Kuχ J Appl. Micorbiol. Bioeng.
Vol.‘ 16, No. 1, 38-45 (1 98에
Intrasporangium속 방선 균의
3-Deoxy-D-Árabino-Heptulosona te 7-Phosphate Synthase Regulation조원대 1 ,최용진 2 ,양한철 1*
고려대학교 식품공학과 2 고려대학교 유전공학과
Regulation of 3-Deoxy-D .. Arabino-Heptulosonate 7-Phosphate Synthase in Intrasporangium sp.
Cho, ’Won-Dai\ Yong-Jin Choi\ and Han-Chul Yangl *
lDepartment 01 Food Technology
2Department 01 Genetic Engineering, College 01 Agriculture, Korea University, SeouI136-701, Korea
Studies were made on the regulation of 3-deoxy .. D .. arabino .. heptulosonate 7 .. phosphate synthase (DAHP synthase) of a species of Intrasporangium. The formation of the enzyme was increased dur ..
ing exponential phase reaching its maximum value(approximately 19 units/mg) at the early sta ..
tionary phase of the cell growth. Among the three aromatic amino acids and the intermediary meta ..
bolites of the biosynthetic pathway tested as potential inhibitors of DAHP synthase activ,ity. Only tryptophan caused inhlbition showing about 33070 inhibition of the partially.purified enzyme activity at O.lmM tryptophan. The inhibition was strongly dependent on the pH and maximum at pH 6.0.
Tryptophan inhibited the enzyme activity noncompetivively with respect to both phosphoenol ..
pyruvate and erythrose 4 .. phosphate. Tryptophan hydroxamate and 6 .. fluoro .. DL-tryptophan, anal- ogues of tryptophan, were found to be stronger inhibitors than tryptophan. Synthesis of DAHP syn- thase was also subject to feedback repression controI. Only tryptophan caused feedback repression and the degree of repression was 43% at the presence of 10mM tryptophan in the culture medium.
DAHP synthase of a 6-fluoro-tryptophan resistant mutant of Intrasporangium sp., which ac- cumulated highly elevated level of L-phenylalanine in the cuIture medium, showed no altered sen- sitivity to tryptophan inhibition, but the enzyme production was derepressed. The enzyme activity of the mutant strain was about 6 times higher than that of the parent strain. Reg띠ation features of the DAHP synthase described above implied that in Intrasporangium sp. the first step in the common aromatic synthetic pathway was catalized‘by one single tryptophan inhibitable and repressable DAHP synthase.
분지를 이 루고 있는 ·생 합성 대 사과정 (branched biosynthetic pathway) 은 대부분의 경우 각 분지 대 사경로 (branched pathwâys) 의 최종 대사산물이 공 통 대사과정 (common pathway) 의 첫 번째 반응을 촉매하는 효소를 여러가지 서로 다른 복잡한 방식에 의해 feedback control을 함3로써 전체 대사과정을 적절히 조절하고 있는 것이 일반적이다.
대다수의 미생울과 식울체에서 phenylalanine,
tyrosíne 빚 tryptophan 등의 방향족 아미 노산과 p-amínobenzoíc acid 빚 p-hydroxybenzoic acid 등의 몇가지 방향족 화합물은 동일한 공통 대사경로와 각 각의 고유분지 경로를- 거쳐 생합성되고 있우며 이 대사경로에 대해서는 지금까지 많은 연구가 이루어 져 그 대사 조절기구가 비교적 잘 밝혀진 대사과정 중의 하나라고 하겠다 (1-4)
특히 방향족 아미노산 생합성 대사과정중 공통 경
Key words: Enzyrne regulation, 3-dehydroxy-D-arabinose-Heptulosonate-7-Phosphate synthase, Intrasporangium sp.
'Corresponding author
38
Vo1.16, No.l 39
로의 첫 번째 반응, 즉
erythrose4-phosphate 와
D- er'ythrose 4- phosphate (sod. salt) , phosphoen- phosphoenolpyruvate 를 축 함, 5-deoxy-D-arabino-heptulosonic acid
7-phosphate(DAHP) 를 생 성 하 는
반응은 DAHP synthase (E.C. 4. l. 2. 15) 에 의 해 촉 매되며 이 효소는 생물체의 종류에 따라 lsoenzyme의 구성이 다를 뿐만 아니라 다양하고 복잡한
feedbackinhibition과
feedbackrepression을 받음 o
로써 이 반응은 최종 산울인 방향족 아미노산 생산 량에 매우 중요한 영향을 마치는 주요 대사조절 단·계가 되고있다 (1 , 5). 예를 들연 E. coli (10, 11), s.
