Interpretation of Adaptational Environment of the genus Trapa in Korea Through peat deposit
1Jin-Hee Lee2 , Dae-Jae Kim2*
ABSTRACT
Studies on Trapa of Korean, Japanese, and Chinese origins have been generally limited to currently living Trapa, thereby demonstrating the lack of phylogenetic approaches. In the present study, we conducted a numerical taxonomic study on 34 Korean origin Trapa species newly discovered in peat deposits. By using the numerical taxonomy as well as geological data, we created an appearance map based on the periods and taxonomic groups of the Korean Trapa. Further, by using this appearance map, we reviewed the limitations that exist in the taxa and simultaneously discussed the adaptational environment of Trapa in each period.
Although other studies have discussed new variants of Trapa worldwide, including T. litwinowii V. vassil. var.
chihuensis S.F.Guan & Q.Lang of China (Guan and Lang, 1987), the present study is significant because it provides data on the origin of Korean Trapa species. The availability of such data is essential in determining the adaptational environment of the presently living Trapa. To our knowledge, this is the first report of approximately 10 newly discovered and other presently living species of Trapa. Further, the appearance map in the present study allows the use of the 10 newly discovered species and variants of 34 Trapa species indigenous to South Korea for over 1000 years, for a comparison with the currently living Trapa species. Thus, the present study results could serve as important basic data to understand the adaptational environment of the genus Trapa. from the past to the present in Korea.
KEY WORDS: TRAPACEAE, SYSTEMATICS, CLUSTER ANALYSIS, FACTOR ANALYSIS, GEOLOGIC RESEARCH
1 Received 01 October 2014; Revised (1st: 20 January 2015, 2nd: 03 February 2015); Accepted 04 February 2015 2 Dept. of, Chungbuk National Univ., Cheong ju, 361-763, Korea.
* Corresponding author: Tel: +82-43-261-3260, Fax: +82-43-260-3361, E-mail: [email protected]
INTRODUCTION
Trapa Linnaeus belongs to the family Trapaceae and the species of the genus are annual viny, aquatic floating herbs (Cronquist, 1981). The Trapa is native to the temperate regions of Europe and Asia, and around 30 species (Cook et al., 1974) have been discovered worldwide.
The genus Trapa was set up by Linnaeus in 1753, and subsequently around 10 species were announced in India, China, Manchuria, Japan, and other places. Nakano (1913, 1964) put various Trapa species in China and Japan together, and newly reported the Chinese no-horned
Trapa, and 4 species and 5 variants at the same time.
Nakai (1942) newly reported T. taiwanensis Nakai, T.
pseudoincisa Nakai, and T. jeholensis Nakai. However, since the time of Linnaeus, stem, leaf, and petal of the genus Trapa has not been much different in the shape among each species, there has been no classification standard, and there has been incompatibility in the classification results among scholars, so that there has been a great diversity of opinion on even the number of species.
Because the variation is so wide, it is hard to set limits on the species range. As to the number of taxa of the genus Trapa in Korea, it takes on a complicated aspect, such
70 Jin-Hee Lee, Dae-Jae Kim Korean J. Environ. Ecol. 29(1) 2015
A-san
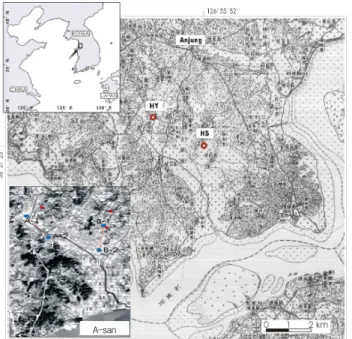
Figure 1. Topographical map for research area.
(HS-Hwangsan-ri, Hyunduck-myeon, Pyeongtaek CT, Gyeonggi-do, Korea: N36° 57′ 30″, E126°
55′ 45″, HY-Hwayang-ri, Hyunduck- myeon, Pyeongtaek CT, Gyeonggi-do, Korea: N36°
57′ 25″, E126° 55′ 52″)
Sample No.
alt.
(m) material Lab. No.
conventional
14C age (14C yrBP)
Trapa nut
HY 6-8 5.115 peat GX32139 4270±50
Absent HY 6-20 4.995 sediment bulk SNU05-369 4540±40
HY 6-40 4.795 sediment bulk SNU05-370 5050±60 HY 5-26 4.435 sediment bulk SNU05-368 5560±40
Present HY 4-6 4.135 sediment bulk SNU05-365 5800±40
HY 4-20 3.995 sediment bulk SNU05-367 6160±40 HY 3-28 3.415 sediment bulk SNU05-364 6560±60 HY 2-16 3.035 sediment bulk SNU05-362 6970±40
Absent HY 2-36 2.835 sediment bulk SNU05-363 7300±60
HY 1-12 2.575 sediment bulk SNU05-361 21040±180
Table 1. Sample list with conventional radiocarbon dates (14C yrBP)
as an omission of a species, changes in a botanical name, a new record, a new combination, and a new variant, so that plant books cannot be put in order up to now (Chung et al, 1987). In Korea, genus Trapa has been classified into 27 units of OTU (Operational Taxonomic Unit) the fruits collected out of 17 different places to look into the variation of the genus Trapa in Korea. By studying the quantitative and qualitative characters of the fruits in the way of numerical taxonomy, Trapa was classified into 5 species and 2 variants according to the characters of the fruits of the living genus Trapa, which Chung et al.(1987) recorded as T. bicornis var. coreanus as a new variant.
