85
저산소 환경에서의 유전자 발현 조절
윤 여 민ㆍ하 윤
연세대학교 의과대학 Brain Korea 21 PLUS Project, 연세대학교 의과대학 신경외과학교실, 척추신경연구소
Controlled Gene Expression System under Hypoxia Conditions
Yeomin Yun, M.S., Yoon Ha, M.D., Ph.D.
Brain Korea 21 PLUS Project for Medical Science, College of Medicine, Yonsei University, Department of Neurosurgery, Spine & Spinal Cord Institute, College of Medicine, Yonsei University, Seoul, Korea
Purpose: To understand the phenomenon of gene expression under hypoxia conditions, hypoxia-inducible vector system was constructed. Hypoxia-inducible enhancer and 3’ untranslated region was responsible to overexpress so that regulates gene expression under hypoxia conditions. Methods: We used several kinds of hypoxia-inducible factors to regulate gene expression under hypoxia conditions. Erythropoietin (Epo) enhancer and 3’Epo-untranslate region (Epo-3’-UTR) was used respectively upstream or down- stream of SV promoter. Mouse neural stem cells (mNSCs) transfected each vector was cultured under normoxia or hypoxia conditions. Results: Luciferase assay and ELISA revealed that gene expression level under hypoxia conditions was higher than normoxia conditions. Plasmid containing both Epo enhancer and Epo-3’-UTR increased the best compared to normoxia conditions. However, Epo enhancer showed the highest gene expression under hypoxia conditions and relative to SV promoter. Conclusions: As a result of gene expression level under hypoxia conditions compared to normoxia conditions, all of those 3 kinds vector have an effect on overexpression. On the basis of that, we can regulate gene expression under hypoxia conditions so that it can be applied to improve gene or cell therapy.
Key words: Hypoxia, Gene expression regulation, Hypoxia-inducible, Enhancer
접수: 2015년 11월 16일, 심사완료일: 2015년 12월 8일 교신저자: 하 윤
우 03722 서울시 서대문구 신촌로 50 연세대학교 의과대학 신경외과학교실 Tel: 02 2228 2808, Fax: 02 393 9979 E mail: [email protected]
This work was supported by the Brain Korea 21 PLUS Project for Medical Science, Yonsei University.
I. 서 론
높은 고도의 항공환경에서는 저산소상태(hypoxia), 가속력 (acceleration), 감압력(depression), 미세중력(microgravity)이 주 요 환경변화 요인이다. 고도가 상승함에 따라 대기압은 감 소하고, 대기압 감소는 산소의 분압을 감소시킨다. 상공 약 2.4 km에서는 해수면의 75% 수준으로 산소 분압이 낮아져 경미한 저산소증 증상이 나타나기 시작하며 해수면 15 km 상공에서는 10%까지 산소 분압이 떨어진다[1,2]. 대기 중
산소 분압이 감소하면 신체 내로 산소를 흡수하는 능력 또 는 산소를 운반하는 능력이 떨어져 산소에 대한 요구도가 높아진다[3]. 감소한 산소 분압에 따라 발생하는 저산소증 이란 조직 내의 산소결핍상태로서 특히 중추신경계가 매우 취약하다. 저산소증은 조직독성(histotoxic), 과혈류성(hyperemic), 울혈성(stagnant), 저산소성(hypoxic) 저산소증 4가지로 분류 할 수 있다. 조직독성(histotoxic) 저산소증은 세포가 조직 내 에 도달한 산소를 이용하는 능력이 저하되는 것이고 과혈 류성(hyperemic) 저산소증은 적혈구가 일산화탄소 중독과 같이 산소를 운반하는 능력이 떨어져 발생한다. 울혈성(stagnant) 저산소증은 전신 또는 국소적으로 혈류량이 감소하는 경우 이고 저산소성(hypoxic) 저산소증은 폐포내의 산소분압 감소 로 인해 일어나며 항공환경에서 가장 흔하게 발생하는 현 상으로 10,000피트 이상으로 올라가게 되면 피로, 두통, 의 식소실, 운동조절 기능 이상 및 중추신경계 이상이 초래된 다. 본 연구에서는 해수면과 달리 비정상적인 항공환경 요 인 중 하나인 저산소증과 저산소 환경에서의 유전자 발현
메커니즘을 이해함으로써 항공환경에서 취약한 신경계를 보호할 수 있는 유전자 발현 조절에 대해 다루고자 한다.
