J. S. Han ( )
경북대학교 생태환경대학 생태환경전공, 경북대학교 대학원 생태과학과
(Department of Ecological Environment, Kyungpook National University, Sangju 742-711, Korea, Department of Ecological Science, Graduate School, Kyungpook National University, Sangju 742-711, Korea)
e-mail: [email protected] M. H. Jeong
농촌진흥청 국립농업과학원 농산물안전성부
(Department of Agro-Food Safety, National Academy of Agricultural Science, Rural Development Administration, Suwon 441-707, Korea)
I. K. Kang ・ C. K. Kim ・ C. Choi 경북대학교 농업생명과학대학 원예과학과
(Department of Horticultural Science, Kyungpook National University, Daegu 702-701, Korea)
K. I. Park
영남대학교 자연자원대학 원예생명과학과
(Department of Horticulture and Life Science, Yeungnam University, Gyeongsan 712-749, Korea)
애기장대 H+-pyrophosphatase 발현 형질전환 배추의 건조스트레스에 대한 생리적 반응
정미혜 ・ 강인규 ・ 김창길 ・ 박경일 ・ 최 철 ・ 한증술
Physiological responses to drought stress of transgenic Chinese cabbage expressing Arabidopsis H+-pyrophosphatase
Mihye Jeong ・ In-Kyu Kang ・ Chang Kil Kim ・ Cheol Choi ・ Kyung Il Park ・ Jeung-Sul Han
Received: 25 July 2013 / Accepted: 28 August 2013
ⓒKorean Society for Plant Biotechnology
Abstract Plant tolerance to drought is a beneficial trait for stabilizing crop productivity under water deficits. Here we report that genetically engineered Chinese cabbage expressing Arabidopsis H+-pyrophosphatase (AVP1) shows enhanced physiological parameters related to drought tolerance. In comparison with wild type plants under soil water deficit stress created by cessation of irrigation, soil water potential in pot with AVP1-expressing plants was more rapidly decreased that might lead to increased relative water content in leaves, while both genotypes had indistinguishable wilting phenotypes. Transgenic plants subjected to drought treatment also exhibited higher photosystem II quantum yield in addition to lower electrolyte leakage and H2O2-3,3’-diamino-
benzidine content when compared to wild type plants.
Keywords Chinese cabbage, H+-pyrophosphatase, Genetic transformation, Drought tolerance
서 론
작물에 영향을 미치는 다양한 비생물 스트레스 중 건조 스트레스와 염스트레스는 세계 경작지의 10% 이상에 영 향을 미쳐 생산성을 심각하게 훼손하고 있고 지구온난화 에 따른 기후변화와 더불어 그 피해는 지속적으로 증가 될 것으로 예측된다(Aiguo 2011; Baltels and Sunkar 2005).
우리나라의 연평균 강우량은 대략 1,200 mm로 세계 연 평균 강우량인 약 970 mm에 비해 적지는 않지만 60% 정 도의 강우가 특징적으로 여름철에 집중되어 있어 농경지 는 침수와 한발의 피해를 동시에 받고 있다. 특히 여름철 호냉성 채소를 대규모 재배하는 고령지는 배수는 양호한 반면 대부분 지역이 관개시설의 설치가 어려워 관수를 자연강우에 의존하고 있기 때문에 한발의 피해에 고스란 히 노출되어 있다고 할 수 있다. 실제로 우리나라 주요 채소 중 하나인 고령지 결구배추의 가격 등락폭이 해에 따라 매우 심한데, 그 직간접적 원인 중 하나로 여름철 한발이 지목된다. 따라서 내건성을 갖춘 품종의 개발과 재배는 배추를 비롯한 다양한 작물의 안정생산에 기여할 수 있는 수단 중 하나가 될 수 있을 것이다.
스트레스 종류별로 식물체의 방어 전략에 차이가 있기 는 하지만 건조스트레스를 포함한 비생물 스트레스를 받 은 식물체는 분자수준에서 서로 연관성을 가지며 복잡하 게 네트워크화된 신호전달, 전사조절, 특이 효소계 발현 DOI:http://dx.doi.org/10.5010/JPB.2013.40.3.156
Research Article
을 통한 무독화, 삼투조절 및 이온수송을 통한 각종 항상 성 회복 체계 등을 작동시킴으로써 스트레스에 저항한다 (Krasensky and Jonak 2012; Liu et al. 2000; Wang et al. 2003;
Zhu 2003). 그 중 애기장대에서 동정된 액포막 소재 H+ pump의 하나인 H+-pyrophosphatase (AVP1)는 막을 사이에 두고 H+ 구배를 형성하여 다른 이온수송체에게 전기화학 적 동력을 제공함으로써 간접적으로 이온항상성 회복을 담당하며 Pinformed 1 auxin efflux facilitator의 분포에도 영 향을 미치는 것으로 알려져 있다(Hirschi 2001; Li et al.