typhimμy낌m
(12, 13)벚 Neuγospora
crassa (14, 15)등의 균종에서는 방향족 아미노산 각각에 의해
feedback
control을 받는 3종의 isoenzyme이 존재 하 며
Sac. cerevisiae (16)및
Schizosac. pombe (17)등
의 효모에서는 2종의 isoenzyme 이 확인되고 있고 Bacillμs sμbtilis (18, 19), Thiobacillus neopolitaηus(20), Pseudomonas aerugiηosa (7, 21) 와 수 종 의 Streptomyces속 방선균 등 (22, 23) 은 저해양식은 서
로 다르나 한 종류의 단일 단백질의
DAHPsynthase를 가지고 있다고 보고되고 있다.
한편 Jensen 등 (6, 7) 은 약 91종의 미생물을 대상 A로 하여 DAHP 축합반응의 대사조절기구를 비교 연 구 한 결 과 균종 에 따 라 isoenzyme feedback inhibition, cumulative feedback inhibition 맞
sequential feedback inhibition 등의 서 로 다른 다양 한 저해양식을 나타내고 있다고 보고하고 있으며 또
Shiio 등 (8)
에 의 하연
Brevibacteriumflavz‘m에 서 는
conserted feedback inhibition에 의 해 DAHP 합성 이 조절되고 있다고 한다.저자 등은 현재 각종 항생울질 생산 등 산업적으 로 매우 유용하게 이용되고 있는 방선균을 이용, 방 향족 아0] 노산의 대량생산 가능성을 타진하고저 우 선 L-phenylalanine 생산능이 있는 방선균을 토양으
로부터 분리, 분리균의 동정과
L-phenylalanine생
산 조건을 조사, 이마 보고한 바 있으며 (9) 본 연구 에서 는 lntrasporaηgzum속의 균주로 동정된 토양분 리 방선균의 DAHP synthase 대사조절 특성을 조사,그결과를보고한다.
재료 및 방법
사용균주 및 시약
본 연구에 사용한 균주는 Iηtrasporangiμm속의 방
선균.9...로 동정된 (9) 토양 분리율이며 사용 시약중olpyruvate(monocyclohexyl ammonium salt), bo- vme serum albumin 빛 c1ithiothre:‘01 등은 Sigma
제품, 기타 일반시약은 시판 일급 이상의 분석용 시
약을 사용하였다.배지 및 배양조건
효소생산을 위 한 균체증식 배지 는
buffer 56 (24)에 탄소원A로
glucose (10g/ll 를 첨가한 최소배지를
사용하였으며 동일 배지에서 48시간 전배양한 균체 를 종균으로 하여 2%정도 접종, 30.C 에서 48시간 배양하여 얻은 균체를 효소원으로 이용하였다.조효소액 조제
48시간 진탕배양한 배양액을 4.C 에서 10분 (3, 000
rpm) 간 원심분리하여 균체를 분리,
O. 1 M soclium phosphate buffer(pH 7. 0)로 2회 세 척 하였다.세척한 균체는
0.1 mMCoCI
2가 함유된
0.1M soclium phosphate buffer(pH 7.이 에 현 탁 시 킨 후
sonicator(Karl kolb 200-w) 를 이용, 4.C 에서 maxl- mumpower로 세포를 i배한 다음
4.C 12,000 rpm에서 20분간 원심분리하여 얻은 상등액을 조효소액
a로 사용하였다.DAHP synthase의 활성 측정
DAHP synthase의 활성 측정은 Gollul;J 등의 방법
(25) 에 따라 40 μ mole potassium phOsphate buf- fer(pH 7.
이
, 1. 0μ
mole phosphoenolpyruvate및
O.5
μmole
erythrose4-phosphate를 함유한 반응액
O.6ml를 3TC 에서 5분간 예온한 다음 효소액
0.4ml를 첨가, 정확히 10분간 반응시키고 10% trichlor- oacetic acid
용액 0.2ml를 가하여 반응을 정 지 시 켰 다. 반응을 정지시킨 반응액은 얼음울에서 냉각된 후 원심분리하여 얻은 단백질이 제거된 상등액
0.8ml를 취 해
0.2 M sodium metaperiodate(9 M H3PO‘용액) 용액
0.1ml를 가하여 3TC 에서 5분간 반응시 켰 다. 상기 반응액 에
0.8 M soclium arsenite용액
(0.5M
HCI 송 역)
0.5ml를 가하고 5분간 방치한 후
0.6% thiobarbituric acid용액 3.0ml를 가해 끓는 물에서 15분간 반응, 냉각시키고
cyclohexanone 4. 0ml을 가하고 약 10초간 흔화하여 색소를 축출한 후
원심분리
(3000 rpm,10분)하여 얻은 상등액을
549nm에서 흡광도를 측정, 생성된
DAHP량을 산출하
였 다. 이 때 사 용 한 DAHP 의 molar extinction40
coefficient는
68,000이 었 다.