저산소 환경에서 발현되는 혈관생성성장인자(angiogenic growth factor), 항세포사멸 유전자(anti-apoptotic gene), 해당유 전자(glycolytic gene) 등은 전사, 전사 후, 번역 후 과정에서 발현이 조절된다[4-6]. 저산소유도인자-1(hypoxia-inducible factor-1; HIF-1)는 저산소 환경에서 활성화 되는데[5,7]
HIF-1α와 HIF-1β 두 가지 subunit으로 구성되어 있다.
HIF-1β는 산소 농도에 관계없이 일정한 단백질 레벨을 유 지하는데 반해 HIF-1α는 정상 산소 조건에서는 분해되고, 저산소 조건에서는 단백질이 축적되어 HIF-1β와 hetero- dimer를 형성한다[8-10]. 축적된 HIF-1은 유전자의 promoter와 enhancer에 구성된 저산소반응요소(hypoxia response element;
HRE)와 결합하여 유전자의 전사를 촉진시킨다[6,11,12]. 이 메커니즘이 저산소 환경에서 발현이 촉진되는 유전자의 전 사에 의한 조절이다. 전사 후 조절은 mRNA를 안정화 시킴 으로써 이루어진다. mRNA는 5’과 3’에 비암호화부위(non- coding region)인 비해석부위 (untranslated region; UTR)를 갖고 있고, 저산소에서 유도되는 유전자는 특이적인 5’-, 3’-UTR 를 갖고 있어 저산소 환경에서 특정 단백질이 UTR에 결합 함으로써 mRNA를 안정화시킨다[13,14]. 안정화된 상태의 mRNA가 증가하여 효율적인 번역을 유도함으로써 저산소 조건에서 유전자의 발현이 증가한다.
에리트로포이틴(erythropoietin; Epo)은 저산소에서 발현이 유도되는 해당단백질이다. Epo enhancer는 HIF-1이 결합할 수 있는 HRE를 갖고 있어 저산소 환경에서 전사가 유도된 다. Epo RNA 결합단백질(Epo RNA binding protein; ERBP)은 Epo mRNA의 3’-UTR에 결합하여 Epo mRNA를 안정화 시킨 다. 안정화 상태의 mRNA가 증가함에 따라 효율적인 유전 자 번역이 유도되어 저산소 환경에서 단백질 발현을 증가 시킨다[13,15,16]. Epo의 enhancer와 3’-UTR을 이용하면 저산 소 환경에서 유전자의 전사, 전사 후 단계를 통해 목적 유전 자의 발현 조절이 가능하다.
이에 저자들은 Epo enhancer 그리고/또는 Epo-3’-UTR을 갖 고 있는 유전자 발현 시스템을 제작하여 마우스 유래 신경 줄기세포(mouse neural stem cell; mNSC)에 도입하였다. 정상 산소 조건과 저산소 조건에서 유전자 발현 양상을 비교하 기 위해 유전자가 도입된 세포는 정상산소 또는 저산소 환 경에서 배양하였다.
II. 재료 및 방법 1. 플라스미드 제작
SV promoter에 루시퍼라아제(luciferase)를 reporter gene으로 사
용한 벡터시스템 pSV-Luc을 기본으로 pEpoSV-Luc, pSV-Luc-UTR, pEpoSV-Luc-UTR을 재조합 하였다. pSV-VEGF, pEpoSV-VEGF를 제작하기 위해 luciferase gene을 VEGF로 재 조합 하였다. 제작된 벡터 시스템은 한양대 이민형교수님 께 제공받았다.