2005; Zhu 2003). 실제로 AVP1을 과발현하는 다양한 식물 종에서 내건성 획득, 내염성 획득 및 생물량 증가라는 표 현형이 동시에 나타난다(Gaxiola et al. 2001; Park et al.
2005; Pasapula et al. 2010; Yang et al. 2007).
우리는 지난 연구 보고(Park et al. 2012)에서 AVP1 유전 자를 배추 내혼계통에 형질전환하고 T2 고정세대를 육성 한 후 생리검정을 수행하여 배추에서도 AVP1 과발현이 생물량 증가와 염스트레스 내성 향상에 기여할 수 있음 을 검증하였다. 금번 연구에서는 AVP1 형질전환 배추 T2
고정세대로부터 T3 세대로 진전한 후 AVP1 과발현의 또 다른 주요 영향 중 하나인 내건성에 초점을 맞추어 다양 한 생리검정을 수행함으로써 배추에서 역시 AVP1 과발 현이 내건성을 향상시킬 수 있음을 확인하였다.
재료 및 방법
AVP1 형질전환 배추 계통
LB::T35S::AVP1::2×P35S::PNOS::Bar::TNOS::RB T-DNA 로 배추 (Brassica rapa L.) 내혼계통 ‘KSSD2810’을 형질전환 한 후 세대진전과 선발을 거쳐 육성한 목표유전자 고정 T2 독 립 계통 중 비형질전환체(wild type: WT)에 비해 생물량 이 많고 점진적으로 강화시킨 염스트레스 조건에서도 실 광합성 효율이 높으면서 체내에 활성산소족의 하나인 H2O2 존재량이 적은 것으로 확인된 ‘CCAVP05’와 ‘CCAVP06’
계통의 T2세대 종자를 본 연구의 시작 식물재료로 사용 하였다(Park et al. 2012).
형질전환 T3세대 진전
형질전환 배추 ‘CCAVP05’와 ‘CCAVP06’의 T2 세대 종자 를 각각 파종하여 육묘한 본잎 5~6매 시점의 유식물체를 냉장진열장(ADVANZA: Carrrier, Korea)에 입상하여 8±3°C, 16 h (약 20 μmol․m-2․s-1) 조건에서 간헐적으로 관수하며 약 4 주간 춘화처리 시켰다. 춘화처리가 완료된 식물체는 상 온으로 이동하여 관리하다가 내엽 윗면의 왁스층 발달이 관찰되는 시점에 내경 30 cm 화분으로 이식하여 비배관
리하며 추대를 유도하였다. 꽃가루 오염을 방지하기 위 해 개화기 전에 봉지 씌우기를 하였으며 만개기에 봉지 를 벗겨 인공 자가 뇌수분하였다. 각 계통 당 미숙모본으 로 5개체를 활용하였으며 교배 후 평균 6주경에 씨꼬투 리를 채취하였고 채취한 씨꼬투리는 음건한 후 탈종하여 각 계통의 T3 세대 종자를 확보하였다(Fig. 1A).
생리검정용 유식물체 양성
AVP1 전사체 탐지, 제초제 저항성 검정, 관수중단에 따른 토양수분포텐셜 측정, 광계II 양자수율(фPSII) 측정, 상대 적 수분함량 측정, 전해질누출 측정 및 3,3’-diaminobenzidine (DAB: Sigma-Aldrich, USA)염색 등을 위해 수확한 T3 세대 각 계통의 종자를 최아하여 유근의 생장이 유사한 실생 만을 고른 후 상토를 담은 32공 plug tray에 이식하고 배 양실(25±1°C, 16 h 일장, 약 20 μmol․m-2․s-1, 상대습도 45±5%)에서 주기적으로 저면관수하며 본잎 3~4매기까지 유식물체를 양성하였다(Park et al. 2012).
