Introduction
The genus Hemibarbus of Korea comprised of three species: H. labeo, H. longirostris and H.
mylodon. These species are also known as semi- bottom dwellers (Hosoya, 1986). Among them H.
labeo and H. longirostris which live in the most middle area of rivers were widely distributed in Korea, but H. mylodon, well known as Korean endemic species and natural monument Nos. 238 and 259, was restricted only within the narrow region of upper part in Han river and Keum river in Korea (Kim, 1997). Taxonomic studies for these fishes has been accomplished based on the exomorphological and osteological characters by many workers (Banarescu and Nalbant, 1973;
Kim, 1984, 1997; Hosolya, 1986; Howes, 1991;
Kang, 1991). Particularly Ramaswami (1955a) and Kang (1991) reported about taxonomic as- pects based on the osteological characters of the skull and weberian apparatus in the cyprinids,
and Kang and Park (1973) researched about the karyotypes and DNA values of gobioniid fishes including Hemibarbus. Recently Kim (1984) reas- signed the taxonomic position of the species from Gonoproktopterus mylodon into H. mylodon on the basis of several morphometric characters.
But until now detailed osteological comparison of these three species in Hemibarbus from Korea has not been executed yet. The aim of this study is to describe the interspecific morphological cha- racters of their head part among three species.
Materials and Methods
For this study, all skeletal specimens were made by according to Taylor’s (1967) revised me- thod. All observations and figures were accom- plished by a binocular dissecting stereo-micro- scope set with camera lucida (Wild M8 and Olympus SZH-10) and lengths of skeleton were measured by dial caliper (1/20 mm) and microme-
─
─ 212 ──
Skeletal Morphology of the Head in the Genus Hemibarbus (Cypriniformes: Cyprinidae) from Korea
Chung-Lyul Lee and Yong-Ho Kim
Department of Biology, College of Natural Science, Kunsan National University, Kunsan 573-701, Korea
We investigated morphological characters of the head skeleton in three species of the genus Hemibarbus from Korea, H. labeo, H. longirostris and H. mylodon. We found many interspecific differences in morphological and sutural features among the three species: in the cranial region, forms of preethmoid, prevomer, supraeth- moid, basioccipital and type of suture in both frontal and parietal; in the circumor- bital region, the form of lacrimal bone and the opening direction of the sensory canal of the 4th and 5th infraorbital; in the branchio--hyal region, forms of basihyal and urohyal, and shapes of pharyngeal teeth and articulating face between interhyal and epihyal.
We consider an appropriate taxonomic position of H. mylodon, an endemic species from Korea, is the genus Hemibarbus, although it shows in its osteology several characteristics that separate it from the other Korean species of the genus.
Key words : Head, osteology, Hemibarbus, H. labeo, H. longirostris, H. mylodon
ter. The terminologies of skeleton followed Rawaswami (1955a, b) and Howes (1980). Exa- mined specimens for this study is as follows:
H. labeo: 83.0~86.2 mm standard length (SL), (2 specimens), Sept. 22, 1984, Ganggyung-eup Nonsan-si, Chungcheongnam-do (Keum river);
111.4~116.3 mm SL, (2), Oct. 26, 1984, Munsan -eup, Paju-gun, Kyonggi-do (Imjin river); 69.3
~99.9 mm SL, (17), Dec. 13, 1991, Kangkyeong- eup Nonsan-si, Chungcheongnam-do (Keum river); 120.6~143.6 mm SL, (3), March 3, 1995, Paldang, Wabu-eup, Namyangju-si, Kyeonggi- do (Han river).
H. longirostris: 114.6~125.5 mm SL, (2), March 3, 1995, Paldang, Wabu-eup, Namyangju-si, Kyeonggi-do (Han river); 66.1~90.5 mm SL, (4), Jun. 4, 1983, Jangseong-eup, Jangseong-gun, Jeollanam-do (Youngsan river); 70.9~100.7 mm SL, (12), July 31, 1983, Sagok-myeon, Gongju- gun, Chungcheongnam-do (Keum river); 84.4~
100.5 mm SL, (4), Aug. 23, 1980, Unam-myeon, Imsil-gun, Jeollabuk-do (Keum river).