단백질 정량
단백질은
Lowry등의 방법
(26)에 따라
bovine serumalbumin을 표준단백질로 사용하여 750nm에 서 흡광도를 측정, 단백질 량을 산출하였다.
비활성측정
DAHP synthase 1 urùt는 3TC, 1 분간 반응으로 0.1 μmole의 DAHP를 생산할 수 있는 효소량으로
정의하였으며 비활성은 효소단백질 mg당 활성
unit (U) 로 표시하였다.효소정제
모든 정제 조작은 4.C 에서 행하였으며 사용 완충
용액은
0.1 mMCoC1
2를 함유한
O. 05 M potassium phosphate buffer(pH 7.0)이 었 다.
조 효 소 액 에
ammonium sulfate플 첨 가 하 여
30-50%포화농도에서의 단백침전물을 분리,
potas, sium phosphate buffer에 용해 시 킨 후 약 6시 간 동안 동일 완충용액중에서 투석시켰다. 투석이 끝난
효소용액은 ml당 20mg의 단백질 농도가 되도록 희석 하고
acetone분획 을 실시 하여
45-55% acetone분획의 단백질 침천을 완충용액£로 용해, 투석과정 을 거쳐 얻은 부분정제 효소를 효소활성 저해 실험 에 사용하였다.
6-Fluoro-DL -tryptophan 내성변이주의 분리 대수증식 중기의 율체를 분리, 세척한 후 tns- maleic acid buffer에 현 탁시 키 고 500 μ g/ml 농도의 N-methyl-N'-nitro-N-nitrosoguanidine (MNNG) 을
첨가, 30.C 에서 1시간 처리하였다.
MNNG처리한 세포는 셰척 후 5X lO-',M의 6-fluoro-tryptophan을 첨가한 기본배지에 도말, 30.C 에서 6일간 배양하여 plate 상에 자란 내성변이주를 일차 분리하고 2차로
cross-feeding시험과 직접 배양시험을 거쳐 분리,
내성균 중에서
L-phenylalanine생산능이 높은 변 01 주를 최종 선별하였다.
결과 및 고찰
배양시간과 DAHP synthase 세포내 함량
E.
coli 를 비 롯한 대 다수의 미 생 물
DAHP synthase는 애우 불안정한 효소로서 일정 배양시간Kor. J Appl. Mic:ηibio!. Bioeng.
이 경과하면 세포내 함량이 급격히 감소되는 경향을
나타낸다고 보고하고 있다(1,
5, 7).따라서 본 연구
의 시험균인 Iηlrasporaηgium 속 분리 방선렌을 “재료 빛 방법”란에 서솔한 배양조건하에서 진탕배양
하면서 배양시간에 따른 균체증식속도, DAHP synthase의 세포내 함량 등을 조사, Fig.1과 같은 결과를 얻었다. DAHP synthase의 활성은 균체증식 초기 정체기에서 최고치를 나타냈으며 그 이후 부터는 급격한 활성감소를 보여주고 있어 본 균주의
DAHP synthase역시 셰포내에서 극히 불안정하다 는 것을 알 수 있었다. 따라서 조효소액을 얻기 위 한 이후 실험에서는 48시잔 배양한 균체를 효소원으
로 사용하였으며 DAHP synthase의 세포내 안정 화 문제는 본 시험균에 의한 방향족 아미노산 생산에 중요한 영향을 미칠 것으로 사료된다.DAHP synthase의 정제
초기 정체기의 균체로부터 얻은 조효소액을 Table 1에 표시되 어 있는 바와 같이 ammonium sulfate 분 획과 acetone 분획 과정을 실시하여 약 8.6배 정제 된 부분 정제효소을 얻을 수 있었다• E. coli (27, 28) , S. lyphimuriμ m(29) , B. subtilis(l9, 30), Brevibacterium flavum (31) 빛 Streþlomyces aμ r,
eofacieηs(23) 등의 다수 균종에서 는 각종 DAHP synthase
isoenzyme을 균일 단백질까지 정제하였다
고 보고하고 있으나 본 균의 DAHP synthase는 m겨
\읍E
효 3 3Q j 6
gζ
2 홍 L 공
-효.t잉여-‘‘、
2α합1
1 1융
1212 36 60 84
Culture time(hrs.)