2. 유전자 도입
유전자를 도입하기 16시간 전에 10% fetal bovine serum (FBS; Thermo Scientific HyClone, Logan, UT, USA), 1%
penicillin/streptomycin (P/S; Gibco, Grand Island, NY, USA)가 포 함된 Dulbecco’s Modified Eagle Medium:Nutrient Mixture F-12 (DMEM/F12; Gibco, Grand Island, NY, USA) 세포 배양 배지를 이용하여 마우스 유래 신경 줄기세포(mouse neural stem cells;
mNSCs; CRL-2925, ATCC, Manassas, VA, USA)를 1×105 cells/well로 6 well plate에 분주한다. 1% P/S DMEM/F12 무혈 청 배지로 교환 후 Branched polyethylenimine (PEI 25000 D;
Sigma Aldrich, St. Louis, MO, USA)와 2ug DNA를 N/P ratio 5 비율로 혼합한 후 상온에서 10분간 반응 시킨 뒤 mNSCs 에 도입한다. 유전자가 도입된 세포를 정상산소 세포배양 기(20% O2) 또는 저산소 세포배양기(1% O2)에서 48시간동 안 배양한다.
3. 발광효소분석(luciferase assay)
배양 48시간 후, 0.5% trypsin EDTA를 이용하여 세포를 걷 어 1.5 ml tube에 옮긴다. 1,200 rpm에서 3분 간 원심분리 한 뒤 상층액은 버리고 세포 펠렛은 남긴다. 200 ul Pro-prep (iNtRON biotechnology, Gyeonggi-do, Korea)으로 세포 펠렛을 풀어주고 4oC에서 한 시간 동안 세포를 lysis시킨다. lysis된 세포를 4oC 원심분리기에서 13,000 rpm으로 10분간 원심분 리 한 후 상층액을 새로운 1.5 ml tube로 옮긴다.
Luminometer plate에 20 ul의 lysis된 세포를 넣고 100 ul의 Luciferase Assay Reagent (promega)를 첨가한 후 Microplate luminometer (Berthold, Bad Wildbad, Germany)를 이용하여 luciferase 발광량을 측정한다. 측정된 발광량을 BCA kit (pierce, Rockford, IL, USA)를 이용하여 측정한 총 단백질 농도 로 나누어 주고 이 때의 단위는 relative light units (RLU)/
milligram of total protein (mg)으로 나타낸다.
4. 효소결합 면역흡착 분석법(Enzyme linked immuno- sorbent assay; ELISA)
위와 같은 방법으로 pSV-VEGF, pEpoSV-VEGF를 mNSCs에 PEI를 이용하여 도입한 후 정산산소 세포배양기 또는 저산 소 세포배양기에서 48시간동안 배양한다. 48시간 후, 배지
Fig. 1. Construction of hypoxia-inducible gene expression system.
In order to express luciferase under hypoxia conditions, Epo enhancer and/or 3’-UTR was combined with SV promoter.
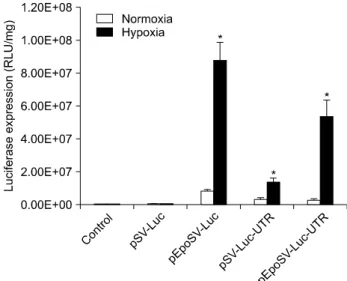
Fig. 2. Luciferase expression under normoxia or hypoxia conditions. Vectors including Epo enhancer and/or 3’-UTR showed higher gene expression under hypoxia conditions than normoxia conditions.
를 걷어 1200rpm에서 3분간 원심분리한 후 상층액을 모아
−20oC에 보관한다. 배지로 분비된 VEGF는 ELISA kit (Abfrontier, Seoul, Korea)를 이용하여 측정하였다. VEGF mono- clonal antibody가 코팅되어 있는 96well plate에 100ul/well로 샘플을 분주한 후 37oC에서 90분간 반응시킨다. 샘플을 blotting 방법으로 버린 후, VEGF antibody 를 100 ul/well로 분주한다. 37oC에서 60분간 반응시킨 후, 0.01 M phosphate buffered saline (PBS)로 washing 한다. Pre-warmed Avidin- Biotin-Peroxidase Complex (ABC)를 100 ul/well로 분주하여 37oC에서 30분간 반응시킨 후 같은 방법으로 PBS washing을 한다. 90 ul의 Tetramethylbenzidine (TMB) color developing agent를 첨가하여 빛을 차단하고 상온에서 20분간 반응 시 킨 후 TMB stop solution으로 반응을 종료시킨다. 샘플의 색 이 노란색으로 변하는 것을 확인한 후 450 nm 파장으로 O.D.값을 측정한다.