AVP1 전사체 탐지와 제초제 저항성 검정
무작위로 선택한 각 계통의 4개 유식물체로부터 어린 잎 을 채취하여 Han (2010)의 방법에 따라 RNeasy Plant Mini Kit (Qiagen GmBH, German)을 사용하여 총 RNA를 추출한 후 오염되어 있을 수 있는 DNA를 제거하기 위하여 DNase I (Invitrogen, USA)으로 2회 처리하였다. RNA는 전기영동 을 통해 대략적 농도와 순도를 파악하는 한편 분광광도 계(NanoDrop 2000: Thermo Fisher Scientific Inc., USA)로 실 측한 후 시료당 동일한 농도가 되도록 희석하였다. 희석한 등량의 RNA를 주형으로 Super Script III First-Strand Synthesis System (Invitrogen, USA)을 사용하여 cDNA를 역전사하였 으며, 합성된 cDNA는 전기영동을 통해 확인하였다. cDNA 와 AVP1 유전자 특이적 forward (5’-gtcggcgctgaccttgtcggtaaa-3’) 및 reverse (5’-ccaatgataactttaggggtcaaa-3’) primer를 각각 PerfectShot Ex Tag Kit (Takara Bio Inc., Japan)에 첨가하여 중합효소연 쇄반응(PCR)을 수행하여 AVP1 전사체 유래 cDNA를 증폭 하였다. 한편, 0.3%로 희석한 제초제 BastaTM (Bayer Crop Science, Korea)를 유식물체 전체에 살포하고 10일 후 그 피해를 관찰하였다.
관수중단과 토양수분포텐셜 추적
배양실에서 동일한 방법으로 생장시킨 각 계통의 유묘가 재식된 plug tray에 포장용수량까지 최종 관수한 후 관수 를 중단하고 토양수분장력계(T5×/INFIELD7: USM GmbH, Germany)로 깊이 4 cm 위치의 토양수분포텐셜을 관수중 단 6일 후까지 계통당 5개체에 대하여 매일 측정하였다.
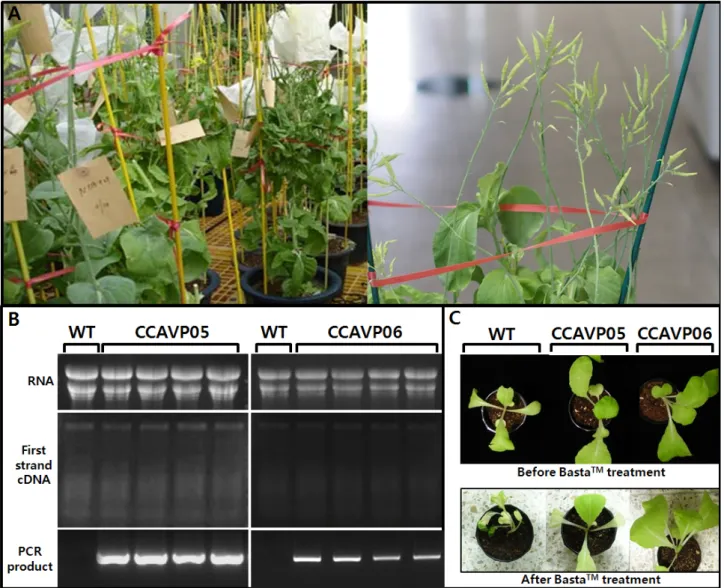
Fig. 1 Advancement forward T3 generation, transcript detection, and herbicide bioassay in transgenic Chinese cabbage expressing Arabidopsis H+-pyrophosphatase (AVP1). (A) Flowering mother plants at T2 generation in a greenhouse (left) and siliques normally set by artificial self-pollination (right), (B) Detection of AVP1 transcript by reverse transcriptional polymerase chain reation using T3 plants, (C) Herbicide bioassay for the selectable marker gene Bar
엽록소형광소멸분석
관수중단 후 Park 등(2012)의 방법에 따라 Pulse amplitude modulation fluorometer (Mini-PAM: Heinz Walz GmbH, Germany) 를 사용하여 각 식물체의 제3엽을 대상으로 광계II 형광 (Chl-F)을 측정하였다. фPSII는 (Fm’-Ft)/Fm’식으로 구하였 는데 Fm은 암적응된 상태에서의 최대 Chl-F, Fm’은 명적 응된 상태에서의 최대 Chl-F, Ft는 명적응 후 포화광 pulse 조사 직전 Chl-F 값을 의미한다(Genty et al. 1989).
