34 책임저자:노주원, 210-340, 강릉시 대전동 290번지
한국과학기술연구원 강릉분원 Tel: 033-650-7102, Fax: 033-650-7499 E-mail: [email protected]
접수일:2009년 3월 3일, 게재승인일:2009년 3월 18일
Correspondence to:Chu Won Nho
Natural Products Research Center, Korea Institute of Science and Technology Gangneung Institute, 290, Daejeon-dong, Gangneung 210-340, Korea Tel: +82-33-650-7102, Fax: +82-33-650-7499
E-mail: [email protected]
Cancer Suppressing Activity of Aceriphyllum rossii Extract Exerted through Induction of G0/G1 Arrest and Apoptosis
in HCT116 Human Colorectal Cancer Cells
Kyungsu Kang, Eun Hye Jho, Eun Ha Lee and Chu Won Nho
Natural Products Research Center, Korea Institute of Science and Technology Gangneung Institute, Gangneung 210-340, Korea
Aceriphyllum rossii, a wild edible green is distributed in Gyeonggi and Gangwon Province of Korea.
In order to examine cancer suppressing activity of the ethanolic extract of the aerial parts of A. rossii, we measured the antiproliferative activity of the extract in HCT116 human colorectal adenocarcinoma cells by using MTT assay. The extract induced a dose- and time-dependent cell growth inhibition in HCT116 cells. We also examined cell cycle distribution and apoptosis induction after treatment of the extract to find out the cause of its anti-proliferation activity. We discovered that the extract induced cell cycle arrest at G0/G1 phase through flow cytometric DNA content analysis. We also found out that the extract induced apoptosis inference from peculiar biochemical changes: the increase of the sub G0/G1 content (%) and externalization of phosphatidylserine through flow cytometric analysis. In addition, the extract induced apoptotic morphological changes such as chromatin condensation and the formation of apoptotic bodies. In brief, A. rossii extract exerted antiproliferative activity through the induction of G0/G1 arrest and apoptosis in HCT116 cells. Therefore, the ethanolic extract of A. rossii may be served as a potential chemopreventive agent for prevention and treatment of human colorectal cancer. (Cancer Prev Res 14, 34-39, 2009)
Key Words: Aceriphyllum rossii, Apoptosis, Cancer, G0/G1 arrest, HCT116
INTRODUCTION
Cancer chemopreventive agents are known to inhibit, reverse or retard carcinogenesis, which is a complex and multi-step process. Most of cancer chemopreventive agents are dietary phytochemicals. Cancer chemopreventive agents are divided into two classes: blocking agents and suppressing agent. Cancer blocking agents block the initiation step through the enhancement of detoxification or DNA repair. On the other hand, cancer suppressing agents inhibits the later stages of tumorigenesis such as promotion and progression stages. In case of suppressing agents, cell cycle arrest and apoptosis are important molecular mechanisms because they can inhibit or
eliminate abnormal cells.1,2) Many plant-derived extracts possess potent cancer chemopreventive effect. Combination of phyto- chemicals and whole plant extract often exert a stronger cancer chemopreventive effect than isolated single compounds, which are called additive and synergetic effects. Therefore, it is mean- ingful to investigate the cancer blocking and suppressing effects of not only single phytochemical but also whole plant extract for the development of cancer chemopreventive agent.2∼4) Aceriphyllum rossii, a wild plant is distributed in Gyeonggi and Gangwon Province of Korea. It is a popular ornamental plant due to its unique appearance, its genus name and Korean name are originated from its shape of maple-like leaves. As a wild edible green, its leaves and stalks before flowering are used as Korean diet.5) It is known to have the various pharmacolo-
gical activities such as anti-bacterial, anti-oxidant, anti-comple- mentary, anti-cancer and ACAT (Acyl-CoA: cholesterol acyl- transferase) inhibition activities.6∼11) Especially it was reported that the anti-cancer effects of A. rossii against human promy- elocytic leukemia cells is due to cytotoxic triterpenes present in the plant.9,12)
In this report, in order to examine cancer suppressing activity of the ethanolic extract of the aerial parts of A. rossii, we measured the antiproliferative activity of the extract in HCT116 human colorectal adenocarcinoma cells. We also examined cell cycle distribution and apoptosis induction after treatment of the extract, which are important molecular targets of cancer suppression.