5. 통계학적 분석
데이터는 mean±SEM 값으로 나타내었다. 그룹 간 유전자 발현량 비교를 위해 t-test 검정을 이용하였다. P 값이 0.05 이하인 경우 유의성이 있는 것으로 판단하였다.
III. 결 과 1. 유전자 도입
유전자 발현을 위해 일반적으로 사용되는 SV promoter에 luciferase를 reporter gene으로 사용한 기본 벡터 pSV-Luc를 확 보하였다. 기본 벡터에 유전자의 전사단계를 조절하여 저 산소에서 발현을 증가시키는 Epo enhancer를 promoter의 upstream에 재조합하여 pEpoSV-Luc, 전사 후 단계를 조절하 는 Epo-3’-UTR을 재조합한 pSV-Luc-UTR, 두 가지를 모두 재 조합한 pEpoSV-Luc-UTR 세가지 벡터를 확보하였다(Fig. 1).
각 벡터를 신경줄기세포 (mNSCs)에 PEI를 이용하여 N/P ratio 5 값으로 도입하였다. 도입된 세포는 정산산소 세포배
양기 또는 저산소 세포배양기에서 48시간동안 배양한 후 발광효소분석을 통하여 유전자 발현량을 측정하였다.
2. 정상산소 또는 저산소 조건에서 발현한 발광효소 분석
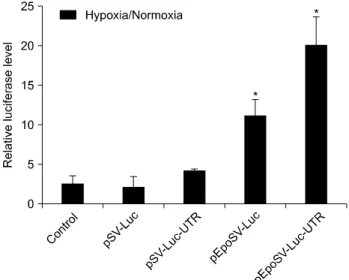
pSV-Luc, pEpoSV-Luc, pSV-Luc-UTR, pEpoSV-Luc-UTR에서 발현한 luciferase는 발광을 촉매하는 산화효소로서 luciferin 을 기질로 한다. lysis시킨 세포에 luciferin을 첨가함으로써 발현된 luciferase의 발광량를 측정할 수 있다. 발광량은 상대 적인 값이기 때문에 RLU/mg으로 나타낸다. 저산소 조건에 서 세포를 배양하였을 때, pEpoSV-Luc 벡터에서 가장 많은 발광량을 보였고, pSV-Luc-UTR 벡터에서 가장 낮은 발광량 을 보였다. Epo enhancer와 Epo-3’-UTR가 모두 존재한 벡터 의 발광량은 저산소 조건에서 증가하였지만 Epo enhancer만 있는 경우보다 낮게 발현하였다(Fig. 2). pSV-Luc에 비해 Epo enhancer가 존재한 경우 저산소 조건에서 약 205배 유전자의 발광량이 증가하였다. 하지만 Epo-3’-UTR이 있는 벡터에서 는 30배 증가, Epo enhancer와 Epo-3’-UTR이 동시에 존재한 벡터에서는 116배 증가로 Epo enhancer에 비해 적은 비율로 증가하였다(Fig. 3). Epo의 전사 후 조절보다 전사 단계 조절 을 이용하였을 때 더 효과적인 유전자 증가가 가능하다. 유 전자의 최종 발현량과 pSV-Luc에 비해 증가한 비율은 pEpoSV-Luc에서 가장 높았지만 정상산소 조건에 비해 증가 한 비율은 pEpoSV-Luc-UTR에서 가장 높았다. pSV-Luc-UTR 은 정상산소 조건에 비해 4배, pEpoSV-Luc는 10.5배 증가하 였고, Epo enhancer와 Epo-3’-UTR이 동시에 존재한 벡터의 경우 18.5배 증가하였다(Fig. 4). Epo enhancer와 Epo-3’-UTR
Fig. 3. Relative gene expression level compared to pSV-Luc showed 200 folds higher maximally.
Fig. 4. A vector including Epo enhancer and 3’-UTR both increased gene expression the most compared to normoxia conditions. Vectors including Epo enhancer or 3’-UTR showed 10 or 5 folds increased.