This study confirmed that the pericarp was discovered at the peat deposits as fossil plants (Hwang et al, 1997;
Lee, 2005), and classified into 34 units of OTU the 272 Trapa fossils which abundantly appeared for the past one thousand years. This study also integrated the data of morphological analysis on the each formation dating geological environment analysis (4 types of geological environmental region); and numerical taxonomy, and then made a reliable list of appearing species and the table of frequency thereof. Next, this study deduces the variation and genealogy of the genus Trapa by dividing into 4 different characteristic Trapa Zones and 3 Groups according to geological environment change.
The purpose of this study is not only to provide comparative data with the present fruit by analyzing the past genus Trapa being first discovered by various periods, but to review the systematically unsorted genus Trapa in the East Asia and to be an help to hierarchy and lineage evolution for the genus Trapa by setting up accurate classification classes and classification limits.
MATERIALS AND METHODS
Plant materials collected by the researchers of Korea Institute of Geoscience and Mineral Resource at peat deposits around Pyeongtaek-City in Kyeonggi-Province from October 2005 to July 2006, were used, and we performed trench excavation surveys at HS and HY with a forklift (Figure 1). We made long trenches measuring 300 m × 2 m east and west along the Daeban-river east, named them A-1 and A-2 in the direction of north and south centering around the spots where the peat deposits were confirmed, and took samples from the 2.075~5.175-
meter-high and 3.100-meter thick peat deposits. At Hwangsan-ri in Hyungduk-myeun, we made extended trenches measuring 250 m × 2 m, named them B-1 and B-2 in the direction of north an south, and took samples from 3.912~5.412-meter-high and 1.500-meter- thick peat deposits. In the area B, samples were divided into 10 samples ranging from No.1 to No. 10, and collected 500 g on each layer. However, seeds of the genus Trapa could not be collected from this area.
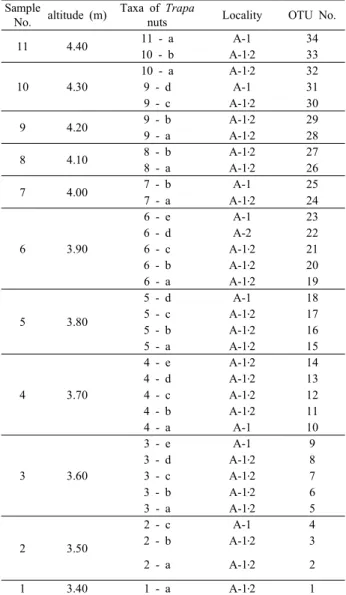
We chose the area A-1 and A-2, and collected the seeds
Sample
No. altitude (m) Taxa of Trapa
nuts Locality OTU No.
11 4.40 11 - a A-1 34
10 - b A-1․2 33
10 4.30
10 - a A-1․2 32
9 - d A-1 31
9 - c A-1․2 30
9 4.20 9 - b A-1․2 29
9 - a A-1․2 28
8 4.10 8 - b A-1․2 27
8 - a A-1․2 26
7 4.00 7 - b A-1 25
7 - a A-1․2 24
6 3.90
6 - e A-1 23
6 - d A-2 22
6 - c A-1․2 21
6 - b A-1․2 20
6 - a A-1․2 19
5 3.80
5 - d A-1 18
5 - c A-1․2 17
5 - b A-1․2 16
5 - a A-1․2 15
4 3.70
4 - e A-1․2 14
4 - d A-1․2 13
4 - c A-1․2 12
4 - b A-1․2 11
4 - a A-1 10
3 3.60
3 - e A-1 9
3 - d A-1․2 8
3 - c A-1․2 7
3 - b A-1․2 6
3 - a A-1․2 5
2 3.50
2 - c A-1 4
2 - b A-1․2 3
2 - a A-1․2 2
1 3.40 1 - a A-1․2 1
Table 2. Materials and collection data of Trapa nuts in Korea which were used in morphological and numerical analysis
of the genus Trapa at the height above sea level of 3.415~4.435 meter. In the area A-1, we collected 11 samples ranging from sample No.1 to sample No.11 (Table 2). The genus Trapa was discovered at intervals of 10㎝
at every 22 peat samples. In the area A-2, the 10 samples ranging from sample No.1 to No.10 were able to be collected at the same height as the one in the area A-1, and there were discovered the genus Trapa in the samples except for in the sample No. 11. The collected samples were delivered kept air-tight lest it should boil away and shrink. They were also kept refrigerated and divided by the 2nd step of Standard Water Sieve (2.0 ㎜, 0.5 ㎜). The divided genus Trapa and seeds were fixed in FAA
solution.
1. External morphological research
External morphological characters of Trapa nuts were preserved in its shape, and was measured by observing the collected samples of each taxon through a dissecting microscope. The terms being used for the external morphological description made up for the various morphological characters newly discovered so that it offered the new classification standard.