MATERIALS AND METHODS 1. Plant materials
A. rossii was collected in Pyeongchang, Gangwon Province of Korea in August, 2007. A voucher specimen (D-012) has been deposited at the KIST Gangneung Institute. The dried aerial parts (30 g) of A. rossii were refluxed with 95% ethanol (700 ml) for 3 h and this residue was evaporated in vacuo to yield the total extract (1.1 g). The extract was dissolved in DMSO (dimethyl sulfoxide) for cellular treatment.
2. Cell culture
HCT116 cells were obtained from the American Type Culture Collection (Rockville, MD, USA). HCT116 cells were maintained in Minimal Essential Medium (MEM/EBSS) supple- mented with 10% fetal bovine serum (FBS, Hyclone, Logan, UT, USA), 100 units/ml penicillin and 100μg/ml streptomy- cin. The cells were maintained in a humidified 95% air, 5%
CO2 atmosphere at 37°C.
3. MTT cell proliferation assay
HCT116 cells (5×103 cells per well) were plated onto 96-well plates, incubated at 37°C for 24 h, then the cells were treated with the various concentrations (0∼200μg/ml) of the extract for 24 and 48 h. Cell viability was measured by using Cell Counting Kit (CCK-8, Dojindo Laboratories, Tokyo, Japan) as previously described method.13) Cell survival (%) was calculated as the percentage of the absorbance at 450 nm of extract-treated cells and vehicle control cells. IC50 values were calculated from the equation of the logarithmic line determined
by fitting the best line (Microsoft Excel) to the concentration-cell survival (%) curves. The IC50 value was obtained from the equation for y=50 (50% cell survival).
4. Flow cytometric cell cycle analysis
The cell cycle was evaluated by flow cytometric analysis for measuring DNA content with propidium iodide (PI) labeling method as previously described.14) HCT116 cells (5×105) were seeded on 60 mm dishes, and incubated at 37°C for 24 h, then the cells were treated with the various concentrations (0, 12.5, 25, 50μg/ml) of the extract for additional 24 and 48 h.
5. Flow cytometric analysis of membrane symmetry
In order to evaluate the apoptotic cell death, we measured the externalization of phosphatidylserine, a phospholipid which is precise apoptosis marker by using flow cytometric analysis according to previously described method.14) HCT116 cells (5×105) were seeded on 60 mm dishes, and incubated at 37°C for 24 h, then the cells were treated with the extract (0, 25, 50μg/ml) for 48 h.
6. Morphological change
HCT116 cells (5×105) were seeded on 60 mm dishes, and incubated at 37°C for 24 h, then the cells were treated with the extract (0, 25, 50μg/ml) for 48 h. The cellular morphology was observed by phase contrast microscope, OLYMPUS CK40 (Olympus, Tokyo, Japan). The chromatin condensation was evaluated by fluorescent microscopy after PI staining. After treatment of the extract, both floating and adherent cells were harvested by trypsinization. The cells were then washed with Dulbecco’s phosphate buffered saline (DPBS), and fixed with 70% ethanol at −20°C at least 24 h. Then the cells were washed with DPBS again, and incubated in 300μl of PI staining solution (40μg/ml in DPBS) for 1 h at room tem- perature in darkness. The stained cells were visualized by using fluorescence microscope, TE2000U (Nikon, Kanagawa, Japan).
RESULTS
The ethanolic extract of the aerial parts of A. rossii showed a dose- and time-dependent cell growth inhibition against HCT116 cells. The IC50 values were 44.1 and 25.5 μg/ml for 24 and 48 h of post-treatment, respectively (Fig. 1). We further examined that the molecular mechanism of cell growth
Fig. 1. Antiproliferactive actitivity of the ethanolic extract of A.
rossii in HCT116 cells. HCT116 cells were treated with the indicated concentrations (0∼200 μg/ml) of the extract for 24 and 48 h. Cell viability was determined by MTT assay. Each value represents mean±SD from three independent ex- periments with triplicate.