Fig. 5. Plasmid construction of VEGF therapeutic gene with Epo enhancer. Luciferase reporter gene in pSV-VEGF, pEpoSV-VEGF was replaced with the therapeutic VEGF gene to generate a hypoxia-inducible VEGF expression system.
Fig. 6. VEGF expression level under normoxia or hypoxia conditions. Gene expression was increased in the presence of Epo enhancer.
이 모두 존재할 때에 증가 비율은 가장 높아 지지만 가장 많은 유전자 발현을 보여준 것은 pEpoSV-Luc이다. pEpoSV- Luc 경우, 정상산소 조건에서도 다른 벡터에 비해 유전자 발현이 약 2.5배 높기 때문에 증가 비율에 비해 총 발현량이 가장 높다.
3. 저산소 환경에서 치료유전자 VEGF의 발현 조절
가장 효율적인 유전자 발현을 보여준 Epo enhancer를 이용 하여 치료 유전자인 혈관내피성장인자(Vascular endothelial
growth factor; VEGF)의 발현을 확인하기 위해 pSV-VEGF, pEpoSV-VEGF 벡터를 제작하였다(Fig. 5). ELISA로 확인한 저 산소 조건에서 VEGF 발현량은 pEpoSV-VEGF가 약 5200 pg/ml로 pSV-VEGF에 비해 25.5배, 정상산소 조건에 비해 4.4 배 증가하였다(Fig. 6). Luciferase를 이용하였을 때와 같은 결 과로 pEpoSV-VEGF 벡터는 정상산소 조건에서도 유전자 발 현이 증가하였다. VEGF는 혈관생성인자로 저산소 손상을 받았을 때 발현이 유도되는 것으로 알려져 있다. 혈관을 생 성함으로써 산소 공급을 하여 저산소에 의한 손상을 회복 시키는데 SV promoter 하에서 저산소에 의한 발현 증가를 보이지 않았다.
IV. 고 찰
저산소에서 발현하는 단백질의 특정 유전자 부위를 이용 하여 저산소 환경에서 목적 유전자의 발현을 유도하는 벡 터 시스템을 확립하였다. Epo enhancer를 이용하여 유전자의 전사 효율을 높이고, Epo-3’-UTR 부위를 사용하여 전사 후 mRNA를 안정화 시킴으로써 효과적인 유전자 발현을 유도 하였다. 일반적으로 사용되는 SV promoter에 Epo enhancer가 결합된 pEpoSV-Luc, Epo-3’-UTR이 결합된 pSV-Luc-UTR, Epo enhancer와 Epo-3’-UTR이 모두 결합된 pEpoSV-Luc-UTR 세
가지 벡터 시스템을 제작하였다. Reporter gene으로 luciferase 를 사용하여 벡터의 정상산소 조건과 저산소 조건에서의 유전자 발현량을 확인하였다. 세 개의 벡터 모두 정상산소 조건에 비해 저산소 조건에서 4~18.5배 발현량이 증가하였 고, pEpoSV-Luc 벡터의 발현량이 가장 높았다. Epo enhancer 나 Epo-3’-UTR이 없는 pSV-Luc에 비해 Epo enhancer가 있는 경우 205배, Epo-3’-UTR이 있는 경우 30배, 둘 다 포함한 경 우 116배 증가로 pEpoSV-Luc 벡터의 발현량이 가장 높았다.
하지만 정상산소 조건에 비해 저산소 환경에서 증가한 유 전자 발현 비율은 Epo enhancer와 Epo-3’-UTR이 모두 있는 벡터가 약 20배로 가장 많이 증가하였다. 최종 발현량이 가 장 높은 pEpoSV-Luc 벡터의 luciferase 유전자를 혈관생성 기 능을 하는 VEGF 치료유전자로 대체한 벡터 시스템 pEpoSV- VEGF를 제작한 후 mNSCs에 도입하였을 때 역시 저산소 조 건에서 유전자 발현량이 25.5배 증가하였다.