2. A numerical taxonomic research
For studies of numerical taxonomy, the collected taxa were classified into 34 OTUs based on the morphological characteristics and the average was obtained. 12 quantitative and 9 qualitative morphological characteristics were examined. By using the 21 characteristics, we created a data matrix to perform cluster and factor analyses of the morphological characteristics. From the analyses results, we found the similarity and communality of the principal components of the genus Trapa, which were used as basic data to assess the similarity among species and phylogenetic evolution.
Statistical analysis of the results was performed using Statistical Package for the Social Sciences (SPSS; ver.
12.0) program and IBM compatible PC.
3. Geologic research
Research on Trapa was conducted with Korea Institute of Geoscience and Mineral Resource, Quaternary Environment Research Team at the same area from October 2005 to February 2006. This study was focused on the Trapa and adaptational environment and variation of the Trapa was analyzed according to the origin of organic matter, indraft of magnetic mineral, distribution of diatom and pollen through Sedimentologic analysis, Organic Geochemical analysis (TC, TOC, TN, TS), Componential analysis of principal component and trace element, Paleomagnetic analysis, Paleontologic (pollen) analysis, Paleontologic (diatom) analysis, Geochronologic (AMS 14C) analysis (Jin et al., 2005. Study on Earth's Environmental Change using Quaternary Sedimentary
72 Jin-Hee Lee, Dae-Jae Kim Korean J. Environ. Ecol. 29(1) 2015
a
b g
e d h
l
c i
j f
k A
lower calyx
calyx stripes lower
horns stripes
calyx
tube calyx
tube hair upper
horns
tubercle
upper horns stripes lower
horns B
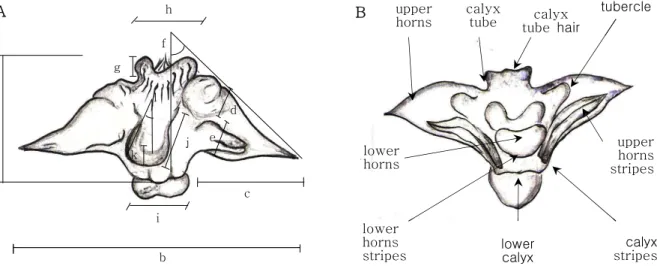
Figure 2. A: The qualitative characters of Trapa nut (The neutral attributes were also included into classification standards, and then more detailed data matrix was made and used as a preliminary data for a numerical taxonomic research)B: The quantitative characters of Trapa nut (Adopted and modified from Staszkiewicz and Wojcicki, 1979-a: Height of nut with calyx tube b: Width of nut at the surface of upper horns c: Width of upper horns d: Length of upper horns e: Thickness of upper horns f: Inclination of upper horns g: Height of nut calyx tube h: Width of nut calyx tube i: Width of lower horns j: Length of lower horns k:
Thickness of lower horns l: Inclination of lower horns)
No. Characters Character state
1 Calyx tube shape 0=not extend, 1=trapezoid, 2=vertically-risen, 3=extension
2 Calyx tube hair 0=absent or short, 1=present
3 Lower calyx shape 1=horizontal, 2=trapezoid, 3=inverted triangle, 4=tetragonal
4 Calyx stripes 0=absent, 1=present 5 Existence of tubercle 0=absent, 1=present
6 Upper horns shape 0=absent, 1=acicular, 2=conoid, 3=rectilineal, 4=semicircular, 5=circular
7 Upper horns stripes 0=absent, 1=present
8 Lower horns shape 0=absent, 1=vestiges, 2=acicular, 3=tubercle, 4=thickish, 5=semicircular, 6=round
9 Lower horns stripes 0=absent, 1=present
Table 3. Qualitative characters and character states for cluster analysis by morphological study of Trapa*
Records: Phase 1. Korea Institute of Geoscience and Mineral Resource research reports OAA2004002-2005(2), Ministry of science & Technology. 267 p.).
RESULTS AND DISCUSSION
1. External morphological research by period.
To take a measurement of external morphological characters of the genus Trapa, the nuts were separately measured by their quantitative characters (Figure 3) and qualitative characters (Figure 4), such as the forms of Trapa nuts, the shape of lower calyx, size of fruits, the shape of horn, stripe, size, angle, attached tubercle, size of each organ, and etc.
Following character analysis, we were able to draw the following conclusion from the analysis results of qualitative characters and quantitative characters for Trapa nuts. The shape and size of the main organs of the Trapa nut are predicted to be closely associated with mutation.
They have important roles in determining species differentiation (Robert, 1999). Consequently, the change in the overall shape and size of the Trapa nut is one of the important aspects of morphological evolution of the genus Trapa. Although a certain overlap exists among the
different species, the morphological differentiation of the species plays an important role in differentiating the genus Trapa. While the morphological characteristics are closely associated with ecological factors, certain morphologies are not clearly defined or exhibit neutral attributes.
Consequently, the overall shape of the Trapa nut as well as the neutral attributes of the horns, lower calyx, and stripes were included in the classification standards to create a detailed data matrix to be used in numerical taxonomic analyses (Figure 2, Table 3).
