M. Park
농촌진흥청 국립원예특작과학원 채소과
(Vegetable Division, National Institute of Horticultural & Herbal Science, Rural Development Administration, 540-41 Tap-dong, Suwon 441-440, Korea)
H.-Y. Won
농촌진흥청 국립원예특작과학원 채소과, 경북대학교 대학원 생태과학과
(Vegetable Division, National Institute of Horticultural & Herbal Science, Rural Development Administration, 540-41 Tap-dong, Suwon 441-440, Korea, Department of Ecological Science, Graduate School, Kyungpok National University, 2559 Gyeongsangdaero, Sangju 742-711, Korea)
C. K. Kim
경북대학교 농업생명과학대학 원예과학과
(Department of Horticultural Science, College of Agriculture &
Life Sciences, Kyungpook National University, 80 Daehakro, Bukgu, Daegu 702-701, Korea)
J.-S. Han ( )
경북대학교 생태환경대학 생태환경전공, 경북대학교 대학원 생태과학과
(Department of Ecological Environment, College of Ecology &
Environmental Science, Kyungpook National University, 2559 Gyeongsangdaero, Sangju 742-711, Korea, Department of Ecological Science, Graduate School, Kyungpok National University, 2559 Gyeongsangdaero, Sangju 742-711, Korea) e-mail: [email protected]
애기장대 H+-PPase(AVP1) 과발현 배추에서 바이오매스 증가와 내염성 향상
박미희 ・ 원희연 ・ 김창길 ・ 한증술
Increased biomass and enhanced tolerance to salt stress in Chinese cabbage overexpressing Arabidopsis H+-PPase (AVP1)
Mehea Park ・ Hee-Yeun Won ・ Chang Kil Kim ・ Jeung-Sul Han
Received: 1 November 2012 / Accepted: 16 November 2012
ⓒKorean Society for Plant Biotechnology
Abstract On the basis of the reported agriculturally valuable phenotypes resulted from ectopic overexpression of Arabidopsis vacuolar H+-PPase (AVP1), we generated the Chinese cabbage lines expressing AVP1 which then subjected to salt stress to determine the AVP1 expression if it consistently confers the capability for increasing biomass and enhancing tolerance to salinity in other species. Collectively, here we demonstrate that the transgenic young plants show more vigorous growth and higher tolerance to salt stress than wild-type ones. Increased biomass phenotype by AVP1
expression was supported by comparing fresh and dry weights of transgenic and wild type plants grown under normal condition, while higher salt tolerance trait was confirmed by tracing the kinetics of photosystem II quantum yield and DAB-staining under gradually intensified salt stress induced by MS salt or NaCl, followed by normal condition.
Keywords Chinese cabbage, AVP1, transformation, salt tolerance, photosystem II quantum yield, DAB staining
서 론
혐염성 작물(glycophytic crop)의 생산성을 저하시키는 원 인 중 하나인 고염스트레스(이하 염스트레스)는 세계 경 작지의 약 20%(관개농지의 약 50%)에 피해를 주고 있다 (Zhu 2001). 염스트레스는 주로 식물 세포의 삼투항상성과 이온항상성을 교란시키고 추가적으로 산화스트레스를 유 발하여 피해를 주기 때문에(Apse et al. 1999; Greenway and Munns 1980; Qiu et al. 2002; Wang et al. 2003; Zhu 2001) 식물체는 고염스트레스에 저항하여 정상적으로 생육하 기 위해서 세포내의 복잡하게 네트워크화된 abiotic stress 내성 획득 장치를 작동시켜야 한다. 이러한 네트워크를 단순하게 1) SOS경로(Salt Overly Sensitive pathway)에 존재 하는 일련의 이온수송체를 up-regulation 시켜 이온항상성 을 회복, 2) MAPK(Mitogen-Activated-Protein Kinase) 연쇄반 응 등을 유발하여 삼투조절물질을 축적함으로써 삼투항상성 을 복구, 3) 저온이나 건조스트레스 내성 획득 과정과 유사하 게 CBF/DREB(C-Repeat-Binding/Dehydration-Responsive Element- Binding) 등 전사조절인자를 작동시켜 RD29A와 같은 스트 DOI:http://dx.doi.org/10.5010/JPB.2012.39.4.253
Research Article
레스 단백질을 발현시킴으로써 산화 유발물질을 무독화 시키는 것 등으로 요약할 수 있다(Kasuga et al. 2004; Liu et al. 2000; Zhu 2001). 그 중 살아있는 식물 세포의 특징 중 하나라 할 수 있는 이온항상성은 다양한 이온수송체 에 의해 직간접적으로 조절되는데, 특히 염스트레스 하 에서는 세포질에 고농도 K+과 저농도 Na+의 유지가 필요 하고 이를 위해 원형질막 또는 세포소기관 막에 소재하 는 Na+/H+ antiporter 등 이온 특이적 수송체와 H+ pump가 중 요한 역할을 수행한다(Apse et al. 1999; Hirschi 2001; Liu and Zhu 1998; Sanders et al. 2002; Zhu 2003). H+ pump는 막을 사이에 두고 양측간 H+ 구배를 형성함으로써 다른 이온수송 체에게 전기화학적 동력을 제공하는 것으로 알려져 있고 식 물에서는 대표적으로 H+-ATPase와 H+-pyrophosphatase(H+- PPase)가 알려져 있다(Hirschi 2001; Zhu 2003).