H. mylodon: 135~136 mm SL, (2), Oct. 20, 1999, Youngwoul-eup, Youngwoul-gun, Kangwon -do (Han river).
Results
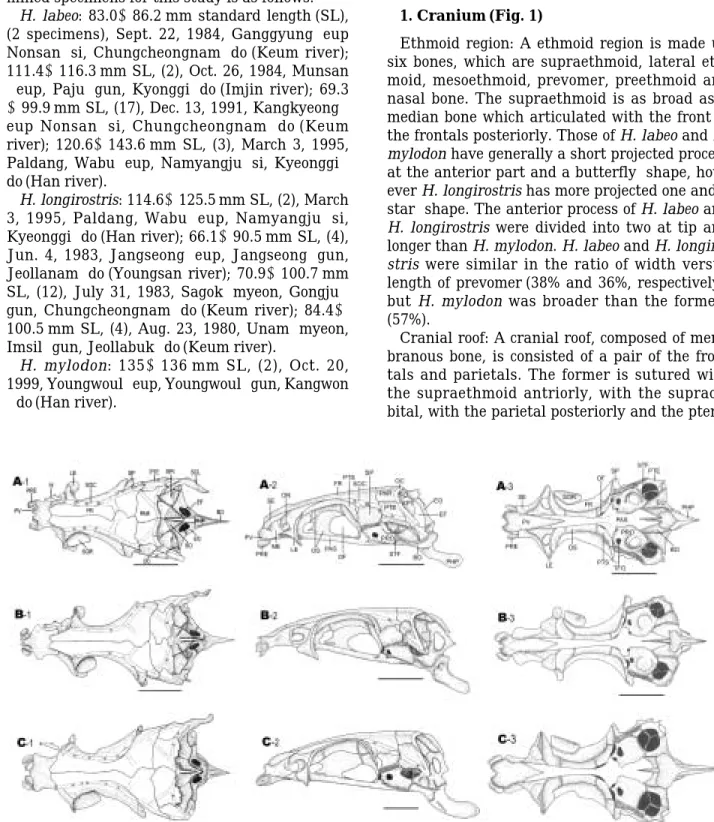
1. Cranium (Fig. 1)
Ethmoid region: A ethmoid region is made up six bones, which are supraethmoid, lateral eth- moid, mesoethmoid, prevomer, preethmoid and nasal bone. The supraethmoid is as broad as a median bone which articulated with the front of the frontals posteriorly. Those of H. labeo and H.
mylodon have generally a short projected process at the anterior part and a butterfly-shape, how- ever H. longirostris has more projected one and a star-shape. The anterior process of H. labeo and H. longirostris were divided into two at tip and longer than H. mylodon. H. labeo and H. longiro- stris were similar in the ratio of width versus length of prevomer (38% and 36%, respectively), but H. mylodon was broader than the formers (57%).
Cranial roof: A cranial roof, composed of mem- branous bone, is consisted of a pair of the fron- tals and parietals. The former is sutured with the supraethmoid antriorly, with the supraor- bital, with the parietal posteriorly and the ptero-
Fig. 1. Comparison of dorsal (1), lateral (2) and ventral views (3) of skull of Hemibarbus labeo (A) , H. longirostirs (B) and H. mylodon (C), Scale bars indicate 5 mm. BO, Basioccipital; EF, fenestra in the exooccipital; EO, exooccipital; EPI, epiotic; FR, frontal; LE, lateral ethmoid; N, nasal; OC, occipital canal; PAR, Parietal; PHP, pharyngeal process;
PRE, preethmoid; PTE, pterotic; PV, prevomer; SCL, supracleithrum; SO, supraoccipital; SOC, supraorbital canal;
SOR, supraorbital; SP, sphenoid.
tic post-laterally in the orbital region. In the direction of cranial suture, H. labeo and H. mylo- don were located on the right, but H. longiro- stris, on the left.