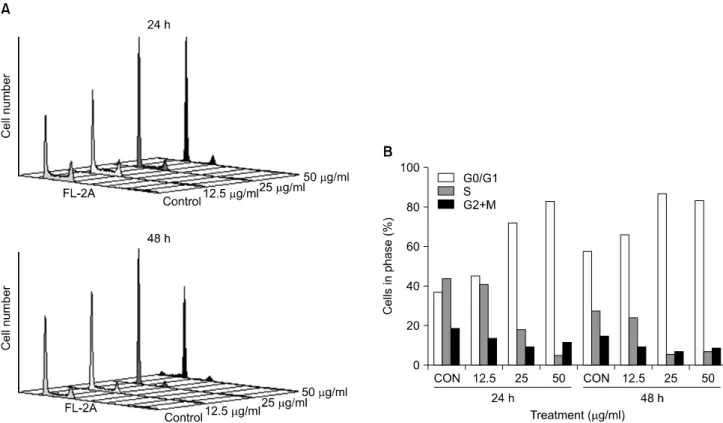
Fig. 2. Effect of A. rossii extract on cell cycle distribution of HCT116 cells. (A) The cell cycle of HCT116 cells treated with the extract (0, 12.5, 25, 50 μg/ml) for 24 and 48 h was evaluated by flow cytometric analysis for measuring DNA content with PI labeling method. The cell cycle graph was schematized by using CellQuest Pro software. (B) The cells in phase (%) of each cell cycle graph in (A) were analyzed by Modfit LT 3.0 software. The data are representatives from three independent experiments.
inhibitory activity of the extract. We hypothesized that the antiproliferative activity of the extract might be exerted
through cell cycle interruption and induction of apoptosis.
The extract strongly induced an increase of cell population at G0/G1 phase accompanied by a decrease of cell population at S phase in a dose-dependent manner. In detail, the extract showed 45.1, 72.4 and 82.9% cells in G0/G1 phase after 24 h treatment at the concentrations of 12.5, 25 and 50μg/ml, respectively, while the vehicle control showed only 37.0% cells in G0/G1 phase. We obtained similar results in the case of 48 h treatment (Fig. 2).
Based on the flow cytometric DNA content analysis, we discovered that the extract induced cell cycle arrest at G0/G1 phase. From the same analysis, we also recognized that the extract induced an increase of the sub G0/G1 content (%) in a dose- and time-dependent manner, which is an indirect clue of apoptosis. The sub G0/G1 content reached to 3.3 and 17.2% after 48 h treatment of 25 and 50μg/ml of the extract, respectively, while the vehicle control stayed at 1.9%. The increase of the sub G0/G1 content of the 48 h-post treatment was stronger than that of the 24 h-post treatment (Fig. 3A).
The extract also induced the externalization of phosphatidy-
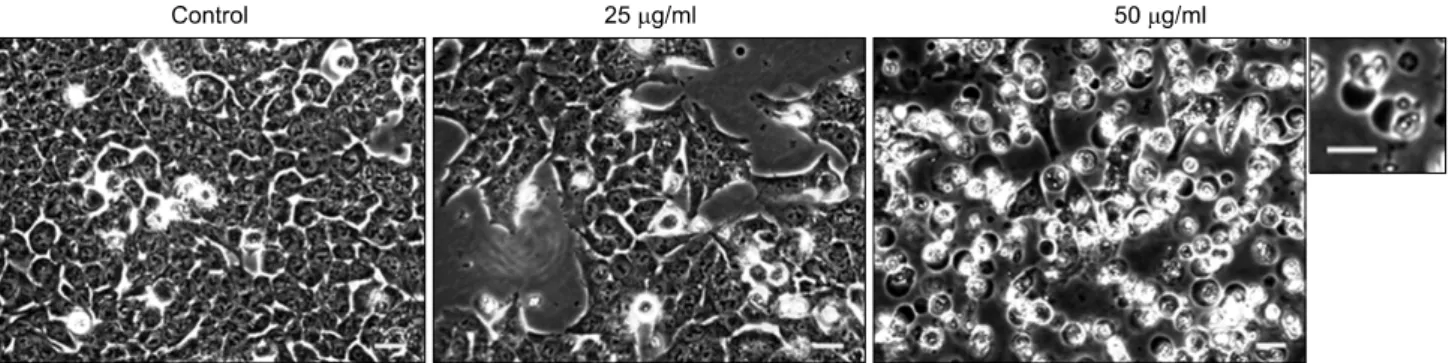
Fig. 4. Morphological changes of HCT116 cells treated with A. rossii extract. HCT116 cells were treated with the extract (0, 25, 50 μg/ml) for 48 h. Phase contrast microscope images were photographed. The magnified picture of apoptotic cells induced by the treatment of 50 μg/ml of the extract was represented on the right side of the original picture (bar=20 μm). The pictures were representatives from three independent experiments.