애기장대(Arabidopsis thaliana) H+-PPase(AVP1)는 액포 막에 풍부하게 존재하고 pyrophosphate 의존적으로 H+을 수송한다는 기능과 그 유전자와 단백질의 분자적 구조가 동정(Sarafian et al. 1992; Zhen et al. 1997)된 이래로 세포 질의 pH 유지와 더불어 식물체 생육에 중요한 호르몬인 오옥신의 수송을 간접적으로 조절한다는 사실이 추가적 으로 밝혀졌다(Li et al. 2005). 최근에는 모델 식물종인 애 기장대 뿐만 아니라 토마토, 목화 및 벼에서 AVP1 과발 현이 내염성 또는 내건성을 향상시킬 수 있으며, 흥미롭 게도 식물체의 바이오매스와 인산효율까지 증가시킨다 는 것이 알려졌다(Gaxiola et al. 2001; Gonzalez et al. 2010;
Park et al. 2005b; Yang et al. 2007). 이러한 AVP1의 다면발 현은 언급한 바와 같이 AVP1 발현이 다양한 이온수송체 에 동력을 제공하여 활성화 시키고(Hirschi 2001; Zhu 2003), 한편으로는 세포질 pH 변화를 야기시켜 원형질막 에 P-type H+-ATPase와 PIN 1(PINformed 1 auxin efflux fa- cilitator)을 양적으로 증가시키며 연이어 오옥신의 화학삼 투적 수송을 향상시킬 수 있다는 보고(Li et al. 2005)와 무 관하지 않은 것으로 사료된다. 이상 살펴본 바와 같이 몇 몇 식물 종에서 AVP1 유전자의 과발현은 내염성과 내건 성 뿐만 아니라 바이오매스 증가라는 농업적으로 상당히 매력적인 표현형으로 나타난다.
배추(Brassica rapa L.)는 한국, 중국, 대만 및 일본 등에 서는 중요한 채소 중 하나이고 다양한 작형으로 재배되 며 통상 연작을 하게 된다. 대부분의 농가는 재배 토양의 성분 분석을 생략하고 관행적으로 매년 기비와 추비를 시용하기 때문에 배추를 포함한 다양한 작물의 재배 토 양에서 염류집적으로 인한 피해가 문제점으로 부각되고 있다. 따라서 본 연구에서는 내염성과 관련이 있는 AVP1 유전자를 배추에 형질전환시킨 후 염스트레스에 대한 생 리반응과 바이오매스 변화에 초점을 맞추어 유식물체의 표현형을 관찰함으로써 AVP1 유전자의 농업적 활용가능 성을 확인하였다.
재료 및 방법
배추 유전자형, 기내 파종, 형질전환용 절편 조제
경북 포항에서 수집한 재래종 배추(Brassica rapa L.)를 포 트에 파종하여 본엽 4 ~ 8매기에 춘화처리(7℃, 3주)를 실 시하고 상온의 생장상 또는 온실에서 추대와 개화를 유도 한 후 인공 자가 뇌수분을 실시함으로써 개체별로 다음세 대 종자를 얻었다. 동일한 방식으로 각각 5세대를 진전 (변형 1개체 1계통육종법: single seed descent method)시켜 유묘기 원예적 특성이 양호하고 표현형 비분리가 명확한
‘KSSD2810’ 고정계통을 본 연구의 재료로 사용하였다.
배추 종자를 2% CloroxTM(유효 염소 4% 이상) 용액에서 표 면살균 한 후 sucrose(Duchefa Biochemie, The Netherlands) 20 g/L와 gelrite(Sigma-Aldrich, USA) 4 g/L가 첨가된 1/2× MS 배지(Duchefa Biochemie)(pH 5.8)에 파종하여 22±2℃에 서 암배양하였다. 파종 4일 후 하배축 절편을 채취하여 Agrobacterium 공동배양 기법을 적용한 형질전환용 재료 로 사용하였다.
플라스미드, Agrobacterium 균주, AVP1 형질전환
애기장대의 액포막 소재 H+-PPase를 coding하는 유전자(AVP1:
NCBI accession #NM 101437)와 서로 역방향으로 Pnos::Bar construct(Xiang et al. 1999; supplied by Dr. Sunghun Park at Kansas State Univ.)를 GUS와 Bar 유전자 발현 cassette가 제거된 식물형질전환용 벡터 pCAMBIA3301(http://www.
cambia.org)의 제한효소 Xho I과 Bst EII site 사이에 재조합 하였다. AVP1은 pRT103(Töpfer et al. 1987)에서 cloning한 Cauliflower mosaic virus(CaMV) Cabb B-D strain의 35S promoter double enhancer와 pCAMBIA3301의 CaMV35S ployA에 의해 조절된다(Fig. 1A). 재조합된 plasmid는 freeze-thaw method (Holsters et al. 1978)를 통해 A. tumefaciens LBA4404 strain 에 도입하였고 이를 배추 내혼계통 ‘KSSD2810’의 하배축 절편을 이용한 형질전환에 사용하였다.