Orbital region: An orbital region consisted of the orbitosphenoid, the orbitosphenoid septum, the pterosphenoid and the parasphenoid. The orbitosphenoid has a Y-form when cuts the up- per part of it. It is anteriorly joined with the la- teral ethmoids and posteriorly with the ptero- sphenoids. On the other hand, the parasphenoid is joined with the prevomer posteriorly, the proo- tic post-laterally and the basioccipital posteri- orly in the ventral view. In H. longirostris and H.
mylodon, a suture of the pterosphnoid and orbi- tosphenoid extended to the orbitosphenoid sep- tum, but H. labeo did not. The sutures of bet- ween parasphenoid and basioccipital, parasph- enoid in H. labeo and H. longirostris are shown a sharp forked shape, but H. mylodon was a round.
Otic region: An otic region consists of a pair of
the prootics, sphenotics, pterotics and epiotics.
The prootic possesses a trigeminofacial orifice and is broadly sutured with the parasphenoids in the anterior part. The sphenotic is joined with the pterosphenoid at the post-region. While it is joined with the exooccipital and basioccipital pos- teriorly. The pterotic is joined with the frontal at the posterior margin, with the parietal dorsally, and with the prootic and exoccipital at the below.
It is also articulated with the hyomandibular.
Occipital region: An occipital region consisted of the supraoccipital, the exoccipital, the basi- occipital and intercalar. The supraoccipital arti- culated with a pair of the parietal anteriorly, the epiotic at left and right and the exoccipital pos- teriorly. The exoccipital sutured with the basio- ccipital ventrally is the large bone at the occipi- tal region and has a large exoccipital foramen.
2. Circumorbital bones (Fig. 2)
The circumorbital bones consisted of a supraor-
Fig. 2. Comparison of lateral view of circumorbital series of Hemibarbus labeo (A), H. longirostris (B) and H. mylodon (C).
Scale bars indicate 5 mm. IO, infraorbital; SOR, supraorbital.
Fig. 3. Comparison of lower (dorsal view) and upper jaw bones (lateral view) of Hemibarbus labeo (A), H. longirostris (B) and H. mylodon (C). Scale bars indicate 5 mm. AN, angular; DEN, dentary; MAX, maxilla; PMX, premaxilla; R, rostral; RA, retroarticular.
bital, lacrimal (1st infraorbital) and four infraor- bitals. All infraorbitals which made into mem- branous bone form incomplete sensory canals, where they opened downward from 1st to 3rd infraorbitals, but those of 4th and 5th ones differ at direction of its opening according to the spec- ies. The opening type of sensory pore at the 4th and 5th infraorbitals varies according to species.
That is, that of H. labeo opened at 2 o’clock and H. mylodon, 12 o’clock, but H. longirostris has a complete circled sensory canal without opening.
The upper process of lacrimal is articulated with lateral ethmoid. Those of H. labeo and H.
mylodon were quadrangular type without dis- tinct process dorsally, but H. longirostris, a fairly pointed process dorsally (Fig. 2).
3. Jaws (Fig. 3)
The jaws consisted of the premaxilla, maxilla, rostral, dentary, angular and retroarticular. The premaxilla was situated at the more fore part than other bones showed L-form in the lateral view and the ascending process of premaxilla and rostral was linked by a ligament. The dorsal -anterior process of maxilla was linked with palatine by a ligament.
The dentary possesses three or four sensory pores. H. labeo and H. mylodon have three, but H. longirostris has four. The first sensory pore was placed at the end of anterio-upper part of dentary while the remaining pores were placed at medial part of it.