Fig.1. Variation in DAHP synthase specific activity and cell growth during culture of Intrasporangium sp.
〔농lls were grown at 300 C in 10Omlof the ba싫 medium with reciprocal sh와cing. 0, specific activity; ., cell growth; 6., pH
Vc,z. 16, No. 1 41
Table 1. Purification of DAHP synthase of
“
trasporan- 과 혼합하여 첨 가했을 때 현저 한 활성 저 해 효과를 glUm sp.Total Total Specific Fold/
Step Fraction activity protein activity Yield purifi- (units) (mg) (U /mg) (%) cation Crude
extract 49,000 613 80 100 1 II (NH4)2S04 35,000 264 134 72 1.7
30-50%
III Acetone 16,500 24 687 34 8.6 45-55%
vzvo에서 뿐만 아니라
invitro에서도 매우 불안정함
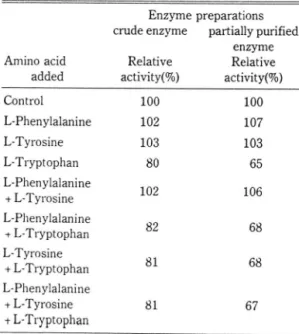
으로 어떤 특별한 안정화 방안을 강구하지 않는 한 더 이상의 정제가 어려울 것으로 생각된다.최종 대사산물에 의한 DAHP synthase
활성
저해주요 최종 대사산울인
phenylalanine, tyrosine벚 tryptophan을 만독, 2종 흑은 3종을 흔합, 최종 농 도가
l mM되게 효소반응액에 첨가하여 이들 아미 노산의
DAHP synthase 효소활성저해효과를 살펴본 결과 (Table
2)조효소와 부분 정제효소 오두
phenylalanine과tyrosme은
전혀 저 해효과를 나타내 지 않았으나 tryptophan은 만독 또는 다른 아미노산 Table 2. Inhibition of DAHP synthase activity by aromatic amino acids.Enzyme preparations crude enzyme partially purified
enzyme Amino acid Relative Relative added activity(%) activity(%)
Control 100 100
L. Phenylalanine 102 107
L-Tyrosine 103 103
L-Tryptophan 80 65
L-Phenylalanine
102 106
+ L-Tyrosine L-Phenylalanine
82 68
+ L-Tryptophan L-Tyrosine
81 68
+ L-Tryptophan L-Phenylalanine
+ L-Tyrosine 81 67
十L-Tryptophan
보였다. 따라서 본
효소활성은
tryptophan에 의해 서만특이적으로 활성이 저해되는 것으로 생각된다.
1 mM tryptophan에 의 한 저 해 율은 조효소가 약 20%,
부분정제 효소가 약
35%정도로서 저해농도
에는차이가 있으나 각 아미노산에 대한 전반적인
저해 양식에는 조효소와 부분정제 효소가 갇은 경향을 나타냈다. 이와 같은 결과는 본 시험균의
DAHP synthase가 한 종류의 단백질, 즉 tryptophan에 의해서만 저해를 받는 단일효소로 구성되어 있다는 것
을 암시해 주고 있다.또한
tryptophan첨 가 농도에 따른 저 해율의 차이
를 조사해 본 결과 농도가 증가할수록 저해율도 바례적
으로높아져
1 X 10-2 M tryptophan을첨 가했을
때약 60% 의 높은 저해율을 나타냈다 (Table
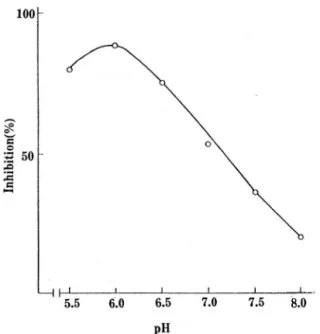
3).Tryptophan의 저해효과는 반응액의 pH에 따라서 도 큰 차이를 나타내
pH6.0부근의 약산성에서는 가장
높은저해를 나타낸 반면 알카라성이 될수록 저해효과가 현저히 강소됨을 보이고 있다 (Fig.2).