채취한 하배축 절편을 sucrose 20 g/L, NAA(Sigma-Aldrich) 1 mg/L, BAP(Sigma-Aldrich) 4 mg/L 및 plant agar(Duchefa Biochemie) 9 g/L가 첨가된 MS배지(pH 5.2)에서 16 h(50 μ mol・m-2・s-1) 일장 조건으로 2일간 전배양한 후 Agrobacterium 균주(OD600=0.5 ~ 0.6)를 접종하여 동일한 배지에서 2일간 암배양하였다. 공동배양한 후 cefotaxime(Duchefa Biochemie) 500 mg/L, AgNO3(Sigma-Aldrich) 4 mg/L 및 DL-phosphinothricin (PPT; Duchefa Biochemie) 5 mg/L가 추가 첨가된 선발배지 에 이식한 후 16 h(50 μmol・m-2・s-1) 일장에서 3주 간격으 로 계대배양 하였다. 약 1개월 후부터 재분화된 신초는 절취하여 sucrose 20 g/L와 plant agar 9 g/L이 첨가된 MS 배지(pH 5.8)에 이식하여 발근을 유도한 후 발근된 유식
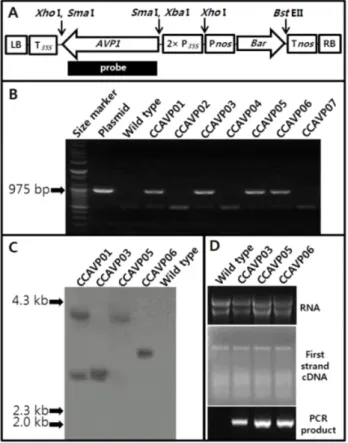
Fig. 1 Schematic map of the T-DNA of transformation vector and nucleic acids analysis of AVP1-expressing Chinese cabbage plants. (A) T-DNA map. LB: left border, T35S: Cauliflower mosaic virus (CaMV) 35S terminator, AVP1: cDNA encoding vacuolar H+-pyrophosphatase of Arabidopsis thaliana, 2× P35S: CaMV double 35S promoter, PNOS: nopaline synthase promoter, Bar: phosphinothricin resistance gene, TNOS: nopaline synthase terminator, RB: right border, probe: full length AVP1 cDNA restricted by Sma I for Southern blot analysis. (B) PCR detection of AVP1 genes in the putative transgenic Chinese cabbage regenerants. (C) Southern blot analysis of the PCR-positive T0 plants. (D) Detection of AVP1 transcripts by RT-PCR.
물체는 Han 등(2004)의 방법에 따라 기외 이식하여 순화 시켰다.
식물체의 핵산분석
유식물체의 유연한 잎을 채취한 후 DNeasy Plant Mini kit (Qiagen GmBH, German)을 이용하여 genomic DNA(gDNA) 를 추출하였고 gDNA와 forward(5’-gtcggcgctgaccttgtcggtaaa-3’) 및 reverse(5’-ccaatgataactttaggggtcaaa-3’) primer 를 각각 최 종농도 0.8 ng/μL와 200 µM이 되도록 PerfectShot Ex Tag Kit(Takara Bio Inc., Japan)에 첨가하여 중합효소연쇄반응 (polymerase chain reaction: PCR)을 수행하였다. 더불어 Southern 혼성화 분석을 수행하였는데 식물체 gDNA의 절단은 T- DNA를 1회 절단하는 제한효소 Xba I을, plasmid로부터 probe 제작을 위해서는 제한효소 Sma I을 사용하였으며(Fig. 1A) 절단된 gDNA의 분획, 분획된 gDNA의 membrane 전이 및
probe와의 혼성화 등은 Park 등(2005a)의 방법을 따랐다.
한편, 형질전환 식물체에서 AVP1 유전자의 전사체 측정 을 위하여 역전사중합효소연쇄반응(reverse transcriptional polymerase chain reaction: RT-PCR)분석을 수행하였다. RT-PCR 분석을 위해서 Han(2010)의 방법을 준용하였는데, 추출된 RNA는 역전사 전에 DNA 오염을 제거하기 위하여 DNase I(Invitrogen, USA)으로 2회 처리하였고 분광광도계(NanoDrop 2000: Thermo Fisher Scientific Inc., USA)로 정량한 후 시료 당 등량의 RNA를 cDNA 합성용 주형으로 사용하였으며 합성된 cDNA는 전기영동을 통해 합성량을 확인하는 등 준정량적 실험기법을 적용하였다.
AVP1 동형접합 T2 세대 육성
AVP1 유전자가 1 copy 삽입되고 정상적으로 전사되는 T0 재 분화 식물체를 본엽 6 ~ 7매 시점에 냉장진열장(ADVANZA:
Carrier, Korea)에서 8±3℃, 16 h(약 20 μmol・m-2・s-1) 조건으 로 3주간 춘화처리 시켰다. 춘화처리된 식물체는 상온으 로 이동시켜 추대시킨 후 유산지 봉투로 꽃대를 씌워 꽃 가루 오염을 방지하였으며 각 개체에서 꽃이 피면 꽃가 루를 채취하여 자가 뇌수분 시켰다. 각 개체에서 T1 세대 종자를 채취하여 유식물체를 육성한 후 PCR 분석을 수 행하였고 목표 유전자의 증폭이 확인된 개체는 다시 춘 화처리와 자가 뇌수분 및 채종하여 T2 집단을 육성하였 다. T2 세대 각 집단의 유식물체를 대상으로 다시 PCR 분 석을 수행하여 AVP1 유전자의 분리 여부를 판단하였다.
고정된 T2 집단은 염스트레스 하에서의 생리특성 분석을 위한 재료로 사용하였다.