상대적수분함량 측정
관수중단 후 6일차에 계통당 5개 식물체로부터 각각 제3 엽을 채취하여 즉시 생체중(FW)을 측정한 후 증류수에 띄워 냉장고(4°C)에서 15시간 재흡수 시켰다. 잎 표면의
수분을 화장지를 이용하여 흡습 제거한 후 재흡수 생체 중(RFW)을 측정하였으며 70°C에서 15시간 처리하여 건조 중(DW)을 측정하였다. 상대적 수분함량(RWC)은 (FW-DW)/
(RFW-DW) 식으로 구하였다.
전해질누출 측정
관수중단 후 8일차에 각 계통의 4개 식물체로부터 제3엽 을 채취하여 탈이온수로 부드럽게 세척한 후 각 잎당 직 경 7 mm의 구멍을 2개씩 뚫어 검정용 시료를 채취하였 다. 15 mL 탈이온수를 담은 시험관에 시료를 넣고 잎 절편 표면의 공기방울을 제거하기 위하여 펌프(DOA-P704- AC: GAST Manufacturing Inc., USA)를 사용하여 6 psi에서 5분 간격으로 3회 감압하였다. 시료가 담긴 시험관을 비 스듬히 세워 300 rpm으로 2시간 진탕한 후 전기전도도계
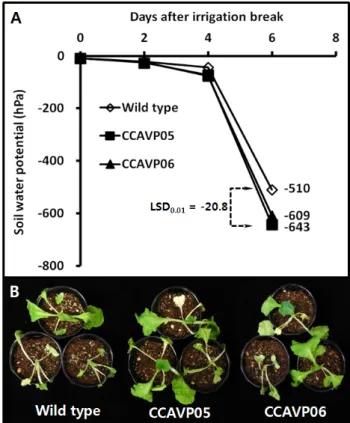
Fig. 2 Wilting phenotypes of AVP1-expressing Chinese cabbage plants under gradually intensified drought stress by irrigation break. (A) Changes of soil water potential after irrigation break, (B) Morphologies of each genotype on eight days after irrigation break. Mean value for least significant difference at the 1% level (LSD0.01; n=5) was directly indicated in frame A
(WalkLAB, Trans Instruments Ltd., Singapore)로 전기전도도 를 측정하였다. 시험관을 고압증기멸균기에 넣어 121°C 에서 20분간 끓인 후 용액의 전기전도도를 다시 측정하 였다. 전해질누출(%)은 시료를 끓이기 직전 측정한 전기 전도도를 끓인 후 측정한 전기전도도로 나눈 후 100을 곱하여 계산하였다(Shou et al. 2004).
DAB 염색
DAB 염색 처리를 통해 활성산소족의 하나인 H2O2를 탐 지하였다(Wu et al. 2012). Park 등(2012)의 방법을 일부 수 정하였는데, 관수중단 6일차에 잎을 채취하여 25±1°C, 암 상태로 0.1% DAB 용액(pH 3.8)에 12시간 침지하였다. 처 리 후 70°C, 94% 에탄올로 4회 탈색시켜 H2O2-DAB 산물 의 발색을 관찰하였다.
결과 및 고찰
형질전환 T3 계통에서 AVP1 전사체와 제초제 저항성 발현
애기장대 AVP1 유전자로 형질전환된 배추 내혼계통
‘KSSD2810’의 T2 세대 ‘CCAVP05’와 ‘CCAVP06’ 계통 각 식물체는 인공 자가 뇌수분에 의해 정상적으로 씨꼬투리 를 착생하였으며 T3 세대의 종자를 생산하였다(Fig. 1A).
각 T3 계통의 유식물체로부터 추출한 총 RNA로부터 역 전사를 통해 합성한 cDNA를 주형으로 PCR을 수행하였 는데, 무작위로 선택된 모든 식물체에서 T0 세대(Park et al.