그러 고 tryptophan에 의 한
DAHP synthase활성 저 해 는 두
기질 인 phosphoenolpyruvate와
erythrose 4-phosphate에 대해 다 같이 비경쟁켜 저해 양식을따르고
있어
(Fig. 3, 4) Neμrospora crassa DAHPsynthase (14) 와 함께 특이한 저해 양식을 나타내고
있다.
한
편각
종 tryptophan analogue 의 DAHP synthase 활성에마 치
는 효과를 상펴본 결과
Table4에 표시되어 있는 바와
같이 L-tryptophanhydrox -amate가
92%, 6-fluoro-DL-tryptopl-.an이 73% 의 저Table 3. Inhibition of DAHP synthase activity by vari- ous concentrations of L-tryptophan.
Concentration
oítryptophan(~) Inhibition (%)
No addition 0
1 X 10-4 33
1 X 10-3 39
5 X 10-3 57
1 X 10-2 59
The reaction mixture contained enzyme of llunits, 0_5μmole erythrose 4-phosphate, 1.0f..l mole phospho- The amino ~cids were added at a final concentration of enolpyruvate and tryptophan added a한 indicated in a 1 X 10-3~ into the standard reaction mixture detailed in total volume of 1m! of
O.04~
potassium phosphate .buf~aterials and Methods. fer (pH 7.0)
42
100
%
( 옹 ) t
。
•
‘ ·a
i s
I
I
。
5.5 6.0 6.5 7.0 7.5 8.0
pH
Fig.2. Inhibition of DAHP synthase activity by trypto.
phan 잃 a function of pH.
The reaction mixture contained 11 units of enzyme, 0.5 μ
mole erythrose 4-phosphate and 1.0 μ mole pho- sphoenolpyruvate in 1 ml of 0.04M phosphate buffer at various pH values and in the presence of 1 x 10-4M tryp- tophan.
0.4
0.3
~
>l
0.2
10 20
lIPEP (mM-l)
Fig. 3. Double reciprocal plots of initial velocity of DAHP synthase as a function of substrate concentration.
The reaction mixture contained 11 units of enzyme, 0.5 μ
mole erythrose 4-phosphate, various concentrations of tryptophan and phosphoenolpyruvate as indicated in a total volume lml of 0.04M potassium phosphate buffer (pH 7.0) .
.6.,3 x 10-3M; .,3 x 10-4M; 0, no addition tryptophan
Kor. f. Appl. Microbiol. Bioeηg
0.4
0.3
;>
.... 0.2ι-
10 20 l/E4P (mM-l)
Fig. 4. Double reciprocal plots of initial velocity of DAHP synthase as a function of substrate concentration.
The r-eaction mixture contained 11 units of enzyme, 1.0μ
mole phosphoenolpyruvate, various concentrations of tryptophan and erythrose 4-phosphate as indicated in a total volume lml of 0.04M potassium phosphate buffer (pH 7.0) .
.6., 3 x 10-3M; ., 3 x 10-4M; 0, no addition tryptophan
해율을 보임으로써 일반적 A로 tryptophan.'봐다 훨씬
높은 저해효과를 보이고 있는 반면에 같은 불소 유
도체 이 면서도 5-fluoro-DL-tryptophan은 전혀 저 해 효과를 나타내지 않는 특이한 현상을 보였으나 그원인은 더 이상 조사하지 않았다.
다음은
각종 방향족 화합물의 공통의 최종 중간 대사울인
chorismicacid와 기타 주요 중간 대사울의
Table 4. Inhibition of DAHP synthase by tryptophan analogues.
Analogue No addition
Relative activity(%) 100
5-Methyl-DL-tryptophan 50
5-Fluoro-DL-tryptophan 100
6-Fluoro-DL-tryptophan 27
L-Tryptophan-hydroxamate 8
L-Tryptophan 52
The activity of enzyme was determined in the standard readion mixture with the addition of inhibitors at a con- centration of 2 x 10-3M. Compounds of the DL-form were added at a concentration of 4 x 1O-3M
은
E.coli플 비롯한 다른 많은 미생물 (1 ,
17, 32, 33)의 경 우와 같이
feedbackinhibition과 동시 에
feedback repression controI 을 받고 있 어 DAHPsynthase가 촉매 하는 공통대사 경로의 첫 번째 반응 이 방향족 아미노산 생산에 매우 중요한 대사조절단
계가되고 있다는 것을 말 수 있었다.