염스트레스 처리
Petri접시에 고정된 T2 세대 각 집단을 파종하고 탈이온수 를 공급하여 2일간 최아시킨 후 유근의 생장이 유사한 개체만을 골라 각 hole에 상토(High: Pu-nong Co., Korea) 100 mL를 담은 32공 plug tray에 이식하였다. 배양실(25±1℃, 16h 일장)에서 평균 5일 마다 탈이온수로 포장용수량까지 저면관수하며 15일간 식물체를 생장시킨 후 염용액의 공 급을 시작하였다. MS salt는 1×[electric conductivity(EC)=5.0 mS/cm], 2×(EC=9.5 mS/cm), 4×(EC=17.8 mS/cm) 및 8×(EC=31.6 mS/cm) 순으로, NaCl은 50 mM(EC=5.0 mS/cm), 100 mM (EC=9.6 mS/cm), 200 mM(EC=17.6 mS/cm) 및 400 mM (EC=31.0 mS/cm) 용액 순으로 각각 2일 간격으로 포장용 수량까지 저면관수하였다.
광합성 효율 측정을 위한 엽록소형광소멸분석
AVP1 형질전환체와 wild-type 유식물체의 염스트레스 처
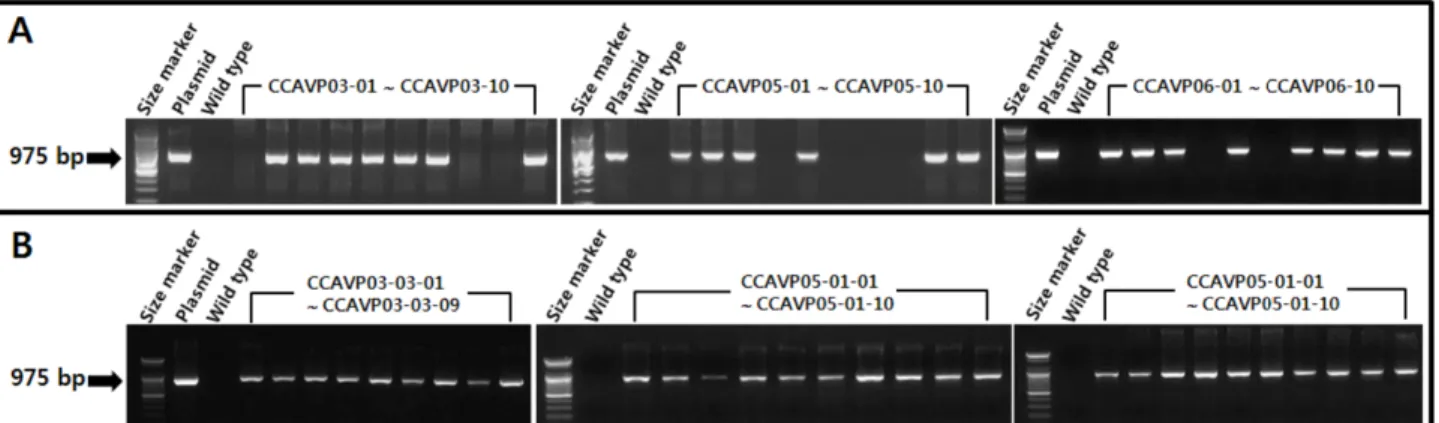
Fig. 2 PCR detection of AVP1 genes in the Chinese cabbage seedlings at T1 and T2 generation. (A) Segregation pattern of AVP1 gene by showing PCR in the three T1 populations corresponding with a Mendelian fashion. (B) AVP1-fixed T2 lines confirmed by PCR analysis.
리에 따른 광합성 효율을 비교하기 위하여 각 농도의 염 용액 공급직전 WinControl software(version 2.08: Heinz Walz GmbH, Germany)가 구동하는 pulse amplitude modulation fluorometer(Mini-PAM: Heinz Walz GmbH)를 활용하여 포 화광 pulse quenching하에서 광계II(photosystem II: PSII) 유 래 형광(Chl-F)을 측정하였다. 유식물체의 제2엽에 Dark Leaf Clip(DLC-8: Heinz Walz GmbH)을 각각 고정하여 30 분 이상 암적응 시킨 후 measuring light intensity 3, actinic light intensity 1 및 saturation pulse intensity 8의 조건으로 20 초 간격으로 14회 quenching을 수행하며 각 induction curve 를 작성한 후 steady stationary phase에서 형광값에 의존하 는 매개변수를 취하여 PSII quantum yield(φPSII: Genty et al.
1989)를 산출하였다. φPSII는(Fm’-Ft)/Fm’식으로 구하였는 데 Fm’은 명적응된 상태에서의 최대 Chl-F, Ft는 명적응 된 상태에서의 포화광 pulse 조사 직전 Chl-F 값을 의미한 다(Han 2010).
DAB 염색
염스트레스에 의해서도 발생할 수 있는 활성산소족의 하 나인 H2O2를 탐지하기 위하여 DAB(3,3’-diaminobenzidine;
Sigma-Aldrich) 염색을 실시하였다(Wu et al. 2012). 8× MS salt 공급 직전(최초 염 공급 후 6일)에 각 유식물체의 제3엽 을 채취하여 0.1% DAB(pH 3.8) 용액에 침지한 후 25±1℃, 암 조건으로 1시간 동안 처리하였다. DAB 흡수 처리 후 7 0℃로 데워진 94% 에탄올로 4회에 걸쳐 탈색시켰으며 해부현미경 하에서 조직내 침착된 갈색의 H2O2-DAB 반 응산물을 관찰하였다.