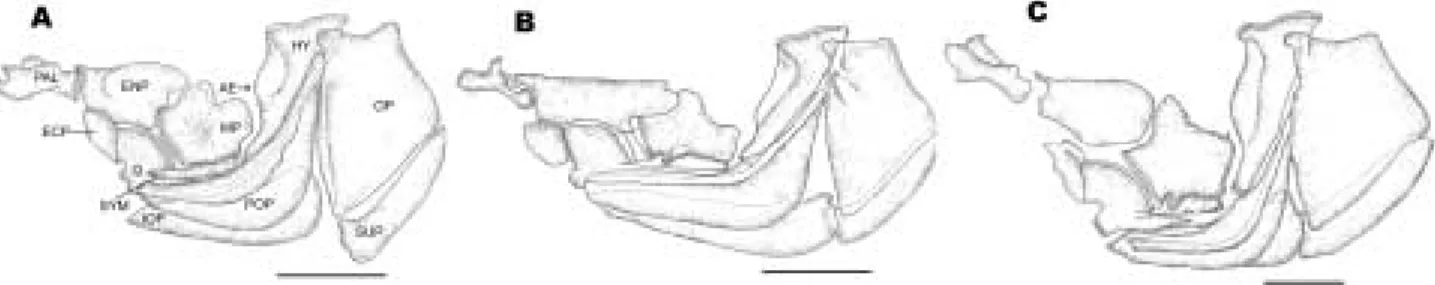
4. Suspensorial elements and opercular bones (Fig. 4)
The suspensorium bones consisted of the pala- tine, entopterygoid, ectopterygoid, metaptery- goid, quadrate, hyomandibular and symplectic.
The opercualr bones contained the preopercular, interopercular, opercular and subopercular. They were also attached by a membrane, so their loco- motions were free compared to other bones. The palatine articulates with the preethmoid later- ally. Posteriorly it was connected with the entop- terygoid. Also anterior part of this bone was arti- culated with the dorsal-anterior process of maxi- lla by ligament. The entopterygoid was joined with the ectopterygoid anterio-ventrally, quad- rate ventrally and metapterygoid posteriorly.
The ratios of width to the length of entopterygoid showed 0.56 in H. labeo, 0.33 in H. longirostris and 0.54 in H. mylodon respectively. The metap- terygoid is located in the center of suspensorial elements. It is articulated with the entopterygoid dorso-anteriorly, the quadrate ventro-anteri- orly, the symphysis ventrally and the under side of hyomandibular posteriorly. The hyomandi- bular is articulated with the lateral facet of cra- nium part which is the sphenotic anteriorly and sphenotic-pterotic region posteriorly. In three species, H. labeo and H. logirostris possessed a cartilaginous articulation with the upper region of it on the whole, but H. mylodon had a cartila- ginous articulation partly at anterior part. The symphysis was a slender bone which is articulat- ed with the quadrate anteriorly, metapterygoid dorsally and connected by a ligament with the under facet of metapterygoid and hyomandi- bular. In the joining part of metapterygoid, it possessed one or two process in Hemibarbus. H.
labeo had two which was very small, but H. lon- girostris and H. mylodon had only one.
The opercular series consisted of the opercular, preopercular, interopercular and subopercular.
The opercular which covered the brachial part was the largest of the series and usually it show- ed an articular process towards the hyomandi-
Fig. 4. Comparison of suspensorial elements and opercular series of Hemibarbus labeo (A), H. longirostris (B) and H.
mylodon (C). Scale bars indicate 5 mm. ECP, ectopterygoid; ENP, entopterygoid; HY, hyomandibular; AE, anterior edge of hyomnadibular; IOP, interopercular; MP, metapterygoid; OP, opercular; PAL, palatine; POP, preopercular;
Q, quadrate; SUP, subopercular; SY, symplectic.
bular anteriorly. Anterior process of the hyoman- dibular had a short sensory canal which is con- necting with the preopercular canal.
The preopercular which is sickle-form passed the sensory canal into the central region. The interopercular which extended fairly long limb towards the preopercular is attached by a mem- brane in the under side of preopercular.
5. Hyoid arch (Fig. 5 and 6)
The hyoid arch composed of seven elements which is the interhyal, the epihyal, the cerato- hyal, the hypohyal, the basihyal, the branchio- stegal rays and the urohyal. The interhyal was a fixed rod bone that is articulated with the hyo- mandibular anteriorly and epihyal posteriorly.