그 러 나 같 은 방 선 균 이 라도 Stγ-eptomyces
aur eofaciens (22) 는 최 종 대 사 산 물 에 의 해 DAHPsynthase
생산이 전혀 저해를 받지 않는다고 보고하
고 있어 본 시험균과는 다른 면을 보였다.
또 Fig.5에 표시 되 어 있는 바와 같이
tryptophan 의 첨가농도가 증가될수옥 이에 비례하여 균체증식도 뚜렷한 저해를 받고 있음을 알 수 있었다. 이와 같은 실헝 결과는 본 시 험 균의
DAHPsynthase는 전 항의
feedback inhibition연구에서 얻은 결과, 즉 본 시 험 균의
DAHP synthase는단일 단백 질로 구성 되
어 있을 것이라는 추정을 확인해 주는 중요한 자료가 된다고 하겠다. 따라서 tryptophan에 의한 생육 저해현상도 배지중
tryptophan첨 가로
DAHPsynthase 생산량의 현저한 강소를 초래하므로 같은
대사과정을 거쳐 생합성되고 있는 tryrosine 과
phenyialanine을 포항한 다른 방향족화합울 생산의 현저한 감소를 가져올 것으로 예상된다. 그 결과 이 들 화합울의 세포내 결펌현상이 유발되어 정상적인율체생육이 저해되기 때문이라고 생각된다.
Vol. 16, No_ 1 43
활성 저 해 효과를
조사해본 결과
(Table 5) shikimicacid에 의한 약간의 저해안을 인정할 수 있었으며 기
타 다른 중간대사울은 거의 저해효과를 나타내지 않 았다. 따라서 본 시험균의 DAHP synthase 는 sequential feedback inhibition 양식 에 의 해 활성 이조절되지 않고 있다는 것을 알
수있었다.
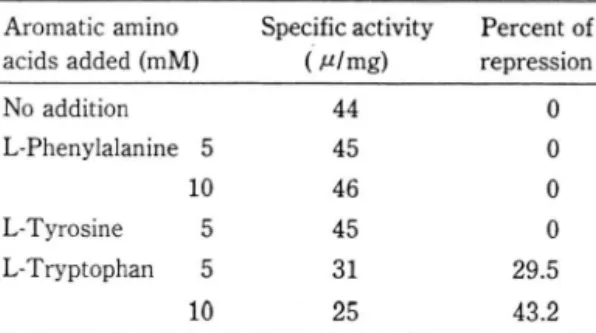
최종 대사산물에 의한 DAHP synthase 생합성 저
최소배지에 각 방향족 아미노산을 Table 6에 표시
되어 있는
농도호첨가, 시험균을 접종배양하여 최종
대사 산울에 의한 DAHP synthase 합성의 feedback repreSSlOn효과를 시 험 해 본 결과 L-phenylalanine과
L-tyrosine은 전혀 효소합성 저해효과를 나타내지 않 았으나 L-tryptophan을 첨가했을 때는 5mM 농도에 서 약 30%, lO mM에서는 약 43%의 효소생산을 저 해하였다.그러으로 본
시험균에 의한
DAHP synthase생산
Table 5. Inhibition of DAHP synthase activity by inter- mediary metabolites.
S{
X )EI
-∞∞ ‘여
·a·(} 0.3
The concentration of the intermediary metabolites tested was 1 x 10-3M and enzyme activities were deter- mined in the standard reaction mixture.
Inhibition (%) 4 0 0 0 6 6 5
A앙
A%
--
Shikimate Chorismate Prephenate Phenylpyruvate
p.Hyd미roxyphen따lylψpyl이ruvate Antπ띠t“hπran띠1니ilate
Indole
Intermediary metabolite
Table 6. Repression of synthesis of DAHP synthase.
Percent of represSlOn Specific activity
( μ Img)
Aromatic amino acids added (mM)
24 48
Culture time(hrs.)
Fig. 5. Effect of various tryptophan concentrations on cell growth of Inlrasporangium sp.
〔농llswere 밍UW!l at 300C in 100ml of the ba잃1 medium sup- plemented with various concentrations of tryptophan. 。’ I뼈le; .ò., 5mM;口, 10mM; -,2OmM;‘, f값nM; .,l(뻐nM;
72 O
O O O 29.5 43.2
μ%
%
뼈 쟁
입 있No addition L-Phenylalanine 5
10 5 5 10 L-Tyrosine L-Tryptophan
lntraspurangium sp. was cultured at 300C for 48 hours in the minimal medium supplemented with the aromatic amino acid as indicated in the table.