수세를 통한 용토 탈염
최종 염처리인 8× MS salt 또는 400 mM NaCl 공급 8일 후 염스트레스 처리를 받은 식물체가 심겨진 plug tray를
4회에 걸쳐 수돗물로 저면관수와 배수를 반복함으로써 용토 내 염을 제거하였다. 용토 수세 후 유묘를 배양실에 서 정상재배하며 생육과 회복 상태를 관찰하였다.
결과 및 고찰
AVP1 유전자 발현 배추 고정계통 육성
배추 내혼계통 ‘KSSD2810’의 하배축 절편을 재료로 사용 하고 Agrobacterium 공동배양 기법을 적용하여 AVP1 유전 자가 도입된 형질전환 T0 식물체를 획득한 후 세대진전 하여 T2 고정계통을 선발하였다. 재분화된 7개 식물체 중 4개체에서 AVP1 유전자 유래 975 bp DNA 단편이 증폭되 었으며(Fig. 1B), Southern 혼성화 분석을 통해 각각 1, 2 copy의 AVP1을 탐지하였다(Fig. 1C). 또한 AVP1이 1 copy 삽입된 것으로 판단되는 형질전환 식물체(‘CCAVP03’,
‘CCAVP05’, ‘CCAVP06’)의 경우 그 양에 차이가 있기는 하지만 정상적으로 전사체를 발현하였다(Fig. 1D). 본 연 구에서의 형질전환 효율은 약 0.6%(자료 미제시)로 기 보 고(Park et al. 2005a; Takasaki et al. 1997)된 효율에 미치지 못하였는데, 이는 Brassica rapa 종 자체가 에틸렌 등의 영 향을 받아 재분화 또는 형질전환이 어렵고 사용한 유전 자형과 배양조건이 달랐기 때문(Takasaki et al. 1997, 2004)으로 사료된다.
1 copy AVP1 삽입 형질전환체를 자가 뇌수분시켜 획득 한 T1 세대 3개 집단에 대한 PCR 분석 결과, 각 집단에서 AVP1 유전자는 멘델식 3:1 분리비(각각의 적합도 P=0.50 ~
0.95, P=0.20 ~ 0.50, P=0.50 ~ 0.95)에 근접하였다(Fig. 2A).
T1 세대 각 집단별로 PCR 증폭산물이 확인된 5개체씩을 자가 뇌수분 시켜 T2 집단을 양성한 후 다시 PCR 분석을 수행하여 AVP1 유전자가 고정된 독립된 3개 T2 집단을 계통으로 최종 선발하였고(Fig. 2B) 이를 유식물체 바이
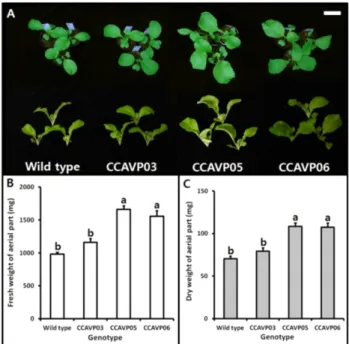
Fig. 3 Increased biomass of the aerial part in AVP1-expressing Chinese cabbage plants grown under normal condition. (A) Morphology of aerial parts of transgenic and wild-type young plants. (B and C) Fresh and dry weight of the aerial parts in different genotype plants. Data represent the mean values (±SE) obtained from five independent young plants. Values in each frame followed by different letter are significantly different at the 5% level (t-test;
n=5).
오매스 비교와 염스트레스에 대한 유묘 생리검정에 사용 하였다.
AVP1 형질전환 식물체의 바이오매스
AVP1 형질전환 계통과 wild-type은 파종 후 17일에 본엽 2 ~ 3매까지 생장하였는데(Fig. 3A), 지상부 바이오매스에 있어 유전자형간 뚜렷한 차이를 나타내었다. ‘CCAVP05’
는 wild-type에 비해 지상부의 생체중과 건물중이 각각 69.6%
와 53.5% 증가하였고, ‘CCAVP06’은 wild-type에 비해 지 상부 생체중과 건물중이 각각 58.8%와 52.3% 증가하였 다. 반면 ‘CCAVP03’은 wild-type과 비교하여 지상부 생체 중과 건물중에 유의한 차이가 없었으며 각 유전자형의 지상부 수분함량 역시 92.8% ~ 93.5%로 차이가 없었다(Fig.
3B, 3C). AVP1 유전자 과발현에 따른 지상부 또는 근권부 바이오매스 증가는 애기장대, 토마토, 벼 및 목화 등 다 양한 종에서 알려져 있는데(Li et al. 2005; Park et al. 2005b;
Pasapula et al. 2011; Yang et al. 2007), AVP1 과발현이 세포 질 pH 변화를 초래하고 이어 P-type H+-ATPaes를 비롯한 다른 이온수송체가 활성화되며 pH 변화에 수반하여 오 옥신 수송량의 증가와 근권부 산성화에 따른 인산 흡수 증가 때문인 것으로 보고된 바 있다(Gonzalez et al. 2010;
Li et al. 2005; Park et al. 2005b; Pasapula et al. 2011; Yang
et al. 2007).