Fig. 3. Proapoptotic activity of A. rossii extract in HCT116 cells. (A) The sub G0/G1 content (%) of HCT116 cells treated with the extract (0, 25, 50 μg/ml) for 24 and 48 h was measured by flow cytometric analysis after PI staining. The data was obtained by using CellQuest Pro software. The graph is a representative from three independent experiments. (B) Externalization of phosphatidylserine of HCT116 treated with the extract (0, 25, 50 μg/ml) for 48 h was measured by flow cytometric analysis after annexin V-fluorescein and PI staining. The cell population (%) of each quadrant is shown, especially the lower right quadrant (annexin V-fluorescene positive and PI negative) represents early apoptotic cell population (%) indicated in bold numbers.
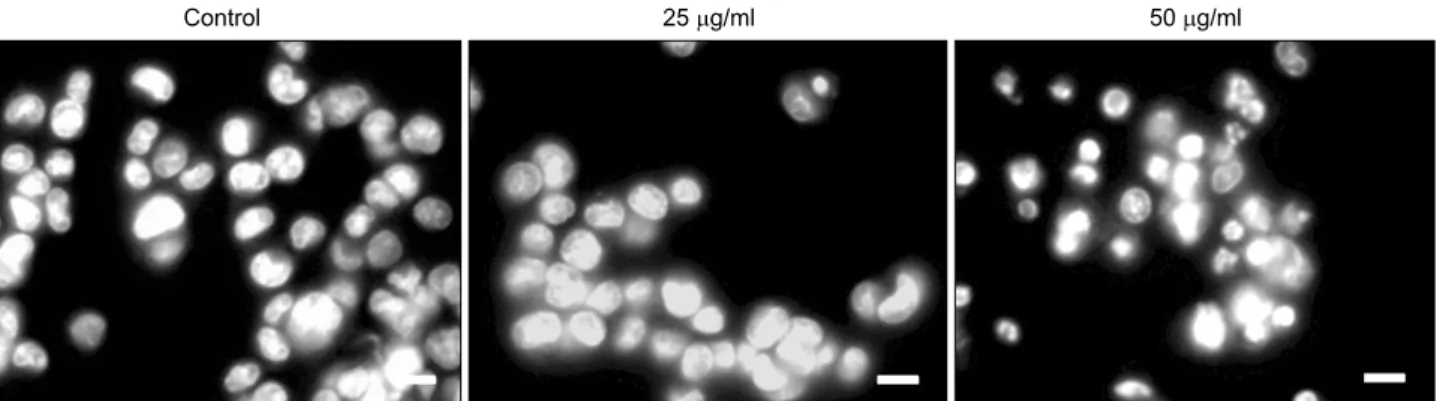
Fig. 5. Chromatin condensation of HCT116 cells treated with A. rossii extract. HCT116 cells were treated with the extract (0, 25, 50 μg/ml) for 48 h and then stained with PI. Chromatin condensation was examined by fluorescence microscopy (bar=10 μm).
The pictures are representatives from three independent experiments.
lserine.The lower left quadrant (annexin V-fluorescein and PI negative, normal phosphatidylserine without loss of the membrane integrity) means normal cell population, which was getting lower by the treatment of the extract. At the same time, the early apoptotic cell population (lower right quadrant, annexin V-fluorescein positive and PI negative, inverted phosphatidylserine without loss of the membrane integrity) were increased to 5.16 and 21.43% by the treatment of the extract at 25 and 50μg/ml for 48 h, respectively, while the vehicle control showed only 2.21%, early apoptotic cell po- pulation. The necrotic or late apoptotic cell population (annexin V-fluorescein and PI positive, inverted phosphatidylserine with loss of the membrane integrity) increased to 16.88% by the treatment of the extract at 50μg/ml for 48 h, on the other hand, the vehicle control showed only that of 2.85% (Fig. 3B).