44
Tryptophan과 마찬가지 로 tryptophan analogue
특히 용fluoro-DL-tryptophan과
5-fluoro-tryptophan은 낮은 농도에서도 강력한 균체증식
억제효과를나
타내 었으며 tyrosine hydroxamate와 m-fluoro-tyr- osne등의
tyrosine analogue역시 높은 농도로 첨가 했을 때 뚜렷한 균체증식 저해효과를 보였다 (Table
7).Tryptophan analogue
중 에 서 전 혀
DAHP synthase 활성저해를 하지 않았던 5-fluoro-trypto-phan은 본 실험에서 현저한 생육저해효과를 나타내 는 특이한 현상이 관찰되었유나 이에 대해서도 더 이상 조사 연구하지 않았다.
한편 시 험균을 MNNG 처 리 하여 L-phenylalanine
생산능이 현저히 증가된 6-fluoro-DL-tryptophan 내 성 변이주를 분리, 분리변이주의
DAHPsynthase을 분석해 본 결과, 특히
FT411변이주의 경우
feedbackinbibition에 대한 감수성에는 뚜렷한 변화 가 없는 반면에
DAHP synthase 비활성이 친주에비 해 약 6배 증가된 값을 나타냈다
(Table 8).이 와 같은
FT411군주의
DAHP synthase비활성 증가 는
feedback repression해 체 와 관련된 어 떤 유전 적 변화에 기인된다고 생각된다.
Table 7. Effect of analogues of aromatic amino acid on ceIl growth of Intrasporangium sp.
Analogue
D L-5-Fl uoro-tryptophan 6-Fluoro-tryptophan 5-Methyl-DL-tryptophan L-Tryptophan hydroxamate p-Amino-DL-phenylalanine p-Fluoro-DL-phenylalanine DL-o-Fluoro-phenylalanine DL-Phenylalanine hydroxamate
β-2-Thienyl-DL-alanine
3-Amino-L-tyrosine m -Fl uoro-D L-tyrosine L-Tyrosine hydroxamate -; not tested
Relative growth rate{%) Conc. of analogue 5 x 10-4M 1 x 10-2M
78 58 80 99
100 100
100 92
100 78
100 71
100 59
100 120
100 35
100 32
The cells were cultured at 300C for 5 days in the mini- mal medium supplemented with the analogues shown in the table.
Kllr. J Appl. Micrllbi‘’1. Billeng
Table 8. Repression and inhibition of DAHP synthase of 6-f1uoro-DL-tryptophan resistant mutants of Intras- poranglllm sp.
Inhibition L-Phenylalanine Biomass Specific by 1mM produced Strain (mg/ml) activity trypto- (μg/m l)
(U Img) phan (%)
Parent 3.3 5.8 23 2.1
FT-108 2.4 17.3 21 33
FT-411 2.3 34.7 19 150
FT-554 2.8 15.2 25 96
The strains were cultured at 300C in the minimal medi- um. Specific activity and L-phenylalanine produced were determined in 2 days and 4 days of cultivation.
이상의 연구결과는 본 시험균을 이용한 방향족 아 마노산의 효율적 생산을 위해서는 우선 무엇보다도 DAHP synthase의 feedback inhibition과 feedback repression
control이 동시에 해제될
수있는 유전적 인 조치가 선결되어야 한다는 것을 나타내 주고 있 다.
요 약
토양 분리 방선균인 Intrasporangiuη2속의 DAHP
synthase는 세포증식의 초기 정체기에서 가장 높은
세포내 함량 (4. 2 units/ mg protein) 을 보였으나 그 이후 부터는 급격한 감소를 보임으로서 생체내에서매우 불안정한 효소임을 알 수 있었다.
한편 본 DAHP synthase는 최종대사산물 중에서
tryptophan에 의 해서 만 효소활성 저해를 받음으로서
Iηtrasporangiμm속의 DAHP synthase~는 단일단백질 로 구성되어 있다고 판단된다.Tryptophan의 저해는 pH6.0
부근에서 가장 높았
고 phosphoenol pyruvate와 erythrose 4-phosphate 두 기질에 대해 비경쟁적인 저해양식을 보였다.Tryptophan analogue 특 히 tryptophan hydrox
amate와 6-fluoro-DL-tryptophan은 tryptophan]효다
훨씬 높은 저해효과를 보였£나 chorismate를 비롯 한 기타 방향족 화합물 중간대사울은 뚜렷한 저해효 과를보아지 않았다.또 본 시 험 균을 10 mM tryptophan이 첨 가된 배 지 에 배 양했을 때 DAHP synthase 합성 이 약 43% 저 해됨과 동시에 생육속도 역시 현저히 감소되었다.