Trapa nut
Quantitative characters Qualitative characters
A B C D E F G H I J K L 1 2 3 4 5 6 7 8 9
11 a 1.20 2.10 0.6 0.22 0.22 80 0.15 0.25 - - - - 2 1 3 1 0 2 0 1 0
10 b 1.20 2.30 0.6 0.4 0.3 93 0.07 0.4 - - - - 1 0 3 1 0 2 1 1 0
a 0.90 2.50 0.9 0.6 0.5 70 0.1 0.27 - - - - 1 1 2 0 1 4 1 1 0
9
d 0.90 2.80 1.15 0.7 0.6 118 0.15 0.4 - - - - 1 1 1 1 1 4 1 1 0
c 0.65 2.40 1.85 0.3 0.2 82 0.17 0.25 - - - - 2 1 2 1 1 3 1 0 0
b 1.60 3.00 1.1 1.1 1.0 92 0.08 0.5 - - - - 1 1 4 1 1 5 1 1 0
a 1.30 1.50 - - - - - - - - - - 0 0 3 0 0 0 1 0 0
8 b 1.10 2.30 0.8 0.75 0.5 112 0.12 0.25 - - - - 1 1 4 1 1 4 1 1 0
a 0.70 2.90 1.3 0.7 0.6 93 0.08 0.3 - - - - 1 1 1 1 1 5 1 1 0
7 b 1.90 2.30 0.3 0.15 0.15 46 0.17 0.3 - - - - 1 1 3 1 1 2 1 1 0
a 1.60 3.50 1.45 0.45 0.82 96 0.45 0.9 0.72 1.6 0.35 113 3 1 2 1 0 3 1 4 0
6
e 1.25 2.30 0.7 0.6 0.55 113 0.1 0.45 - - - - 1 1 4 1 1 4 0 1 1
d 1.70 4.10 1.6 0.75 0.95 128 0.55 1.45 0.52 0.55 0.32 125 3 1 2 1 0 4 1 6 0
c 1.15 5.20 2.2 1.1 1.2 72 0.12 0.5 0.62 0.8 0.6 100 1 1 4 1 1 4 1 6 0
b 0.80 2.30 1.05 0.1 0.1 110 0.37 0.12 - - - - 2 1 3 0 0 1 1 0 0
a 1.30 3.95 1.9 0.45 0.87 92 0.2 0.9 1.2 1.55 0.4 110 1 0 2 1 0 4 1 5 1
5
d 2.30 4.55 1.2 0.7 0.95 92 0.5 0.97 - - - - 3 1 2 1 1 3 1 1 0
c 2.25 5.17 2.1 0.8 1.3 87 0.45 0.95 1.05 0.65 0.6 115 3 1 2 1 1 4 1 5 0
b 1.60 2.90 0.7 0.25 0.25 81 0.17 0.5 - - - - 2 1 2 0 1 2 1 1 0
a 1.35 3.30 1.3 0.5 0.7 102 0.32 0.55 - - - - 2 1 2 0 1 3 1 1 0
4
e 2.20 4.10 1.6 0.75 0.9 74 0.3 0.8 0.8 0.55 0.35 125 3 1 2 1 1 3 1 6 0
d 1.40 3.50 1.35 0.9 0.85 104 0.1 0.5 - - - - 1 1 2 1 1 4 1 3 0
c 1.17 3.10 1.1 0.7 0.65 106 0.1 0.4 - - - - 1 1 2 0 1 4 1 1 0
b 1.30 2.70 0.65 0.55 0.55 103 0.17 0.62 - - - - 1 1 2 1 1 2 1 1 0
a 1.50 3.20 1.45 0.85 0.85 92 0.12 0.35 - - - - 1 0 2 1 1 4 1 1 0
3
e 1.60 3.27 1.25 0.15 0.15 114 0.4 0.27 0.17 1.2 0.17 102 2 1 3 1 0 1 1 2 0
d 1.70 0.85 0.6 0.08 0.08 87 0.1 0.17 0.05 0.75 0.07 139 2 1 3 1 0 1 1 2 0
c 1.10 2.90 1.25 0.6 0.6 93 0.1 0.32 - - - - 1 1 2 0 1 3 1 1 0
b 1.55 2.50 0.75 0.8 0.7 86 0.15 0.52 - - - - 2 1 4 1 1 3 1 1 0
a 1.70 2.90 1.2 0.65 0.5 94 0.15 0.57 - - - - 2 1 2 1 1 3 1 1 0
2
c 1.35 2.50 0.95 0.95 0.8 104 0.1 0.52 - - - - 1 1 2 1 1 4 1 1 0
b 2.10 4.40 1.6 0.95 1.1 105 0.42 1.2 0.65 0.4 0.15 144 3 1 2 1 1 3 1 5 0
a 1.90 4.10 1.15 0.75 0.75 83 0.22 0.65 - - - - 3 1 2 1 1 4 1 1 0
1 a 1.65 2.85 0.95 0.9 0.85 96 0.17 0.65 - - - - 1 1 4 1 1 4 1 1 0
* The sorting standard was added including appearance, hone, calyx tube, lower calyx, and stripes for more detailed data matrix
** Nuts were separately measured by their 12 Quantitative characters and 9 Qualitative characters, such as the forms of Trapa nuts, the shape of lower calyx, size of nuts, the shape of hone, stripe, size, angle, attached tubercle, size of each organ, and etc.