2012)에서와 마찬가지로 전사체가 탐지되었고 ‘CCAVP05’
계통의 발현량이 ‘CCAVP06’ 계통에 비해 많았으며 각 계통의 개체간에는 균일한 정도의 전사량을 나타내었다 (Fig. 1B). 한편, 기내 형질전환에서 선발표지유전자로 사 용한 Bar의 발현을 검정하기 위하여 0.3% BastaTM 용액을 유식물체에 살포한 후 관찰하였는데, WT는 피해를 받아 고사한 반면 ‘CCAVP05’와 ‘CCAVP06’ 계통 모두는 정상 적 생육을 진행하였다(Fig. 1C). 따라서 AVP1 형질전환 배 추 두계통은 T3 세대에서도 다양한 원인에 의해 일어날 수 있는 transgene silencing (Dietz-Pfeilstetter 2010; Hammond et al. 2001) 없이 정상적으로 T-DNA를 유전 받았으며 최소 한 Bar 유전자는 정상적으로 작동한다고 판단할 수 있다.
관수중단 후 토양수분 변화와 형질전환체의 표현형
관수중단 후 유식물체가 재식된 토양의 수분포텐셜은 관 수중단 4일경을 기점으로 가파르게 감소하였는데, 관수 중단 6일차에 WT 재식 토양은 -510 hPa인 반면 형질전환 각 계통을 재식한 토양은 각각 -609 hPa과 -643 hPa로 형
질전환체를 재식한 토양의 수분포텐셜이 뚜렷하게 낮았 다(Fig. 2A). 비록 토양수분포텐셜이 형질전환체 재식 토 양에서 훨씬 낮기는 하였지만 관수중단 8일 후 가시적 위조의 정도에 있어서는 유전자형간에 차이가 없었다(Fig.
2B). 우리는 T2 세대를 대상으로 유식물체를 정상관수 조 건에서 생육시켰을 때 형질전환체 지상부의 생체중과 건 물중 모두가 WT에 비해 2배 이상 높다고 보고한 바 있다 (Park et al. 2012). 이러한 결과는 배추에서 AVP1 발현이 생물량 증가라는 표현형으로 나타나기 위하여 주어진 기 간에 생육에 필요한 수분을 토양으로부터 더 빠르게 흡 수하였기 때문으로 해석할 수 있을 것이며, 비록 더 낮은 토양수분포텐셜에서도 형질전환체는 WT과 비교하여 위 조의 정도에 차이를 나타내지 않는다는 점으로 미루어 볼 때, AVP1 형질전환체가 WT에 비해 건조스트레스에 더 둔감하다고 판단 할 수 있을 것이다.
건조스트레스 환경에서 형질전환체의 광합성효율 변화
많은 식물 종은 건조스트레스 조건에서 광합성 기관의 활동능력이 감소하거나 absisic acid 합성에 따라 기공이 폐쇄되고 세포내 CO2 농도가 감소하여 광합성이 저해되
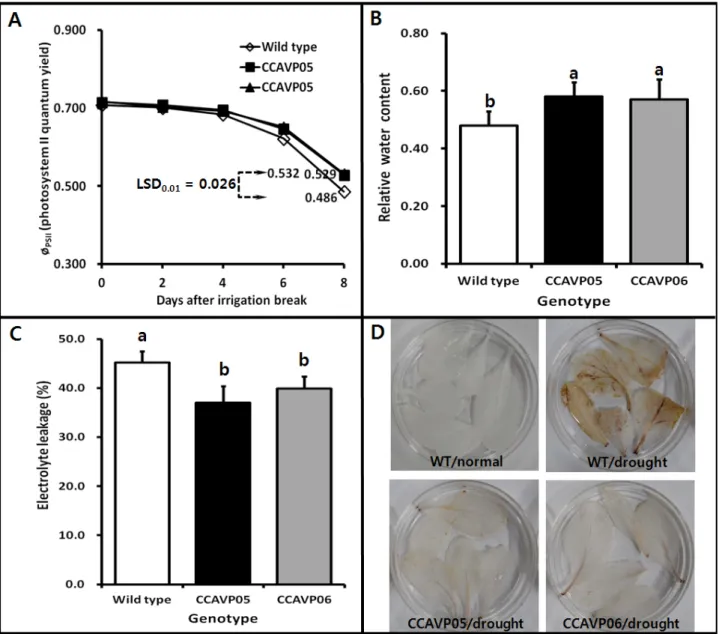
Fig. 3 Differences between AVP1-expressing and wild type Chinese cabbage plants in physiological parameters related to drought stress. (A) Changes of photosystem II quantum yield after irrigation break, (B) Relative water contents of leaves after 6 days of irrigation break, (C) Electrolyte leakage of leaf segments in each genotype after 8 days of irrigation break, (D) Detection of H2O2-3,3’-diaminobenzidine reaction products after 6 days of irrigation break. Mean value for least significant difference at the 1%