또한 L-phenylalanine 생산량이 친주에 비해 약 70
Vill. 16, No. 1 45
배 증 가 된 6-fluoro-DL-tryptophan 내 성 변 이 주,
FT-411은 DAHP synthase가 derepression된 변이주 로서 친주에 비해 약 6배 증가된 비활성을 나타냈 다.
17. Schweingruber, M.E. and H.B. Wyssling; Bio- chim. Bùφhys. Act,ι 350,319 (1974).
참고문헌
18. Jensen, R.A. and E.W. Nester;J Biol. Chem. 241, 3365 (1966).
19. Walter, M. Nakatsukasa and W. Nester; J B씨.
Chem. 247, 5972 (1972)
20. Kelly, D.P.; ArciI. Mikrobiol. 69,360 (1969).
1. Tribe, D.E., H. Camakaris, and ]. Pittard: J Bac- 21. Whitaker, R.J., M.]. Fiske and R.A. ]ensen; J feriol. 127, 1085 (1976). Biol. Chem. 257, 12789 (1982)
2. Umbarger, H.E.; Ann. Rel'. Biochem. 47, 533 22. Görisch, H. and F. Lingens; Biochim. Biophys. Ac-
(1978).α 242, 630 (1971).
3. Doy, C.H.: Rel'. Pure Appl. Chem. 18,41 (1968). 23. Görisch, H. and F. Li디gens; Biochim. Bùφhys. Ac- 4. Gibson, F.; Bacteriol. Reν. 32,465 (1968). fa. 242,617 (1971)
5. Choi, Y.].; ph. D. Thesis, University of New South 24. Monad, J., G. Cohen-Bazire and M. Cohen; Bψ-
Wales (1981). chim. Biophys. Acfa. 17,585 (1951).
6. ]ensen, R.A., D.S. Nasser, and E.W. Nestet; J 25. Gollub, E., K‘P. Liu K.P. and D.B. Sprinson; J
Baclenol. 94, 1582 (1967). Bacferiol. 115, 121 (1973).
7. ]ensen, R.A., D.H. Calhoun, and S.L. Stenmark; 26. Lowry O.H., ]. Rosebrough, A.L. Farr and R.].
Biochim. Bùψhys. Acta. 293, 256 (1973). Randall; J Biol. Chem. 193, 264 (1951).
8. Shiio, 1., S. Sugimoto, and R. Miyajima; J Bψ27. Schoner, R. and K.M. Herrmann; J Biol. Chem.
chem. 7!i, 987 (1974). 251, 5440 (1976).
9. Lee, S.Y., W.D. Cho, Y.]. Choi, and H.C. Yang; 28. Simpson, R.J., B.E. Davidson, T.A.A.,Dopheide S.
고려대학교 농럼논집 27, (1987). Andrews and ]. Pittard; J Bacfeη':01. 107, 798 10. Brown, K.D.; Genetics 60, 31 (1968). (197l).
11. Gibson, F., ]. Pittard; Bac!eriν1. Rel'. 32, 465 29. Hu, C.Y. and D.B. Sprinson; J Bacfeη‘01. 129, 177
(1968). (1977)
12. Gollub, E., H. Zalkin, and D.B. Sprinson; J Bi씨 30. ]ensen, R.A. and E.W. Nester; J Biol. Chem. 241,
Chern. 242, 5323 (1967). 3373 (1966)‘
13. Sanderson, K.E.; Bac!eriol. Rel.'. 31, 354 (1967). 31. Sugimoto, S.1. and 1. Shiio; J Biochem. 87, 881 14. Colin, H. Doy.; Biochim. Biophys. Acla. 159, 352 (1980).
(1968). 32. Kim, S.W., H. Davidson, and ]. Pittard;J Bacteri-
15. ]ensen, R.H. and D.S. Nasser;J Bacteη01. 95, 188 01. 108, 400 (1971).
(1968). 33. Jensen, R.A. and E.W. Nester;]. Mol. Biol. 12,468
16. Lingens, F., W. Goebel and H. Uesseler ; Biochem. (1965).
Z. 346,357 (1966). (Received December 17, 1987)