VEGF는 혈관생성을 통해 생체 내에 산소를 공급함으로 써 손상 조직에 대한 치료유전자로 알려져 있다[17-19]. 혈 관생성뿐만 아니라 세포증식 및 축삭성장, 신경 및 비신경 세포의 생존 등 저산소 조직에서 신경보호 기능도 알려져 있다[17,18,20-22]. 저산소 조직에 혈관을 생성함으로써 산 소 공급을 유도 하지만 부작용으로 과발현될 경우 종양이 생길 가능성이 있다. 저산소 유도 유전자 발현 시스템을 이 용하면 저산소증에 의해 조직이 저산소 환경이 되었을 때, 치료 유전자 발현을 효과적으로 증가시키고, 손상 조직이 회복됨에 따라 유전자 발현이 줄어 치료유전자의 과발현에 의한 부작용을 줄일 수 있다. 저산소증에 의해 손상된 신경 조직을 회복하기 위해 사용할 수 있는 치료 유전자에는 VEGF 외에 뇌유래신경영양인자(Brain-derived neurotrophic factor, BDNF), 글리아세포-유래 신경영양인자(glial cell line- derived neurotrophic factor, GDNF), 과립구 대식세포 콜로니 자극 인자 (Granulocyte macrophage colony-stimulating factor, GM-CSF), 신경 성장 자극 요소(Neurotrophin-3, NT-3)가 있 다.
저산소에서 치료 유전자를 발현시킴과 동시에 신경 조직 에 특이적으로 유전자를 발현시킨다면 저산소증에서 손상 받은 신경조직을 보다 효과적으로 목표로 할 수 있다. 신경 조직에서 유전자 발현을 유도하기 위해 신경 특이적 promoter 를 사용할 수 있다. 시냅신 I (synapsin I), 칼슘/칼모둘린 의존 인산전달효(calcium/calmodulin-dependent protein kinase II;
CaMKII), 튜불린 알파 I (tubulin alpha I), 혈소판 유래 증식인 자 베타 체인 (platelet-derived growth factor beta chain) pro- moters가 대표적으로 알려진 신경 promoter이다. 여러 종류 의 신경 promoter 중 뉴런특이적에놀라아제(neuron specific enolase; NSE)는 neuroendocrine에서 발현하는 glycolytic protein 이다. NSE promoter는 신경뿐만 아니라 신경계열 세포에서 도 발현이 증가하기 때문에 산소조건에 취약한 신경조직에
적합한 후보물질이다[14,23-25]. 저산소에서 가장 많은 발현 을 보인 Epo enhancer와 SV promoter 대신 신경 특이적 promoter를 결합한다면 저산소 환경의 신경조직에서 이중 특이적으로 유전자를 발현시킬 수 있다.
유전자의 발현은 전사, 전사 후 외에 번역 후 조절이 가능 하다. HIF-1은 DNA binding, oxygen dependent degradation (ODD), transactivation 세 가지 domain으로 구성되어 있다[8].
정상 산도 농도에서는 활성화된 프롤린 하이드록실레이즈 (proline hydroxylases; PHDs)이 ODD domain을 하이드록실화 시킨다. 하이드록실화된 ODD domain은 폴리유비퀴티네이 션에 의해 분해된다. 저산소 환경에서는 PHDs가 활성화되 지 않아 HIF-1이 분해되지 않고 축적된다. HIF-1의 ODD domain에 의해 산소 농도에 따라 발현된 단백질이 분해 또 는 유지 되는 것을 번역 후 조절이라고 한다. ODD domain을 목적 유전자와 융합시키면 정상산소 조건에서는 발현된 단 백질이 분해되는 성질을 이용하여 저산소 손상 조직에서는 치료유전자를 발현시키고 손상 조직이 회복됨에 따라 유전 자 발현을 낮출 수 있다.
산소 농도에 대한 신경 손상의 대표적인 질환으로는 허 혈성 뇌졸중(ischemic stroke)이 있다. 뇌졸중은 다양한 원인 에 의해 뇌혈관에 폐색이 발생하여 뇌에 공급되는 혈액량 이 감소하는 현상이다. 뇌혈류 감소에 의해 산소 공급이 줄 어드는 현상이 일정 시간 이상 지속되면 뇌조직의 괴사가 시작되는데 이를 뇌경색(cerebral infarction)이라고 한다. 손상 조직에 산소를 공급할 수 있는 혈관을 생성함으로써 회복 을 유도할 수 있다. Epo enhancer나 Epo-3’-UTR을 이용하여 치료 유전자의 발현을 증가시킴으로써 효과적인 혈관생성 유도 및 과발현에 의한 부작용을 줄일 수 있다.