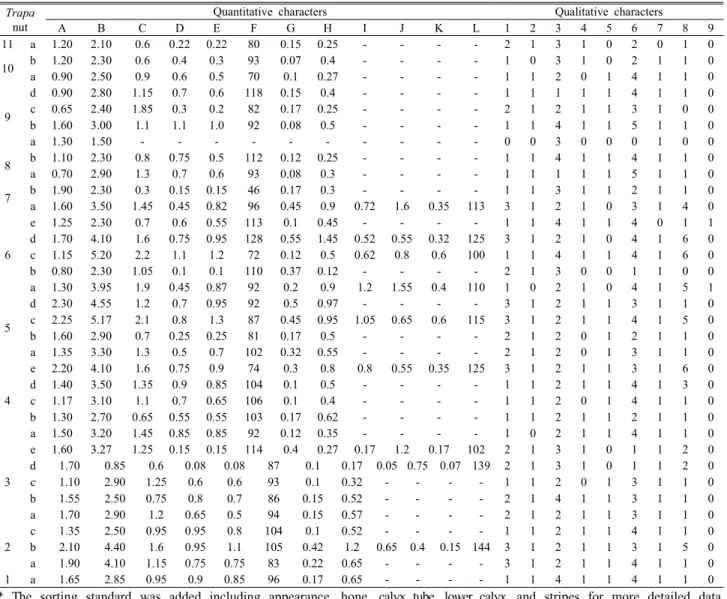
Table 4. Morphology characters of Trapa nut used for the principal component analysis**
2. Numerical taxonomic research
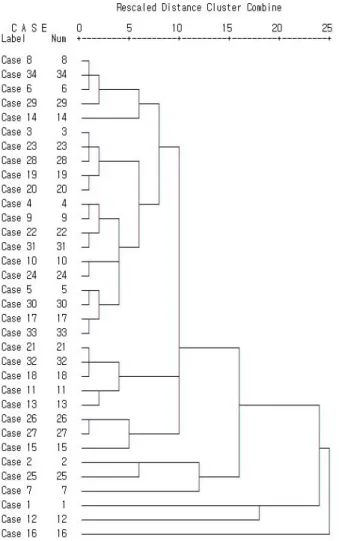
1) Cluster analysis
To classify the Trapa of Korean origin, we performed cluster analysis on the 21 external morphological data (9 qualitative and 12 quantitative characteristics) to draw a dendrogram. The dendrogram for the 21 external morphological characteristics were identical. Cases 11 and 16, 13 and 32, and 18 and 21 exhibited high similarity, thereby forming clusters, respectively. The aforementioned
clusters were placed below a big cluster, which was formed by clustering all the aforementioned clusters with Case 14. Further, the cluster size increased when this big cluster was grouped with Case 7, which marked the final stage of phylogeny.
The following clusters were observed in the dendrogram of 9 external morphological qualitative characteristics (Figure 4): Cases 21, 32, and 18 formed identical clusters with quantitative characteristics; Cases 11 and 13, as well as Cases 26 and 27, formed identical clusters. However,
74 Jin-Hee Lee, Dae-Jae Kim Korean J. Environ. Ecol. 29(1) 2015
Figure 3. Cluster dendrogram created using 12 quantitative characteristics and 21 external morphological data
Figure 4. Dendrogram of 9 morphological qualitative characteristics
cases 14 and 16 formed a cluster by combining with another 4. Consequently, classification based on qualitative characteristics yielded similar classification results as that by using the quantitative data. On the basis of the similarity of the two classification results, the overall differentiation elicited by the cluster analysis can be interpreted to show consistency.
2) Factor Analysis
We extracted 7 factors after factor analysis by using 12 quantitative and 9 qualitative external morphological characteristics. The total explanatory power of factor analysis was 81 %. Thus, the 21 morphological characteristics were reduced to 7 major factors that may have contributed to the formation of clusters. Quantitative characteristics 1, 2, 3, 7, and 8 and qualitative characteristics 1 and 8 formed Factor 1, while quantitative characteristics 4 and 5 and qualitative characteristics 5 and 6 formed Factor 2.
Quantitative characteristics 10 and 11 formed Factor 3.
The quantitative characteristic 12th exhibited high negative loading to Factor 3. The qualitative characteristic 9 formed Factor 4, while qualitative characteristic 7 exhibited high negative loading to Factor 4. The qualitative characteristic 2 formed Factor 5, quantitative characteristic 6 formed Factor 6, and qualitative characteristics 3 and 4 formed Factor 7. The distribution of the 34 Trapa species with respect to Factors 1 and 2 is shown below.
Figure 5. Distribution of the genus Trapa based on Factors 1 and 2
Trapa Zone
Alt.