We also observed the dramatic apoptotic morphological changes including the formation of apoptotic bodies by the treatment of the extract in HCT116 cells. The number of cells in the treatment of 25μg/ml of the extract was much lower than that of the vehicle control. At the concentration of 50μg/ml of the extract, the cells detached from the bottom of culture plate and formed the apoptotic bodies (Fig. 4). The extract at 50μg/ml also triggered the chromatin condensation as a result of the induction of apoptosis (Fig. 5).
DISCUSSION
The various pharmacological activity of A. rossii is known to be resulted from triterpenoids present in the plant. For exam- ple, 3α-hydroxyolean-12-en-27-oic acid, 3β-hydroxyolean-12- en- 27-oic acid, 3α,23-dihydroxyolean-12-en-27-oic acid (aceri-
phyllic acid A) isolated from A. rossii inhibit an ACAT activity, resulting in anti-arteriosclerosis effect.8) In addition, 3α-hyd- roxyolean-12-en-27-oic acid, 3β-hydroxyolean-12-en-27-oic acid, 3α,23-diacetoxyolean-12-en-27-oic acid show the anti-complement activity, and they may be used for treatment of inflammatory diseases.10)
Triterpenoids are thought to be potential agents for the prevention and treatment of cancer and are regarded as new class of multifunctional drugs. For instance, lupane- and oleanane-type terpenoids have been reported to possess cancer chemopreventive activity in animal studies.15∼17) Triterpenoids are also important for cytotoxicity of A. rossii.9) Especially 3β- hydroxyolean-12-en-27-oic acid induces apoptotic cell death both in human ovarian carcinoma HO-8910 cells and human colorectal carcinoma COLO-205 cells. 3β-Hydroxyolean-12-en- 27-oic acid also induces G0/G1 phase arrest in COLO-205 cells.18,19) 3-Oxoolean-12-en-27-oic acid induces apoptosis in human promyelocytic leukemia HL-60 and human cervix adenocarcinoma HeLa cells.12) In this study we found that the extract of A. rossii exerted antiproliferation activity via G0/G1 arrest and apoptosis.
The induction activity of G0/G1 arrest and apoptosis may be due to triterpenoids including 3β-hydroxyolean-12-en-27-oic acid and 3-oxoolean-12-en-27-oic acid which could be isolated from A. rossii.
In addition, it is reported that flavonoids isolated from A.
rossii exert anti-cancer activity through an inhibition of farnesyl protein transferase activity, and these isoflavonoids also have potent antioxidant activities.7,20) The cancer suppressing effects of the ethanolic extract A. rossii may be originated from whole numerous phytochemicals comprising triterpenoids and flavo- noids. Specific combinations of phytochemicals or whole plant extracts sometimes exerted stronger antioxidant and antipro-
liferative effects than isolated compounds alone. The additive and synergetic effects often bring out more potent chemopre- ventive effect.2,3) Therefore, it is necessary to investigate cancer chemopreventive effects of whole plant extracts as well as single phytochemicals.
CONCLUSION
The ethanolic extract of the aerial parts of A. rossii inhibited cell proliferation of HCT116 human colorectal adenocarcinoma cells in a dose- and time- dependent manner. The antiproli- ferative activity of the extract exerted through the induction of cell cycle arrest at G0/G1 phase and apoptosis. Therefore, the ethanolic extract of A. rossii may be developed as a poten- tial chemopreventive agent in human colorectal cancer, which acts on promotion and progression step of tumorigenesis.
ACKNOWLEDGEMENT
This study was supported by KIST Gangneung Institute intramural research grant (2Z03160).
REFERENCES
1) Surh YJ. Cancer chemoprevention with dietary phytochemi- cals. Nat Rev Cancer 3, 768-780, 2003.
2) de Kok TM, van Breda SG, Manson MM. Mechanisms of combined action of different chemopreventive dietary compo- unds: a review. Eur J Nutr 47(2 Suppl), 51-59, 2008.
3) Liu RH. Potential synergy of phytochemicals in cancer preven- tion: mechanism of action. J Nutr 134, 3479S-3485S, 2004.
