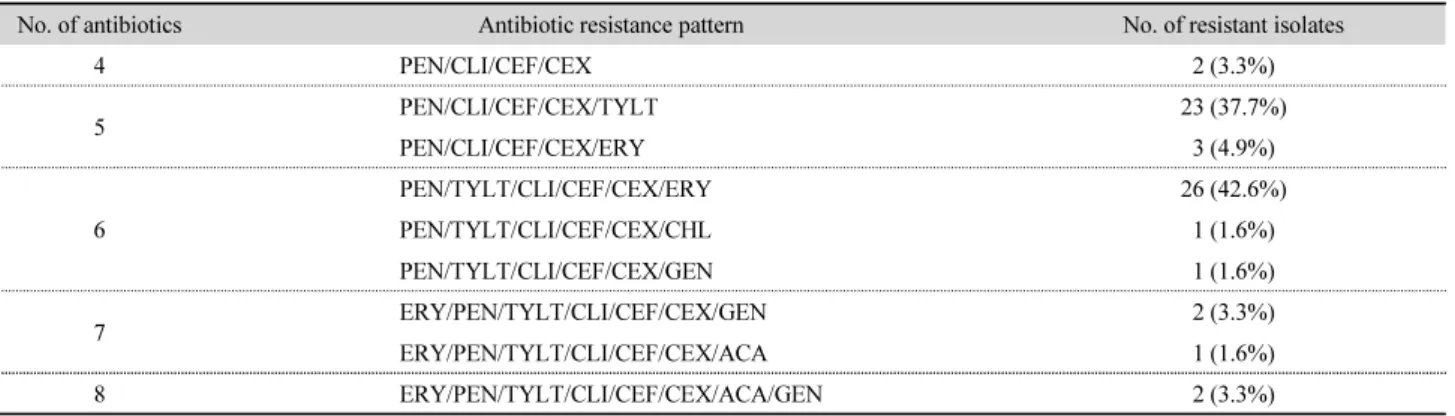
내성 또는 중등도 내성을 보이는 항생제 개수에 따라 균주 를 분류한 결과 총 9개의 패턴이 확인되었다(Table 6). 가장 많 이 확인된 패턴은 페니실린, 틸로신 타르타르산염, 클린다마 이신, 세파졸린, 세폭시틴 및 에리스로마이신이 포함된 그룹 이었으며(42.6%), 8개 항생제에 내성이나 중등도 내성을 보이 는 균주는 2개(3.3%) 있었는데, 2018년 평택 갯벌과 2020년 화성 해수에서 분리된 균주였다. 본 연구에서는 해양 환경에 서 분리한 비브리오 패혈증균에서 다양한 패턴의 항생제 내성 을 볼 수 있었는데, 이렇듯 항생제 내성에 관여하는 유전자는 환경에 흔하게 존재하며(Zhang et al., 2009) 농·축·수산업 및 의료계의 항생제 오남용은 환경에서 새로운 항생제 내성 유전 자가 나타나는데 기여하고, 인체 병원균은 다시 환경균으로부
Table 3. Detection of genes vvhA, glnA, pilA, viuB, wza and rtxA in tested strains
Source of strains Virulence gene
vvhA glnA pilA viuB wza rtxA
Seawater (42) 42
(100%) 42
(100%) 13
(31.0%) 17
(40.5%) 17
(40.5%) 17
(40.5%)
Mud (19) 19
(100%) 19
(100%) 6
(31.6%) 9
(47.4%) 9
(47.4%) 9
(47.4%)
Total (61) 61
(100%) 61
(100%) 19
(31.1%) 26
(42.6%) 26
(42.6%) 26
(42.6%)
Table 4. Number of isolates by number of virulence genes
No. of virulence genes No. of isolates
2 6
(9.8%)
3 25
(41.0%)
4 21
(34.4%)
5 6
(9.8%)
6 3
(4.9%)
Table 5. Antimicrobial susceptibility of Vibrio vulnificus isolates
Class of antibiotics Antibiotics MIC criteria (μg/ml) No. of isolates
R S R I S
Penicillins
Amoxicillin ≥ 32 ≤ 8 0 0 61 (100%)
Penicillin ≥ 0.06 ≤ 0.03 61 (100%) 0 0
Ampicillin ≥ 32 ≤ 8 0 0 61 (100%)
Cephalosporins
Cefazolin ≥ 8 ≤ 2 61 (100%) 0 0
Cefoxitin ≥ 32 ≤ 8 60 (98.4%) 1 (1.6%) 0
Ceftiofur ≥ 8 ≤ 2 0 0 61 (100%)
Carbapanems Imipenem ≥ 4 ≤ 1 0 0 61 (100%)
Aminoglycosides
Amikacin ≥ 64 ≤ 16 0 0 61 (100%)
Gentamicin ≥ 16 ≤ 4 0 4 (6.6%) 57 (93.4%)
Streptomycin ≥ 64 ≤ 32 0 0 61 (100%)
Neomycin ≥ 16 ≤ 8 0 0 61 (100%)
Spectinomycin ≥ 64 ≤ 32 0 0 61 (100%)
Tetracyclins Tetracycline ≥ 16 ≤ 4 0 0 61 (100%)
Oxytetacycline ≥ 16 ≤ 4 0 0 61 (100%)
Macrolides Erythromycin ≥ 8 ≤0.5 0 34 (55.7%) 27 (44.3%)
Tylosin tartrate ≥ 4 ≤ 1 56 (91.8%) 0 5 (8.2%)
Chloramphenicol Chloramphenicol ≥ 32 ≤ 8 0 1 (1.6%) 60 (98.4%)
Florfenicol ≥ 8 ≤ 2 0 0 61 (100%)
Lincosamides Clindamycin ≥ 4 ≤ 0.5 53 (86.9%) 8 (13.1%) 0
Fluorquinolones Enrofloxacin ≥ 2 ≤ 0.5 0 0 61 (100%)
Ciprofloxacin ≥ 4 ≤ 1 0 0 61 (100%)