염스트레스 하에서 AVP1 형질전환 식물체의 광합성 효율과 H2O2 탐지
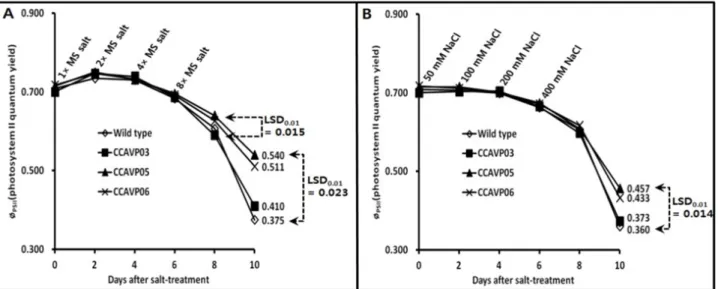
염스트레스 하에서 식물체는 막투과성의 변화로 인한 전 해질 누출과 활성산소족(ROS: ・O2-, H2O2, OH radical 등) 의 생성 뿐만 아니라 광합성 효율 저하 등을 수반하게 된 다(Dionisio-Sese and Tobita 2000; Wang et al. 2003; Zhu 2001, 2003). 본 연구에서는 점진적으로 농도를 높인 MS salt와 NaCl 용액을 용토에 투여하여 조성되는 근권부 염 스트레스 하에서 AVP1 형질전환 식물체의 광합성 효율 을 엽록소형광소멸분석을 통해 추적하였다. 염스트레스 처리 직전인 파종 후 17일 φPSII이 0.700 ~ 0.717로 형질전 환 계통과 wild type간 구별되지 않았지만 MS salt 처리구 는 8일 후(8× MS salt 공급 2일 후), NaCl 처리구는 10일 후(400 mM NaCl 공급 4일 후)부터 유전자형에 따라 확연 한 차이를 나타내었다(Fig. 4A, 4B). 각 염스트레스 처리 6일 후부터 모든 유전자형에서 φPSII가 급하게 저하되었 지만 ‘CCAVP05’와 ‘CCAVP06’은 wild-type이나 ‘CCAVP03’에 비해 상대적으로 기울기가 완만하였다. 특징적인 것은 NaCl 처리구와는 다르게 MS salt 처리구에서 모든 유전 자형이 염처리 4일까지는 φPSII가 염처리 전에 비해 일시 적으로 높았는데(Fig. 4A), 이는 빈영양염 상태의 용토에 서 생장한 유묘에 대한 시비의 효과인 것으로 판단된다.
염스트레스가 광합성 효율에 미치는 영향에 관하여 비교 적 연구가 많이 진행된 벼의 경우에도 형질전환 계통 또 는 비형질전환 품종을 막론하고 유전자형과 염스트레스 의 강도에 따라 광합성 효율 저하의 차이가 분명하게 나 타난다(Dionisio-Sese and Tobita 2000; Hoshida et al. 2000;
Sakamoto et al. 1998; Uchida et al. 2002).
다양한 스트레스에 의해 식물체에서 생성될 수 있는 활 성산소족의 하나인 H2O2를 DAB 염색을 통해 탐지할 수 있 다(Guan et al. 2012; Wu et al. 2012). MS salt 처리 6일 후 채 취한 잎을 DAB로 염색한 결과, wild-type과 ‘CCAVP03’의 엽 병 관다발조직에서만 갈색의 H2O2-DAB 반응산물이 확인되 었다(Fig. 5). 이러한 결과는 염스트레스를 받은 특정 시점 에 wild-type과 ‘CCAVP03’에는 ‘CCAVP05’와 ‘CCAVP06’에 비해 체내에 더 많은 H2O2가 존재하고, 따라서 ‘CCAVP05’와
‘CCAVP06’는 고염에 의해 유발되는 산화스트레스에 대 한 내성이 더 강하다고 판단할 수 있다. 그러나 본 실험 에서 H2O2-DAB 반응산물이 관찰되지 않았다고 해서 염 스트레스 하에서도 ‘CCAVP05’와 ‘CCAVP06’에서는 H2O2가 생성되지 않는다고 보기는 어렵다. 왜냐하면 ‘CCAVP05’와
‘CCAVP06’도 광합성 효율 측면에서 이미 장해를 받고 있 으며(Fig. 4A) 식물체의 비생물스트레스 내성 발현 네트 워크에는 이미 생성된 H2O2의 분해를 촉매하는 catalase를
Fig. 4 Kinetics of photosystem II quantum yield (φPSII) in the AVP1-expressing Chinese cabbage plants subjected to salt stress. (A and B) φPSII slope with the lapse of time in the plants grown under gradually intensified salt stress using MS salt and NaCl. Data represent the mean values obtained from the five independent plants. Values for least significant difference at the 1% level (LSD0.01; n=5) were directly indicated in each frame.
Fig. 5 Effect of stress set by gradually intensifying MS salt on the accumulation of H2O2 in the vascular bundle tissues of AVP1- expressing leaf petioles. Each arrow according to size indicates the different intensity of H2O2-DAB reaction product.
포함한 다양한 활성산소족 소거 효소계와 비효소계가 포 함되어 있기 때문이다(Wang et al. 2003).
수세를 통한 용토 탈염 후 회복 반응
최종농도의 MS salt와 NaCl 공급 8일 후(용토 수세 직전) 모든 유전자형의 잎에 괴사 반점이 수반된 위조현상이 나 타나기는 하였지만 ‘CCAVP05’와 ‘CCAVP06’은 wild-type과
‘CCAVP03’에 비해 상대적으로 그 정도가 경미하였다(Fig.