(m)
14C age Appearance by period of the genus Trapa of Korea
Ⅳ
4.4 4.3 4.2 4.1 4.0 3.9 3.8 3.7 3.6 3.5 3.4
5560±40
1-a 3-a 4-a 5-a 9-a 10-a
11-a
3-b 4-b 5-b
7-a 7-b
8-a 8-b
9-b 10-b
2-a 2-b 2-c
3-c 4-c
5-c
9-c 9-d
3-d 3-e
4-d 4-e
5-d
6-a 6-b 6-c 6-d 6-e
5800±40
Ⅲ
6160±40
Ⅱ
6560±60
Ⅰ
Figure 6. Appearance by period of the genus Trapa of Korea (34 species of the Trapa was sorted and the period was measured by depending on radiocarbon dating from 21 samples. Each species size is in proportion to the real species)
3. Appearance and frequency of the genus Trapa of Korea Appearance map of the genus Trapa of Korea was made by sorting 34 species of the Trapa seed appearing in 21 samples collected at peat deposits and dating by layer.
4. Lineage evolution of Trapa of Korea in accordance with period and environmental change
In order to examine variation of Trapa nut of Korea, Trapa nut appearing at peat deposits was sorted into 34 units of OTU. And then, by making integrated analysis
76 Jin-Hee Lee, Dae-Jae Kim Korean J. Environ. Ecol. 29(1) 2015
of adaptational environment of plant through morphological analysis on the each formation, dating, numerical taxonomy, and geological analysis data (4 types of geological environmental region), the genus Trapa of Korea was sorted into 4 different Trapa periods and then lineage evolution was presumed. Be noted that, although 1-a species appeared at all layers and was supposed to have a variety of variation, it lacked evidence to prove that it should be origin of all species. Therefore, like 3-d, the possibility cannot be ruled out which other species from the outside should come into the flow of linear evolution.
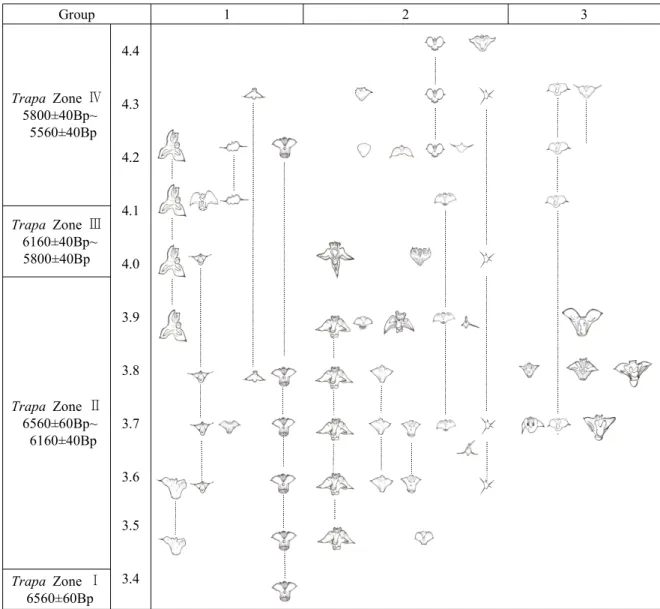
In order to minimize the potential error in linear classification, a reliable list of appearing species (Figure 6) and the table of frequency thereof was made by integrating dating data, detailed external morphological analysis data, geological environment analysis data, and numerical taxonomic data. 4 different Trapa zones were sorted by integrating all of the data above, and a distribution diagram was made (Figure 7).
1) Trapa Zone I: before 6560±60Bp
Total 4 species and 24 individuals were discovered.
Seawater penetrated this zone, and so the most drastic changes happened a lot. So, reaching up to 6560 ± 60 Bp, this zone was presumed gradual transition to a tranquil sedimentary environment. Geological analytic data showed that, before 6560 ± 60 Bp, only 1-a species, assumed to be dominant species and keystone species, appeared, and the number of species was limited due to seawater penetration. Thereafter, changing to tranquil environment, relatively large lower calyx stood out and stripe got distinct.
2-a Species was similar to T. bispinosa classified by T. Nakai (1942) and Ding and Fang (1991) of China. As such, we expected that the 2-a species had existed in South Korea and would have also existed in Japan and China.
In addition, cluster and factor analyses on 2-a and 2-c species showed that they belong to 1Group as allied species. On the other hand, 2-b species belonged to 2Group with 2-c species. In particular, 2-b is estimated to have thrived until 6200 ± 80 Bp, when the arrival of Trapa Zone III led to extinction or mutation of the species because of shallow water depth and the rapidly changing environment of the Zone. Consequently, similar to the 2-b species that thrived in sites with deep water, large species
that exhibit relatively less pointed horns are considered to have had difficulties acclimating to the rapidly changing environment unlike the 3-d species.
2) Trapa Zone II: 6560±60 Bp~6160±40 Bp In this period, there were discovered a variety of species and plenty of Trapa fossils, which totaled 22 species and 128 individuals including newly discovered 19 species. A diatom living in the deep lake mainly and NAP(Trapa, Typha, Nelumbo, Nuphar) appeared then. According to the principal component analysis, plankton living in the environment of lake or marsh, or organism originated from a diatom appeared. So, the environment would be interpreted as deoxidation environment of deep and abundant water. For approximately 400 years, many Trapa species irrespective of their size and appearance of the horn, calyx, and tubercle, flourished in deep water and relatively quiet environments. It is assumed that a variety of mutations occurred during this period. In particular, the majority of species that had thrived during this period, including 2-b, 3-a, 3-d, 4-a, 4-e, 5-b, 5-c, 6-a, 6-c, and 6-d, have not been reported as currently existing, not only in South Korea but also in Japan. In particular, Trapa species such as 3-d and 6-b were shown to be similar to Trapa americana Knowlton of Japan and classified as T.