Sulfonamides
Sulfadimethoxine ≥ 512 ≤ 256 0 0 61 (100%)
Sulfathiazole ≥ 128 ≤ 16 0 0 61 (100%)
Trimethoprim/sulfamethoxazole ≥ 4/76 ≤ 2/38 0 0 61 (100%)
Others
Novobiocin ≥ 8 ≤ 4 0 0 61 (100%)
Amoxicillin/Clavulanic acid ≥ 32/16 ≤ 8/4 0 0 61 (100%)
Ampicillin/Sulbactam ≥ 32/16 ≤ 8/4 0 0 61 (100%)
R, resistance; I, intermediate resistance; S, sensitive.
Table 6. Antibiotic resistance pattern of Vibrio vulnificus by number of resistant or intermediate-resistant antibiotics
No. of antibiotics Antibiotic resistance pattern No. of resistant isolates
4 PEN/CLI/CEF/CEX 2 (3.3%)
5 PEN/CLI/CEF/CEX/TYLT 23 (37.7%)
PEN/CLI/CEF/CEX/ERY 3 (4.9%)
6
PEN/TYLT/CLI/CEF/CEX/ERY 26 (42.6%)
PEN/TYLT/CLI/CEF/CEX/CHL 1 (1.6%)
PEN/TYLT/CLI/CEF/CEX/GEN 1 (1.6%)
7 ERY/PEN/TYLT/CLI/CEF/CEX/GEN 2 (3.3%)
ERY/PEN/TYLT/CLI/CEF/CEX/ACA 1 (1.6%)
8 ERY/PEN/TYLT/CLI/CEF/CEX/ACA/GEN 2 (3.3%)
PEN, Penicillin; CLI, Clindamycin; CEF, Cefazolin; CEX, Cefoxitin; TYLT, Tylosin tartrate; ERY, Erythromycin; CHL, Chloramphenicol; GEN, Gentamicin; ACA, Amoxicillin/Clavulanic acid
터 항생제 내성을 획득하게 된다(Finley et al., 2013). 비브리 오 패혈증은 해양 환경에 상재하는 비브리오 패혈증균이 인체 에 기회감염하는 것으로 알려져 있으며 병증이 빠르게 진행되 는 특성상 감염치료시 항생제 선택이 중요하다. 본 연구를 통 해 해수 및 갯벌에서 분리한 비브리오 패혈증균이 잠재적으로 항생제 내성을 나타낼 수 있음을 확인하였으므로, 감염 치료 시 항생제 내성까지 고려하여 적절한 항생제를 선택하는 것에 주의가 필요할 것이다.
적 요
비브리오 패혈증균(Vibrio vulnificus)은 인체에 기회감염 되어 급성 패혈증을 일으키는 해양 환경 상재균이다. 본 연구 에서는 2016년부터 2020년까지 경기 서해안 해수 및 갯벌에 서 분리한 61균주를 대상으로 하여 독성 및 항생제 감수성 등 의 균주 특성을 조사하였다. 대부분의 균주가 단백질 분해능 과 용혈능을 나타냈으며 6개의 독성유전자 검출 결과, vvhA, 100%; glnA, 100%; pilA, 31.1%; viuB, 42.6%; wza, 42.6%;
rtxA, 42.6%의 검출률을 보였다. 총 27종의 항생제에 대하여 감수성을 분석한 결과, Penicillin 등 5종의 항생제에 대하여 86.9~100%의 내성을 보였으며, Cefoxitin 등 5종의 항생제에 대하여 1.6~55.7%의 중등도내성을 보였다. 본 연구를 통해, 서해안 해양 환경에 상재하는 비브리오 패혈증균이 인체 감염 에 충분한 독성을 가지고 있으며 치료제로 사용되는 클로람페 니콜 약제에 중등도 내성을 가지는 균주가 존재함을 확인하였 다. 이는 해양 환경으로부터 유래되는 비브리오 패혈증균의 감염 예방 및 치료 방향 제시에 도움이 될 것으로 생각된다.
감사의 말
본 연구는 경기도보건환경연구원의 연구 사업으로 진행되 었습니다.
Conflict of Interest
The authors have no conflict of interest to report.
References