6). 즉, AVP1이 형질전환된 ‘CCAVP05’와 ‘CCAVP06’ 두 계 통은 연속적으로 강화되는 염스트레스 하에서 wild-type에 비해 외형적으로도 강한 내성을 나타낸다고 판단 할 수 있다. 염이 축적된 용토를 수세하여 회복을 유도한 2주 후 wild-type과 ‘CCAVP03’의 내, 외엽은 모두 고사하였지 만 대부분의 ‘CCAVP05’와 ‘CCAVP06’ 식물체에서는 외엽
은 고사한 반면 내엽은 새롭게 생장하였다(Fig. 6). 특징적 인 것은 MS salt로 강화된 스트레스 환경에서는 ‘CCAVP05’
와 ‘CCAVP06’이 확연하게 wild-type과 구분되어 내엽이 분 화되었지만 NaCl로 강화된 스트레스 환경에서는 내엽 분 화의 정도가 미미하였다(Fig. 6). 이러한 결과는 지금까지 다른 작물에서 내염성 획득을 목표로 형질전환체를 획득 한 후 생물검정을 수행할 때 대부분 NaCl을 투여하여 염 스트레스 환경을 부여하고 있는 상황에 반하여 중요한 의미를 가진다고 판단된다. 실용적 목적을 가지고 내염 성 형질전환 이벤트를 개발코자 할 때는 간척지와 염습 지를 제외하고는 대부분의 농경지에 축적되는 염의 종류 가 단순하지 않다는 점을 염두에 두어야 할 것으로 사료 된다.
지금까지 연구결과에서 ‘CCAVP03’은 비록 목표유전자 인 AVP1을 가지고 있고 전사체도 발현하지만(Fig. 1B, 1C)
Fig. 6 Different response of the plants subjected to imposed salt stresses. Phenotypes of plants with different genotypes before and after desalinization for recovery are shown. Short description on treatments is directly indicated in each panel.
‘CCAVP05’와 ‘CCAVP06’과는 달리 wild-type과 비교하여 바이오매스, 염스트레스에 대한 생리적 반응 및 회복반 응에 있어 큰 차이를 보이지 않았다. 이는 명쾌하지는 않 지만 AVP1 유전자 1 copy 삽입이 불명확(Fig. 1C)하거나 배추 게놈에 AVP1과 유사한 내생 유전자가 존재하여 발생 하는 post-transcriptional gene silencing(PTGS), 또는 T-DNA 삽입의 위치 효과(transgene positional effect) 등에 기인한 것으로 추측할 수 있을 것이다(Hammond et al. 2001; Stam et al. 1997). 또한, 본 연구에서는 AVP1 형질전환 유식물 체만을 대상으로 바이오매스 증가와 내염성을 검증하였 다. 농업적 측면에서의 활용가능성을 좀 더 명확히 하기 위해서는 인공적이건 염류가 축적된 농가 포장이건 성숙 식물체 단계까지 AVP1 발현에 따른 표현형을 추적 관찰 하는 것과 기 보고된 AVP1 발현이 근권부 바이오매스와 건조스트레스에 미치는 효과(Park et al. 2005b; Yang et al.
2007)에 대한 추가적 관찰이 필요하다고 판단된다.
적 요
애기장대 액포 소재 H+-PPase(AVP1)의 과발현이 농업적 으로 가치 있는 표현형을 나타낸다는 기 보고에 기초하 여, AVP1 발현이 다른 종에서도 일관되게 바이오매스를 증가시키고 염에 대한 내성을 향상시키는지를 확인하기 위하여 본 연구에서는 AVP1 형질전환 배추 식물체를 획 득한 후 고정계통을 육성하여 생리검정 재료로 사용하였 다. 형질전환 배추 유식물체는 비형질전환 유식물체에 비해 생장이 왕성하였으며 염스트레스에 대한 내성도 강 하였다. 정상 재배조건에서 생장시킨 유식물체의 생체중
과 건물중을 비교함으로써 형질전환에 의한 바이오매스 증가 표현형을 확인하였으며 MS 염과 NaCl로 점차 염스 트레스를 강화시키는 조건에서 광계II 양자수율을 추적, DAB 염색 실시 및 최종적으로 용토 탈염 후 회복 실험을 수행함으로써 내염성 향상 표현형을 확인하였다.
사 사
본 연구는 농촌진흥청 차세대 바이오그린21사업(과제번 호: PJ008324)의 지원에 의해 이루어진 것임. 이 논문은 2009년도 정부(교육과학기술부)의 재원으로 한국연구재단 의 기초연구사업 지원을 받아 수행된 것임(2009-0064836).
인용문헌
Apse MP, Aharon GS, Snedden WA, Blumwald E (1999) Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285:1256-1258
Dionisio-Sese M, Tobita S (2000) Effects of salinity on sodium content and photosynthetic responses of rice seedlings differing in salt tolerance. J Plant Physiol 157:54-58
Gaxiola RA, Li J, Undurraga S, Dang LM, Allen GJ, Alper SL (2001) Drought- and salt-tolerant plants results from over- expression of the AVP1 H+-pump. PNAS 98:11444-11449 Genty B, Briantais JM, Baker NR (1989) The relationship between
quantum yield of photosynthetic electro transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 990:87-92 Gonzalez N, De Bodt S, Sulpice R, Jikumaru Y, Chae E, Dhondt S et al. (2010) Increased leaf size: Different means to an end.
Plant Physiol 153:1261-1279
Greenway H, Munns R (1980) Mechanisms of salt tolerance in nonhalophytes. Annu Rev Plant Physiol 31:149-190
Guan Q, Takano T, Liu S (2012) Genetic transformation and analysis of rice OsAPx2 gene in Medicago sativa. PLOS ONE 7:1-8
Hammond SM, Caudy AA, Hannon GJ (2001) Post-transcriptional gene siliencing by double-stranded RNA. Nature Genetics 2:110-119
Han (2010) Suppression of tobamovirus movement toward upper leaves in the tomato plant over-expressing a maize calreticulin.