세포 또는 조직으로 유전자를 전달하는 PEI는 비바이러 스성 유전자 전달 매개체 중 하나로 양이온성 중합체 (cationic polymer)이다. Branched PEI는 질소 원자가 branch마 다 포함되어 높은 양전하를 띄게 되며 완충 작용을 통해 다양한 pH에서 안정적이기 때문에 대표적으로 사용되는 비바이러스성 유전자 전달 물질이다. 양전하의 PEI와 음전 하의 DNA가 결합하여 polyplex를 형성하고 Proton-sponge effect에 의해 세포질로 유전자가 전달된다[26]. Lipid, nanosome 등 다양하게 개발되고 있는 비바이러스성 유전자 전달 물 질 중 비교적 독성이 적고 전달 효율이 효과적이라고 알려 져 있으며 polyplex의 크기, 전하, N/P ratio에 따라 유전자 전달 효율 및 독성이 달라질 수 있다.
V. 결 론
저산소에서 유전자 발현을 유도하기 위해 Epo enhancer와 Epo-3’-UTR 부위를 이용하면 유전자의 전사와 전사 후 단계
를 조절할 수 있다. Epo enhancer 그리고/또는 Epo-3’-UTR이 재조합 된 벡터를 mNSCs에 도입하였을 때, 정상산소 조건 보다 저산소 조건에서 최대 18.5배까지 유전자 발현이 증가 하였고, Epo enhancer나 Epo-3’-UTR이 없을 때보다 최대 200 배 발현이 증가하였다. 이 유전자 발현 벡터 시스템을 이용 하면 저산소증 조직에서 치료 유전자를 효율적으로 발현을 증가시킬 수 있다. 정상산소 조건에서는 발현량이 낮고, 저 산소 환경에서 증가한다는 점을 이용하면 손상 받은 조직 이 회복되었을 때 유전자 과발현에 의한 부작용을 피할 수 있다.
REFERENCES
1. Basnyat B. The physiologic basis of high altitude diseases. Ann Intern Med 2005;142(7):591; author reply 592.
2. Dillard TA and Ewald FW, Jr. The use of pulmonary function testing in piloting, air travel, mountain climbing, and diving. Clin Chest Med 2001;22(4):795 816, x.
3. Mortazavi A, Eisenberg MJ, Langleben D, Ernst P and Schiff RL. Altitude related hypoxia: risk assessment and management for passengers on commerical aircraft. Aviat Space Environ Med 2003;
74(9):922 927.
4. Kim HA, Mahato RI and Lee M. Hypoxia specific gene expression for ischemic disease gene therapy. Adv Drug Deliv Rev 2009;61(7 8):
614 622.
5. Semenza GL, Roth PH, Fang HM and Wang GL. Transcrip tional regulation of genes encoding glycolytic enzymes by hypoxia inducible factor 1. J Biol Chem 1994;269(38):23757 23763.
6. Zhu H and Bunn HF. Oxygen sensing and signaling: impact on the regulation of physiologically important genes. Respir Physiol 1999;
115(2):239 247.
7. Jiang BH, Rue E, Wang GL, Roe R and Semenza GL.
Dimerization, DNA binding, and transactivation properties of hypoxia inducible factor 1. J Biol Chem 1996;271(30):17771 17778.
8. Huang LE, Gu J, Schau M and Bunn HF. Regulation of hypoxia inducible factor 1alpha is mediated by an O2 dependent degradation domain via the ubiquitin proteasome pathway. Proc Natl Acad Sci U S A 1998;95(14):7987 7992.
9. Jiang BH, Semenza GL, Bauer C and Marti HH. Hypoxia inducible factor 1 levels vary exponentially over a physiologically relevant range of O2 tension. Am J Physiol 1996;271(4 Pt 1):C1172 1180.