The epihyal was a flat and conented with the triangular bone posterior to the ceratohyal. In articulating face between the interhyal and the epihyal, H. labeo and H. longirostris was flat, but H. mylodon had sunken. The hypohyal is atta- ched to ceratohyal and joined with the haemal arch process of urohyal by a ligament. The bran- chiostegals was consisted of three pairs blade- form bone and the 3th one was largest. They are
attached to the ceratohyal and the epihyal re- spectively. The basihyal was rod-type bone, the anterior part of it is covered by a cartilage and does articulated with the 1st copulae posteriorly.
From a morphological characters of the basihyal, H. labeo and H. mylodon was a similar, which was wide and stubby anteriorly and narrow pos- teriorly, but H. longirostris was a long and nar- row at before and behind. The ratio of width to length of the basihyal are 0.43 in H. labeo, 0.23 in H. longirostris and 0.52 in H. mylodon.
The urohyal which a single bone is articulated with the hypohyal by strong ligament anteriorly and joined with the cleithrum by the sterno- hyoideus muscle. In all species of Hemibarbus, the vertical plate of it is well-developed and shape of the horizontal plate of H. labeo and H.
longirostris were broad and round, but H. mylo- don was narrow and flat. Its tip of H. labeo and H. longirostris were pointed type, but H. mylo- don truncated.
6. Branchial arch (Fig. 7 and 8)
Branchial arch consisted of the three copulae, the cartilaginous projection, the hypobranch, the
Fig. 5. Comparison of hyoid bones of Hemibarbus labeo (A), H. longirostris (B) and H. mylodon (C). Scale bars indicate 5 mm. BHY, basihyal; CHY, ceratohyal; EHY, epihyal; HYE, hypohyal; IH, interhyal; BR, branchiostegal.
Fig. 6. Comparison of dorsal (upper) and lateral (bottom) view of urohyal of Hemibarbus labeo (A), H. longirostris (B) and H. mylodon (C). Scale bars indicate 2 mm. HA, hemal arch; HP, horizontal plate; VP, vertical plate.
ceratobranchial, the epibranchial, the pharyn- gobranch and the pharyngeal teeth.
The copulae which composed of three flat bone together cartilaginous projection made a row at median line of bottom of the branchial cavity.
The hypobranch was small and paired bones that joined with the ceratobranchial and the copulae.
The ceratobranchials which consisted of four paired elongated bones show an identical shape in three species.
Each bone is articulated with the epibranchials being smaller than it. All species have the gill ra- kers attached to the epibranchials and cerato- branchials. In the outside number of gill rakers of first branchial arch, H. labeo has nineteen (6±
16), H. longirostris, seven (1±6) and H. mylodon,
ten (3±7).
The arch-like pharyngeal bone bearing a strong process took several types of pharyngeal teeth. The pharyngeal teeth in all species had three rows and usually represented as 5, 3, 1-1, 3, 5 in dental formula. The pharyngeal teeth showed considerable variation both in the size of first tooth and the shape of second tooth. The 1st one is well developed in H. labeo and H. longiro- stris, but much reduced in H. mylodon; in the second one of H. labeo and H. longirostris, the tip of tooth was very pointy and its masticatory groove presented a inclining form, but in H. my- lodon, masticatory face showed horizontally without any pointed process.
Fig. 7. Comparison of branchial arches of Hemibarbus labeo (A), H. longirostris (B) and H. mylodon (C). Scale bars indicate 5 mm. C1~3, copulae 1~3; CB1~4, cerobranchial 1~4; CP, cartiaginous projection; EB1~4, epibranchial 1~4;
HB1~3, hypobranchs 1~3; PB1~2, infrapharyngobrnachial 1~2.
Fig. 8. Comparisons of lateral (upper) and dorsal (bottom) view of branchial bones of Hemibarbus labeo (A), H. longirostris (B) and H. mylodon (C). Scale bars indicate 2 mm. OG, occlusal grooves.