4) Russo GL. Ins and outs of dietary phytochemicals in cancer chemoprevention. Biochem Pharmacol 74, 533-544, 2007.
5) Lee YM. Korean wild flower. Seoul, Darunsesang, pp 61-64, 2003.
6) Zheng CJ, Sohn MJ, Kim KY, Yu HE, Kim WG. Olean- 27-carboxylic acid-type triterpenes with potent antibacterial activity from Aceriphyllum rossii. J Agric Food Chem 56, 11752-11756, 2008.
7) Han JT, Bang MH, Chun OK, Kim DO, Lee CY, Baek NI.
Flavonol glycosides from the aerial parts of Aceriphyllum rossii and their antioxidant activities. Arch Pharm Res 27, 390-395,
2004.
8) Han JT, Kim HY, Park YD, Lee YH, Lee KR, Kwon BM, Baek NI. Aceriphyllic acid A, A new ACAT inhibitory triter- penoid, from Aceriphyllum rossii. Planta Med 68, 558-561, 2002.
9) Lee I, Yoo JK, Na M, Min BS, Lee J, Yun BS, Jin W, Kim H, Youn U, Chen QC, Song KS, Seong YH, Bae K.
Cytotoxicity of triterpenes isolated from Aceriphyllum rossii.
Chem Pharm Bull (Tokyo) 55, 1376-1378, 2007.
10) Min BS, Lee I, Chang MJ, Yoo JK, Na M, Hung TM, Thuong PT, Lee J, Kim JH, Kim JC, Woo MH, Choi JS, Lee HK, Bae K. Anticomplementary activity of triterpenoids from the whole plant of Aceriphyllum rossii against the classical pathway. Planta Med 74, 726-729, 2008.
11) Ahn EM, Han JT, Kwon BM, Kim SH, Baek NI. Anti-cancer activity of flavonoids from Aceriphyllum rossii. J Korean Soc Appl Biol Chem 51, 309-315, 2008.
12) Kim SS, Won SJ, Kim NJ, Yoo JK, Bae K, Lee KT. 3- Oxoolean-12-en-27-oic acid isolated from Aceriphyllum rossii induces caspase-8-dependent apoptosis in human promyelocy- tic leukemia HL-60 cells. Biol Pharm Bull 32, 91-98, 2009.
13) Lee SB, Cha KH, Selenge D, Solongo A, Nho CW. The chemopreventive effect of taxifolin is exerted through ARE- dependent gene regulation. Biol Pharm Bull 30, 1074-1079, 2007.
14) Kang K, Lee HJ, Kim CY, Lee SB, Tunsag J, Batsuren D, Nho CW. The chemopreventive effects of Saussurea salicifolia through induction of apoptosis and phase II detoxification enzyme. Biol Pharm Bull 30, 2352-2359, 2007.
15) Fukuda Y, Sakai K, Matsunaga S, Tokuda H, Tanaka R.
Cancer chemopreventive activity of lupane- and oleanane-type triterpenoids from the cones of Liquidamber styraciflua. Chem Biodivers 2, 421-428, 2005.
16) Liby KT, Yore MM, Sporn MB. Triterpenoids and rexinoids as multifunctional agents for the prevention and treatment of cancer. Nat Rev Cancer 7, 357-369, 2007.
17) Setzer WN, Setzer MC. Plant-derived triterpenoids as potential antineoplastic agents. Mini Rev Med Chem 3, 540-556, 2003.
18) Sun HX, Ye YP, Pan YJ. Cytotoxic oleanane triterpenoids from the rhizomes of Astilbe chinensis (Maxim.) Franch. et Savat. J Ethnopharmacol 90, 261-265, 2004.
19) Tu J, Sun HX, Ye YP. 3 Beta-hydroxyolean-12-en-27-oic acid: a cytotoxic, apoptosis-inducing natural drug against COLO-205 cancer cells. Chem Biodivers 3, 69-78, 2006.
20) Ahn EM, Han JT, Kwon BM, H. KS, Baek NI. Anti-cancer Activity of Flavonoids from Aceriphyllum rossii. J Korean Soc Appl Biol Chem 51, 309-315, 2008.