incisa by Ding and Fang (1991) of China. In addition, 3-d is also assumed to be the ancestor of T. incisa referred to by Chung et al. (1987) in South Korea. Further, 4-d is similar to Trapa platycerata reported by Miki (1953) and assumed to not exist presently. However, 4-d was shown to have continued to exist during the tumultuous and rapidly changing environment of Trapa Zone III from 6560 ± 60 Bp to 5560 ± 40 Bp. Further, while T. acornis (Nakano, 1964) that commonly appear in Chinese and Japanese even today have not been reported in South Korea, the present study showed for the first time that T.
acornis (Nakano, 1964) had existed in South Korea in the past, as 7-b and 9-a.
3) Trapa Zone III : 6160±40Bp~5800±40Bp There were appeared total 9 species and 34 individuals including newly discovered 4 species, and there were a lot extinct species (3-c, 4-c, 6-b, 6-e, and 8-b). Especially during this period, sea level was frequently in flood according to organic physical geographic analysis and pollen
Group 1 2 3
Trapa Zone Ⅳ 5800±40Bp~
5560±40Bp 4.4
4.3
4.2
4.1 Trapa Zone Ⅲ
6160±40Bp~
5800±40Bp 4.0
Trapa Zone Ⅱ 6560±60Bp~
6160±40Bp 3.9
3.8
3.7
3.6
3.5
Trapa Zone Ⅰ 3.4 6560±60Bp
Figure 7. Assumed evolutionary lineage adaptation of the genus Trapa of Korea (The following picture was made by integrating the C14 carbon dating data, External morphological analysis data, numerical taxonomic data which is from Cluster Analysis and Factor Analysis, Geologic data and the frequency of the genus Trapa of Korea)
analysis(increase in quantity of pollen-Cyclobalanopsis, Lepidobalanus) and this zone turn to deoxidation environment featuring shallow water depth. So, the variation of Trapa nut and its population rapidly decreased. Because the relatively large species was extinct, small-sized and horizontal Trapa became dominant.
Damage from sea water supposedly affected the growth of plants so that the mean size of Trapa nut became smaller than the one of allied plants. Besides, the small-size and horizontal Trapa like 8-a species became dominant. That is because it occupied ponds under the
circumstance of radical change or shallow water depth, and particularly its long and thin end of hone could readily take root in the bottom of water. While the majority of the large Trapa species that had flourished in Trapa Zone I and II became extinct during this period, the large species 6-a survived in Trapa Zone III because of its large size among the 34 species and the four strongly developed horns that aid in occupy the water base, thereby ensuring survival in the rapidly changing environment. In particular, the existence of 6-a species in the past and present had never been reported, along with 2-b, 4-e, 7-a, 7-b, 8-a,
78 Jin-Hee Lee, Dae-Jae Kim Korean J. Environ. Ecol. 29(1) 2015
8-b, 9-b, and 9-d; 6-a species is estimated to have only appeared in South Korea
4) Trapa Zone Ⅳ: 5800±40Bp~5560±40Bp There were appeared total 10 species and 79 individuals including newly discovered 6 species. In Trapa Zone IV, the variation again got diverse and activated, but, due to deoxidation environment (featuring rapid decrease in quantity of pollen-Alnus, Salix, Trapa, Typha and yielding of melting scraps and dormant spores by diatom); decline in water depth; and exposure of the surface of the earth, the weather changed to become dry and cold so that a variety of variation occurred; overall, the size of Trapa nut became smallest; the horizontal Trapa nut remained dominant.
In 5,560±40Bp, only 3 specie and 7 individuals including a new species appeared. Sediment, accumulated when seal level reached its peak, stagnated water flow, and plenty of organisms much accumulated which supposedly led to decline in water depth and exposure of the surface of the earth. The genus Trapa currently existing in South Korea is estimated to be 3-d of 2Group as well as 10-a and 11-a.
Other species are estimated to not have survived to the present day. Further, 11-a of 2Group is estimated to be the ancestor of T. bicornis var. coreanus reported by Chung et al. (1987) of Korea. In particular, 10-a of 3Group is assumed to be the Trapa species reported by Nakano (1964) of Japan. 10-a is also considered to be an ancestor of T. japonica and is easily found in all parts of South Korea.
Thus, by using the basic external morphological data, numerical taxonomy analysis, and geological data, we classified the genus Trapa by period and group by using cluster and factor analyses.
We reviewed the boundaries among taxa through the key to species of the genus Trapa of Korea and appearance map of the genus Trapa of Korea by sorting the genus Trapa of Korea, being newly discovered at peat deposits, into 34 units of OUT, and making principal component analysis and cluster analysis with detailed external morphological characters. Also, we prudently presumed the linear evolution, and discussed the adaptational environment of the genus Trapa by period by making use of various geologic analytic data. Until now, the study in the genus Trapa has been limited to systematic
classification of the existing Trapa of Japanese and Chinese, and it is not too much say that there has been no study in the past species. Only in Japan (Miki, S. 1952;
Miki, S. 1953), the systematic classification of the past genus Trapa was tried, but had its limitation about the accuracy of species classification and systematic classification due to lack of information on the exact period when the species lived and scientific approach. New variants from all of the world, including T. litwinowii V. vassil. var.
chihuensis S.F.Guan & Q.Lang of China (Guan and Lang 1987), have been reported. However, in order to determine the adaptation of the existing family Trapaceae, the information on the past species are in need.