Discussion
Up to the present, the morphological analyses of skeleton have been treated as a very impor- tant work for classification of fishes (Harrington, 1975; Ramaswami, 1955a; Howes, 1980; Sawada, 1982; Hosoya, 1986; Lee, 1988; Kang, 1991; Kim, 2000). In the many characters of fishes, the morphology of skull composed of many piece of membranous bones can be shown different or unique characters according to the taxon.
In the cranial morphology, the anterior process of supraethmoid of Hemibarbus is longer than other species of the subfamily Gobioninae (Rama- swami, 1995a; Kang, 1991), but that of H. mylo- don is much shorter than other two species of Hemibarbus.
Howes (1980), Kang (1991) and Ramaswami (1995b) described importantly about the shape of the vomer region having the groove of anterior part of prevomer and posterior narrow limb of the vomer in the family Cyprinidae. We also thought that relationship between width and length of it have to deal as a important taxono- mic character (Table 1).
Particularly Ramaswami (1955a) reported that the fishes of Hemibarbus had a longer suprae- thmoid and a large rostral process than other fishes of Gobioninae. Also Howes (1985) regarded the shape of supraethmoid as a important cha- racter in the definition of Phoxinus. Anterior pro- cess of supraethmoid among the fishes of Hemi- barbus from Korea varied on their morphological features: H. labeo has short and pointed; H.
longirostris, long and pointed; H. mylodon, short
and blunt.
Lee (1988) mentioned the differences of cranial suture and of cranial morphology in the bagird fishes as important taxonomic characters: su- tural shapes between the ethmoid and the fron- tal; both the frontal; the frontal and the parietal;
the parasphenoid and the basioccipital. In the direction of cranial suture, H. labeo and H. mylo- don were same direction as right side, but H.
longirostris was the opposite.
The form of basihyal had reported as an impor- tant criterion of phylogeny in the family Acan- thridae (Guiasu and Winterbottom, 1993). But like present note of basihyal feature at Fig. 5, three species of Hemibarbus from Korea are divided into two groups: that of H. labeo and H.
longirostris were same style but H. mylodon was not.
Hosoya (1986) stated that the Hemibarbus has a peculiar features in cranium region: the seg- mentation of the infraorbital canal which is dee- ply roofed by the bony brim and the preoperculo- mandibualr canal. Also in result of this study, these features were in accord with his opinion.
But their morphology showed huge differences among species of Hemibarbus. For example, opening direction of sensory canal at the 4th and 5th infraorbital did not correspond among three species (Table 1, Fig. 2). On the other hand, Nam and Yang (1998) stated that the 2nd infraorbital of H. mylodon is narrower than other species of Hemibarbus. Regan (1911) referred to the posses- sion of a complete series of circumorbital as a primitive characters, but Ramaswami (1955b) mentioned that he was unable to follow Regan’s opinion. In the three species of Hemibarbus, a
Table 1. Comparisons of morphology and morphometric characters of skull among the three species of Hemibarbus from Korea
Characters H. labeo H. longirostris H. mylodon
Anterior process of supraethmoid short and pointed long and pointed short and blunt
Ratio of width to length of prevomer 0.38 0.36 0.57
Direction of cranial suture right left right
State of supraoccipital process serration smooth smooth
Numbers of sensory pore of dentary 3 4 3
Opening direction of 4th and 5th infraorbital cannal 2 o’clock almost complete circle 12 o’clock
Ratio of width to length of entopterygoid 0.54 0.33 0.54
Articular face between interhyal and epihyal f lat f lat sunken
Shape of basihyal anterior-wide, anterior and anterior-wide,
posterior-narrow posterior-narrow posterior-narrow
Ratio of width to length of basihyal 0.43 0.23 0.52
Shape of urohyal broad and round broad and round narrow and flat
Form of urohyal tip pointed pointed truncated
First tooth of pharyngeal teeth development development weak
complete circumorbitals composed of five ossicles which have the irregular broad and incomplete sensory canal. We considered that a circumorbi- tal series of Hemibarbus are to be in a primitive condition in the forms and structures as criterion of Regan (1911).