level (LSD0.01; n=5) was directly indicated in frame A. Mean values followed by different letter in frames B and C are significantly different at the 5% level (n=5 and n=8, respectively) (Duncan’s multiple range test), and error bars denote the standard deviation values
는 것으로 알려져 있다(Bhargava and Sawant 2013; Thapa et al. 2011). 강화되는 건조스트레스 조건에서 AVP1 형질전 환 배추의 실제 광합성효율을 판단할 수 있는 척도인 ф
PSII를 엽록소형광소멸분석을 통해 추적하였다. 관수중단 후 토양수분포텐셜이 급하게 감소하는 4일차(Fig. 2A)부 터 фPSII 역시 연동하여 WT과 형질전환체 모두 급격히 감 소하였다(Fig. 3A). 그러나 형질전환체 фPSII가 wild type에 비해 완만하게 감소하였는데, 관수중단 8일차에 형질전 환 계통이 각각 0.532와 0.529인 반면 WT은 0.486으로 훨 씬 낮았다. 즉, 강한 건조스트레스 조건에서도 형질전환
체는 wild type에 비해서는 상당히 높은 광합성효율을 유 지한다고 할 수 있다.
건조스트레스 환경에서 형질전환체의 상대적 수분함량
건조스트레스를 받은 특정 시점에 WT과 AVP1 형질전환 체의 잎이 유사하게 낮은 수분함량을 가진다면 위에서 관찰한 양 유전자형간 광합성효율의 차이를 설명하기 어 려울 수도 있다(Thapa et al. 2011). 따라서 우리는 관수중 단 처리 6일 후 양 유전자형의 RWC를 측정하였다. WT
의 RWC는 0.48±0.05 (평균±표준편차)로 ‘CCAVP05’ 계통 의 0.58±0.05와 ‘CCAVP06’ 계통의 0.57±0.07에 비해 각각 17.2%와 15.8% 낮았다(Fig. 3B). 이러한 결과는 형질전환 체가 재식된 토양의 수분포텐셜이 WT가 재식된 토양보 다 더 빠르게 감소한 결과(Fig. 2A)를 반영한 것이거나 건 조스트레스 조건에서 형질전환 식물체의 증산량이 상대 적으로 적기 때문이라고 추정할 수 있다. 결과론적으로 형질전환 식물체는 건조스트레스 환경에서 수분 보유 능 력이 우수하며, 따라서 형질전환체가 상대적으로 높은 광합성효율을 가진다는 점을 부분적으로나마 설명할 수 있는 증거가 될 수 있을 것이다. Gaxiola 등(2001)과 Park 등(2005)은 각각 AVP1 유전자를 과발현하는 애기장대와 토마토의 잎이 WT에 비해 건조스트레스 상황에서 높은 수 준의 RWC 또는 수분포텐셜을 가진다고 보고한 바 있다.
건조스트레스 환경에서 형질전환 식물체의 전해질 누출
건조스트레스를 포함하여 다양한 비생물 스트레스를 받은 식물체는 많은 경우 세포막이 피해를 받아 세포내 전해 질이 누출된다(Lu et al. 2013; Wang et al. 2003). 본 연구에 서는 건조스트레스를 심하게 받고 있는 관수중단 처리 8 일 후 AVP1 형질전환체와 WT의 잎 절편을 채취하여 전 해질누출을 측정하였다. 형질전환체의 전해질누출은 각 계통이 37.0±3.4%와 39.9±2.5%로 WT 45.2±2.3%에 비해 각 각 18.1%와 11.7% 적었다(Fig. 3C). 물론 각 형질전환 계 통의 전해질 누출도 절대적으로 꽤 높은 수준이기는 하 지만(Lu et al. 2013; Shou et al. 2004) WT에 비해서는 뚜렷 하게 낮아 형질전환체의 막투과성 유지능력이 상대적으 로 우수한 것으로 사료된다.