10. Jiang BH, Zheng JZ, Leung SW, Roe R and Semenza GL.
Transactivation and inhibitory domains of hypoxia inducible factor 1alpha. Modulation of transcriptional activity by oxygen tension. J Biol Chem 1997;272(31):19253 19260.
11. Iyer NV, Kotch LE, Agani F, Leung SW, Laughner E, Wenger RH, et al. Cellular and developmental control of O2 homeostasis by hypoxia inducible factor 1 alpha. Genes Dev 1998;12(2):149 162.
12. Ambrosini G, Nath AK, Sierra-Honigmann MR and Flores-
Riveros J. Transcriptional activation of the human leptin gene in response to hypoxia. Involvement of hypoxia inducible factor 1. J Biol Chem 2002;277(37):34601 34609.
13. Scandurro AB and Beckman BS. Common proteins bind mRNAs encoding erythropoietin, tyrosine hydroxylase, and vascular endothelial growth factor. Biochem Biophys Res Commun 1998;246(2):436 440.
14. Gorlach A, Camenisch G, Kvietikova I, Vogt L, Wenger RH and Gassmann M. Efficient translation of mouse hypoxia inducible factor 1alpha under normoxic and hypoxic conditions. Biochim Biophys Acta 2000;1493(1 2):125 134.
15. Rondon IJ, MacMillan LA, Beckman BS, Goldberg MA, Schneider T, Bunn HF, et al. Hypoxia up regulates the activity of a novel erythropoietin mRNA binding protein. J Biol Chem 1991;
266(25):16594 16598.
16. McGary EC, Rondon IJ and Beckman BS. Post transcriptional regulation of erythropoietin mRNA stability by erythropoietin mRNA binding protein. J Biol Chem 1997;272(13):8628 8634.
17. Brockington A, Lewis C, Wharton S and Shaw PJ. Vascular endothelial growth factor and the nervous system. Neuropathol Appl Neurobiol 2004;30(5):427 446.
18. Silverman WF, Krum JM, Mani N and Rosenstein JM.
Vascular, glial and neuronal effects of vascular endothelial growth factor in mesencephalic explant cultures. Neuroscience 1999;90(4):1529 1541.
19. Lee RJ, Springer ML, Blanco-Bose WE, Shaw R, Ursell PC and Blau HM. VEGF gene delivery to myocardium: deleterious effects of unregulated expression. Circulation 2000;102(8):898 901.
20. Lu K, Liang CL, Chen HJ, Chen SD, Hsu HC, Liliang PC, et al. Injury severity and cell death mechanisms: effects of concomitant hypovolemic hypotension on spinal cord ischemia reperfusion in rats. Exp Neurol 2004;185(1):120 132.
21. Park E, Velumian AA and Fehlings MG. The role of excitotoxicity in secondary mechanisms of spinal cord injury: a review with an emphasis on the implications for white matter degeneration. J Neurotrauma 2004;
21(6):754 774.
22. Blesch A and Tuszynski MH. Spinal cord injury: plasticity, regene ration and the challenge of translational drug development. Trends Neurosci 2009;32(1):41 47.
23. Hioki H, Kameda H, Nakamura H, Okunomiya T, Ohira K, Nakamura K, et al. Efficient gene transduction of neurons by lentivirus with enhanced neuron specific promoters. Gene Ther 2007;14(11):872 882.
24. Navarro V, Millecamps S, Geoffroy MC, Robert JJ, Valin A, Mallet J, et al. Efficient gene transfer and long term expression in neurons using a recombinant adenovirus with a neuron specific promoter.
Gene Ther 1999;6(11):1884 1892.
25. Forss-Petter S, Danielson PE, Catsicas S, Battenberg E, Price J, Nerenberg M, et al. Transgenic mice expressing beta galactosidase in mature neurons under neuron specific enolase promoter control. Neuron 1990;5(2):187 197.
26. Mahato RI, Lee M, Han S, Maheshwari A and Kim SW.
Intratumoral delivery of p2CMVmIL 12 using water soluble lipopolymers.
Mol Ther 2001;4(2):130 138.