In the pharyngeal teeth, Hemibarbus had a same dental formula as three rows. In the grind- ing face of the 2nd pharyngeal teeth, H. mylodon had much different shape when it was compared with those of H. labeo and H. longirostris (Table 1, Fig. 8). Ramaswami (1955a) has mentioned that the number of teeth rows becomes criteria as indicator of primitive condition of fishes. Here three rows of Hemibarbus were considered as the most primitive type (Regan, 1908; Chu, 1935;
Nichols, 1943).
In the urohyal, the shape of the horizontal and vertical plate of H. labeo and H. longirostlis was similar with each other, but H. mylodon was not (Table 1). Kim and Kang (1989) reported that the horizontal plate of Hemibarbus has narrower or a more elongated rhombus shape which is point- ed posteriorly and its width is narrower than the hight of vertical plate. But of the fishes belong- ing to Hemibarbus, H. mylodon showed remar- kable feature at posterior part of horizontal plate.
In this study, although H. mylodon has many different characters when compared with other two species, H. labeo and H. longirostris, we con- sidered our study as an appropriate result that H. mylodon which was endemic species from Korea is classified into the genus Hemibarbus, based on the common characters of the struc- tural feature of infraorbital canal, the three rows of pharyngeal teeth and the state of last spinal tip of dorsal fin among three species.
Acknowledgments
We are very thankful to Dr. Wan-Ok Lee of Chongpyong Inland Fisheries Research Institute for supplying specimens of Hemibarbus mylodon.
References
Banarescu, P. and T.T. Nalbant. 1973. Pisces, Teleostei, Cyprinidae (Gobioninae). Das Tierreich. Lieferung 93.
Walter de Gruyter. Berlin, 288 pp.
Chu, Y.T. 1935. Compative studies on the scales and on the
pharyngeals and their theeth in Chinese Cyprinids with particular reference to taxonomy and evolution.
Biol. Bull. St. John’s Univ. Shanghai, 2: 225~?.
Guiasu, R.C. and R. Winterbottom. 1993. Osteological evidence for the phylogeny of recent genera of surgeon- fishes (Percomorpha, Acanthuridae). Copeia, 1993(2) : 300~312.
Harrington, R.W.Jr. 1975. The osteocranium of the Ameri- can cyprinid fish, Notropis bifrenatus, with an annotat- ed synonymy of teleost skull bones. Copeia, 1975(4) : 267~291.
Hosoya, K. 1986. Interrelationships of the Gobioninae (Cyprinidae), Indo-Pacific Fish Biology. In: Proceed- ings of the second international conference on Indo- Pacific Fishes. Ichthyology Society of Japan, Tokyo, pp.
484~501.
Howes, G.J. 1980. The anatomy, phylogeny and classifi- cation of bariline cyprinid fishes. Bull. Br. Mus. Nat.
Hist. (Zool.), 37 : 129~198.
Howes, G.J. 1985. A revised synonymy of the minnow genus Phoxinus Rafinesque, 1820 (Teleostei: Cyprini- dae) with comments on its relationships and distribu- tion. Bull. Br. Mus. nat. Hist. (Zool.), 48(1) : 57~74.
Howes, G.J. 1991. Systematics and biogeography; an over- view. In: I.J. Winfield and J.S. Nelson (eds.). Cyprinid fishes. Systematics. Biology and Exploitation Chapman and Hall, London, New York, Tokyo, Melboune. Mad- ras., pp. 1~33.
Kang, Y.S. and E.H. Park. 1973. Studies on the karyotypes and comparative DNA values in several Korean cypri- nid fishes. Korean J. Zool., 16 : 97~108. (in Korean) Kang, E.J. 1991. Phylogenetic study on the subfamily
Gobioninae (Pisces: Cyprinidae) from Korea as evidenc- ed by their comparative osteology and myology. Ph. D.
Thesis Chonbuk Nat. Univ., pp. 1~108. (in Korean) Kim, I.S. 1984. The taxonomic study of gedgeons of the
subfamily Gobioninae (Cyprinidae) in Korea. Bull.