형질전환체에서 H2O2-DAB 반응산물 탐지
DAB 염색 처리를 통해 다양한 비생물 스트레스에 의해 발생할 수 있는 체내 활성산소족의 하나인 H2O2를 탐지 할 수 있다(Guan et al. 2012; Wu et al. 2012). 정상관수조건 에서 생육시킨 WT의 잎에서는 갈색반점으로 대별되는 H2O2-DAB 반응산물이 전혀 관찰되지 않았고, 관수중단 처리 6일 후 형질전환체 잎에서는 중륵의 통도조직만을 중심으로 연갈색의 반응산물이 관찰된 반면 WT의 경우 는 중륵부분 뿐 아니라 엽신 모두에서 짙은 갈색의 반응 산물이 침착되었다(Fig. 3D). 즉, AVP1 형질전환체는 건조 스트레스에 의해서도 활성산소족의 하나인 H2O2를 생산 하며 비록 AVP1 발현이 직접적으로 H2O2 생산을 억제하 였는지, 아니면 H2O2의 분해를 촉매하는 catalase 등을 포 함한 다양한 활성산소족 소거 효소계 또는 비효소계의 활성화에 간접적으로 기여하였는지는 본 실험 결과로는 확정하기 어렵지만(Wang et al. 2003) 최소한 특정 시점에
AVP1 형질전환체의 체내에 WT에 비해서는 상대적으로 낮은 수준의 H2O2가 존재한다고 판단할 수는 있다.
지금까지 AVP1 유전자를 발현하는 배추 T2 계통을 세 대진전한 후 T3 세대 식물체가 건조스트레스를 받았을 때의 수분관계, 광합성효율, 세포막투과성 및 활성산소 족 발생 등의 측면에서 어떻게 생리적으로 반응을 하는 지 WT와 비교하여 검토하였다. 결론적으로 배추에서 AVP1 유전자의 발현은 내염성(Park et al. 2012) 뿐 아니라 내건성 향상에도 기여할 수 있음을 본 연구를 통해 확인 하였다. ‘CCAVP05’ 계통의 전사체 발현량이 ‘CCAVP06’
계통에 비해 많았음에도 불구하고 내건성 관련 생리지표 에서는 둘간에 뚜렷한 차이를 나타내지 않은 점은 좀 더 정밀한 검토가 필요하다고 판단된다. 또한 우리는 여러 번의 시도에도 불구하고 아직 건조스트레스 처리 후 재 관수를 통해 WT와 형질전환체가 각각 고사와 회복을 구 분지을 수 있는 임계 스트레스 강도 등의 실험조건은 조 성하지 못하였다. 그러나 Bhargava와 Sawant (2013)는 영 구위조점을 넘어서는 심한 건조스트레스를 받은 식물체 는 결국 고사하게 되지만 식물체가 회복가능한 범위 내 에서 건조스트레스를 받을 경우 내성을 가진 유전자형은 그렇지 않은 유전자형에 비해 상대적으로 스트레스 관련 생리지표가 양호하다고 언급한 바 있다.
적 요
한발에 대한 식물체의 내성은 수분이 부족한 환경에서도 안정적으로 작물의 생산성을 유지하기 위해 필요한 유용한 특성 중 하나이다. 우리는 애기장대의 H+-pyrophosphatase (AVP1)를 발현하도록 형질전환된 배추가 내건성과 관련 되어 있는 몇몇 생리적 척도에 있어 향상됨을 검증하였다.
관수중단 처리로 조성된 토양수분 결핍 조건에서 AVP1 발현 식물체는 비형질전환체와 비교하여 비록 외형적 위 조의 정도로는 그 차이를 구별할 수 없었지만 형질전환 체가 재식된 토양의 수분포텐셜이 비형질전환체 재식 토 양에 비해 더 빠르게 낮아졌다. 이는 형질전환체 잎의 상 대적 수분함량이 비형질전환체에 비해 더 높은 것과 연 관되어 있는 것으로 사료된다. 또한 건조스트레스 환경 에서 비형질전환체에 비해 형질전환체는 광계II 양자수 율이 높은 반면 전해질누출과 활성산소족의 하나인 H2O2
와 3,3’-diaminobenzidine의 반응산물이 적었다.
사 사
「이 논문은 2010년도 경북대학교 학술연구비에 의하여 연구되었음」
인용문헌
Aiguo D (2011) Drought under global warming: a review. WIREs Clim Change 2:45-65
Baltels D, Sunkar R (2005) Drought and salt tolerance in plants.