J Plant Biotechnol 37:567-573
Han JS, Oh DG, Mok IG, Park HG, Kim CK (2004) Efficient plant regeneration from cotyledon explants of bottle gourd (Lagenaria siceraria Standl.). Plant Cell Rep 23:291-296
Hirschi K (2001) Vacuolar H+/Ca2+ transport: who’s directing the traffic? Trend Plant Sci 6:100-104
Holsters M, De Waele D, Depicker A, Messens E, Van Montagu M, Schell J (1978) Transfection and transformation of A.
tumefaciens. Mol Gen Genet 163:181-187
Hoshida H, Tanaka Y, Hibino T, Hayashi Y, Tanaka A, Takabe T et al. (2000) Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase.
Plant Mol Biol 43:103-111
Kasuga M, Miura S, Shinozaki K, Yamaguchi-Shinozaki K (2004) A combination of the Arabidopsis DREB1A gene and stress- inducible rd29A promoter improved drought- and low- temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol 45:346-350
Li J, Yang H, Peer WA, Richter G, Blakeslee J, Bandyopadhyay A et al. (2005) Arabidopsis H+-PPase AVP1 regulates auxin- mediated organ development. Science 310:121-125
Liu J, Ishitani M, Halfter U, Kim CS, Zhu JK (2000) The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. PNAS 97:3730-3734
Liu J, Zhu JK (1998) A calcium sensor homolog required for plant salt tolerance. Science 280:1943-1945
Park BJ, Liu Z, Kanno A, Kameya T (2005a) Genetic improvement of Chinese cabbage for salt and drought tolerance by constitutive expression of a B. napus LEA gene. Plant Sci 169:553-558
Park S, Li J, Pittman JK, Berkowitz GA, Yang H, Undurraga S et al. (2005b) Up-regulation of a H+-pyrophosphatase (H+-PPase) as a strategy to engineer drought-resistant crop plants. PNAS 102:18830-18835
Pasapula V, Shen G, Kuppu S, Paez-Valencia J, Mendoza M, Hou P et al. (2011) Expression of an Arabidopsis vacuolar H+- pyrophosphatase gene (AVP1) in cotton improves drought- and salt tolerance and increases fibre yield in the field conditions. Plant Biotech J 9:88-99
Qiu QS, Guo Y, Dietrich MA, Schumaker KS, Zhu JK (2002) Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2. PNAS 99:8436-8441 Sakamoto A, Murata A, Murata N (1998) Metabolic engineering of
rice leading to biosynthesis of glycinebetaine and tolerance to salt and cold. Plant Mol Biol 38:1011-1019
Sanders D, Pelloux J, Brownlee C, Harper JF (2002) Calcium at the crossroads of signaling. Plant Cell 14 Suppl.:S401-S417 Sarafian V, Kim Y, Poole RJ, Rea PA (1992) Molecular cloning
and sequence of cDNA encoding the pyrophosphate-energized vacuolar membrane proton pump of Arabidopsis thaliana.
PNAS 89:1775-1779
Stam M, Mol JNM, Kooter JM (1997) The silence of genes in transgenic plants. Ann Bot 79:3-12
Takasaki T, Hatakeyama K, Hinata K (2004) Effect of silver nitrate on shoot regeneration and Agrobacterium-mediated transfor- mation of turnip (Brassica rapa L. var. rapifera). Plant Biotechnol 21:225-228
Takasaki T, Hatakeyama K, Ojima K, Watanabe M, Toriyama K, Hinata K (1997) Factors influencing Agrobacterium-mediated transformation of Brassica rapa L. Breed Sci 47:127-134 Töpfer R, Matzeit V, Gronenborn B, Schell J, Steinbiss HH (1987)
A set of plant expression vectors for transcriptional and translational fusions. Nucleic Acids Res 15:5890(abstr.) Uchida A, Jagendorf AT, Hibino T, Takabe T, Takabe T (2002)
Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci 163:515-523
Wang W, Vinocur B, Altman A (2003) Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta 218:1-14
Wu Q, Lin J, Liu JZ, Wang X, Lim W, Oh M et al. (2012) Ectopic expression of Arabidopsis glutaredoxin AtGRXS17 enhances thermotolerance in tomato. Plant Biotechnol J 10:945-955 Yang H, Knapp J, Koirala P, Rajagopal D, Peer WA, Silbart LK et
al. (2007) Enhanced phosphorus nutrition in monocots and dicots over-expressing a phosphorus-responsive type I H+- pyrophosphatase. Plant Biotech J 5:735-745
Xiang C, Han P, Lutziger I, Wang K, Oliver DJ (1999) A mini binary vector series for plant transformation. Plant Mol Biol 40:711-717
Zhen RG, Kim EJ, Rea PA (1997) Acidic residues necessary for pyrophosphate-energized pumping and inhibition of the vacuolar H+-pyrophosphatase by N,N ’-dicyclohexylcarbodiimide.
J Biol Chem 272:22340-22348
Zhu JK (2001) Plant salt tolerance. Trend Plant Sci 6:66-71 Zhu JK (2003) Regulation of ion homeostasis under salt stress.
Curr Opin Plant Biol 6:441-445