Korean Fish. Soc., 17(5): 436~448. (in Korean)
Kim, I.S. and E.J. Kang. 1989. Comparative study on the urohyal of the subfamily Gobioninae of Korea. Korean J. Ichthyol., 1(1, 2): 24~34. (in Korean)
Kim, I.S. 1997. Illustrated Encyclopedia of Fauna & Flora of Korea. Vol. 37. Freshwater Fishes. Ministry of Edu- cation., Korea. Seoul. pp. 220~225. (in Korean) Kim, I.J. 2000. Phylogenetic study on the comparative
osteology of the subfamily Acheilognathinae (Pisces:
Cyprinidae) from Korea. Ph. D. Thesis, Chonbuk National University, 90 pp. (in Korean)
Lee, C.L. 1988. Systematic studies on the bagrid catfish, family Bagridae (Pisces: Siluriformes) from Korea. Ph.
D. Thesis, Chonbuk Nat. Univ., pp. 1~141. (in Korean) Nam, M.M. and H.J. Yang, 1998. On the circumorbital
bone of the cyprinid fishes in Korea. Korean J. Ichth- yol., 10(1): 128~138. (in Korean)
Nichols, J.T. 1943. The freshwater fishes of China. Natural History of Central Asia. Vol. IX. Ame. Mus. Nat. Hist.
New York, 322 pp.
Ramaswami, L.S. 1955a. Skeleton of cyprinoid fishes in relation to phylogentic studies. VI. The skull and We- berian apparatus in the subfamily Gobioninae (Cyp- rinidae). Acta. Zool. (Stockholm), 36 : 127~158.
Ramaswami, L.S. 1995b. Skeleton of cyprinoid fishes in relation to phylogenetic study. VII. The skull and We-
berian apparatus of Cyprininae (Cyprinidae). Acta.
Zool. (Stockholm), 36 : 199~242.
Regan, C. 1908. A collection of freshwater fishes from Korea. Proc. Zool. Soc. London, pp. 59-63.
Regan, C. 1911. The classification of the teleostean fishes of the order Ostariophsi. 1. Cyprinoidei. Ann. Ma. nat.
Hist., 8(8) : 13~32.
Sawada, Y. 1982. Phylogeny and zoogeography of the superfamily Cobitoidae (Cyprinoidei, Cypriniformes).
Mem. Fac. Fish. Hokkaido Univ., 28(2) : 65~223.
Taylor, W.R. 1967, An enzyme method of clearing and staining small vertebrates. Pro. of U. S. Nat. Mus., 122 (3596) : 1~17.
Received : August 3, 2002 Accepted : September 23, 2002
한국산 누치
( Hemibarbus )
어류의 두부골격 형태 이 충 렬∙김 용 호군산대학교 생물학과
한국산 누치속 어류 3종인 H. labeo, H. longirostris, 그리고 H. mylodon의 두부골격의 형태적 특징을 조사했다. 그 결과 두부골격의 형태와 봉합방식에서 3종간 서로 상당한 차이가 나타났는 데, 이 중 전사골, 상사골, 전서골, 기저후두골의 형태와 좌우 액골과 노정골 사이의 봉합형태에서 매우 다르게 나타났다. 한편 내장골격 중 안위골의 제1안하골 형태, 또 이들과 측사골과의 관절 부위, 그리고 제4, 5안하골의 감각관 개구 방향, 현수골에서는 중 및 후익상골의 형태, 설궁부에서 는 기설골과 미설골 및 인두치의 형태, 간설골의 관절면 등에서 종간 차이가 잘 나타나고 있었 다. 한편 한국 고유어종인 H. mylodon은 골격의 형태에서 독특한 특징을 나타내고 있었지만, 안 하골의 감각관 구조, 3열의 인두치 그리고 등지느러미 마지막 가시 끝의 상태 등에서 서로 일치 되어 이들3종을 Hemibarbus속에 분류하는 것이 타당하다고 사료된다.