Crit Rev Plant Sci 24:23-58
Bhargava S, Sawant K (2013) Drought stress adaptation: metabolic adjustment and regulation of gene expression. Plant Breed 132:21-32
Dietz-Pfeilstetter A (2010) Stability of transgene expression as a challenge for genetic engineering. Plant Sci 179:164-167 Gaxiola RA, Li J, Undurraga S, Dang LM, Allen GJ, Alper SL,
Fink GR (2001) Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc Natl Acad Sci USA 98:11444-11449
Genty B, Briantais JM, Baker NR (1989) The relationship between quantum yield of photosynthetic electro transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 990:87-92
Guan Q, Takano T, Liu S (2012) Genetic transformation and analysis of rice OsAPx2 gene in Medicago sativa. PLoS ONE 7:1-8
Hammond SM, Caudy AA, Hannon GJ (2001) Post-transcriptional gene siliencing by double-stranded RNA. Nature Genetics 2:110-119
Han J-S (2010) Suppression of tobamovirus movement toward upper leaves in the tomato plant over-expression a maize calreticulin. J Plant Biotechnol 37:567-573
Hirschi K (2001) Vacuolar H+/Ca2+ transport: who’s directing the traffic? Trend Plant Sci 6:100-104
Krasensky J, Jonak C (2012) Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J Exp Bot 63:1593-1608
Li J, Yang H, Peer WA, Richter G, Blakeslee J, Bandyopadhyay A, Titapiwantakun B, Undurraga S, Khodakovskaya M, Richards EL, Krizek B, Murphy AS, Gilroy S, Gaxiola R (2005) Arabidopsis H+-PPase AVP1 regulates auxin-mediated organ development. Science 310:121-125
Liu J, Ishitani M, Halfter U, Kim CS, Zhu JK (2000) The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc Natl Acad Sci USA 97:3730-3734
Lu Y, Li Y, Zhang J, Xiao Y, Yue Y, Duan L, Zhang M, Li Z (2013) Overexpression of Arabidopsis molybdenum cofactor sulfurase gene confers drought tolerance in maize (Zea mays L.). PLoS ONE 8:1-12.
Park M, Won H-Y, Kim CK, Han J-S (2012) Increased biomass and enhanced tolerance to salt stress in Chinese cabbage overexpression Arabidopsis H+-PPase (AVP1). J Plant Biotechnol 39:253-260
Park S, Li J, Pittman JK, Berkowitz GA, Yang H, Undurraga S, Morris J, Hirschi KD, Gaxiola RA (2005) Up-regulation of a H+-pyrophosphatase (H+-PPase) as a strategy to engineer drought-resistant crop plants. Proc Natl Acad Sci USA 102:18830-18835
Pasapula V, Shen G, Kuppu S, Paez-Valencia J, Mendoza M, Hou P, Chen J, Qiu X, Zhu L, Zhang X, Auld D, Blumwald E, Zhang H, Gaxiola R, Payton P (2010) Expression of an Arabidopsis vacuolar H+-pyrophosphatase gene (AVP1) in cotton improves drought- and salt tolerance and increases fibre yield in the field conditions. Plant Biotech J 9:88-99 Shou H, Bordallo P, Fan J-B, Yeakley JM, Bibikova M, Sheen J,
Wang K (2004) Expression of an active tobacco mitogen- activated protein kinase kinase kinase enhances freezing tolerance in transgenic maize. Proc Natl Acad Sci USA 101:3298-3303
Thapa G, Dey M, Sahoo L, Panda SK (2011) An insight into the drought stress induced alterations in plants. Biol Plantarum 55:603-613
Wang W, Vinocur B, Altman A (2003) Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta 218:1-14
Wu Q, Lin J, Liu JZ, Wang X, Lim W, Oh M, Park J, Rajashekar CB, Whitham SA, Cheng NH, Hirschi KD, Park S (2012) Ectopic expression of Arabidopsis glutaredoxin AtGRX17 enhances thermotolerance in tomato. Plant Biotechnol J 10:945-955
Yang H, Knapp J, Koirala P, Rajagopal D, Peer WA, Silbart LK, Murphy A, Gaxiola RA (2007) Enhanced phosphorus nutrition in monocots and dicots over-expressing a phosphorus-responsive type I H+-pyrophosphatase. Plant Biotech J 5:735-745 Zhu JK (2003) Regulation of ion homeostasis under salt stress.
Curr Opin Plant Biol 6:441-445