Xinog, Huang and Wang (1990) insisted that the change of the (form) size is one way of evolution of the Trapa, and Ding and Fang (1991) insisted in his thesis of "Study on the pollen morphology of the Trapa from Zhejing" that the process of evolution of the fruit is the degeneration the rectangle corners, and the disappearance of the splinters and tubercle lengths through observation of the solid figure. Study on the Trapa in China (Ding and Fang 1991) and Japan (Nakano, 1913; Nakano, 1964) is based on degeneration of the angles or tubercle lengths. But it is not sure that the Trapa can be analyzed primordial character or differential character through the degeneration because the Trapa has a variety of variation so it is required that studying on the Trapa be based on geological research data from researching the variation of Trapa nut.
In Korea, genus Trapa has been classified into 5 species and 2 variants according to the characters of the fruits of the living genus Trapa (Chung et al., 1987).
Therefore, through Multivariate Analysis it is assumed that Trapa that exists in Korea is similar to 3-d 10-a 10-b 11-a.
About 30 types of Trapa nuts in the past are proved to be different in external morphology. It is considered to be meaningful to establish a new classification standard and apply for new variants systematics worldwide with this study result. Most of the studies on the evolution of the Trapa emphasize the importance of combining the data from many types studies on the Trapa. This study is expected to classify 34 species of Trapa which have been found in the last 1000 years, so it is hoped that this will allow for understanding the systematic adaptational environment and lineage of the Trapa.
REFERENCES
Choi, H.K. (1986) A Monograph of Vascular Hydrophytes in Korea. Ph.D. Dissertation at Seoul National University. 272pp.
(in Korean)
Cook, C.D.K., BJ. Gut, EM. Rix, J. Schneller and M. Seitz. (1974) Water Plants of the World. Dr. W. Junk Publishers, The Hague, 561pp.
Cronquist, A. (1981) An integrated system of classification of flow- ering plants. Columbia Univ. Press, New York, 1262pp Chung, Y.H., H.K. Choi, K.H. Suh and H.C. Shin. (1987)
Numerical taxonomic study of the nuts of genus Trapa in korea.
J Plant Tax. 17(1): 45-54. (in Korean with English abstract) Ding, B.Y. and Y. Fang. (1991) Study on the pollen morphology of
Trapa from Zhejing. J Acta phytotaxonomica Sinica 29(2):
172-177. (in Chinese with English abstract)
Guan, S.F. and Q. Lang. (1987) New species of Trapa and Najas from Jiangxi, China. Chin Sci bull 7(1) 77-79. (in Chinese) Hwang, S.I., S.O. Yoon and W.R. Jo. (1997) The Change of the
Depositional Environment on Dodaecheon River Basin during the Middle Holocene. Geol 32(4): 403-420. (in Korean with English abstract)
Jin, J.W., K.H. Min and J.H. Jang. (2005) Study on Earth's
Environmental Change using Quaternary Sedimentary Records: Phase 1. Korea Institute of Geoscience and Mineral Resource research reports OAA2004002-2005(2), Ministry of science & Technology. 267. (in Korean)
Lee, J.H. (2005) Plant Macrofossils in Peatland of Pyeong-Taek Area. Kor Wetlands Society 312-321. (in Korean with English abstract)
Miki, S. (1952) Trapa of Japan with special reference to its remains.
J Inst Polytech Osaka City Univ. 3: 1-29. (in Japanese) Miki, S. (1953) Morphological character of remains of Trapa and
adaptation of aquatic life. Jap J Ecol. 2: 111-116. (in Japanese) Nakai, T. (1942) Notulae ad Plantae Asiae Orientalis. Jap J Bot
18(8): 421-437. (in Japanese with English abstract)
Nakano, H. (1913) Beitrage zur Ketnis der variation von Trapa in Japan. J Jap Bot 50: 440-458. (in Japanese)
Nakano, H. (1964) Further Studies on Trapa from Japan and its Adjacent Countries. Jap Bot Mag Tokyo 77: 159-167.
Robert, W. (1999) Natural Enemies of Trapa spp. in Northeast Asia and Europe. J Biol Control 14: 168-180.
Staszkiewicz, J. and JJ. Wokcicki. (1979) Biometrical analysis of Trapa L. nuts from Poland. Fragmenta Floristica et Geobotanica Ann. 25: 33-63.
Xiong, Z.Y., D. Huang and W. Wang. (1990) Numerical taxonomic studies in Trapa in hubei. J Wuhan bot 8(1): 47-52. (in Chinese)
80 Jin-Hee Lee, Dae-Jae Kim Korean J. Environ. Ecol. 29(1) 2015
Factors
1 2 3
Quant8 0.862 0.264 -0.085
Quant7 0.836 -0.282 0.099
Quali1 0.791 -0.194 -0.158
Quali8 0.789 0.213 0.114
Quant2 0.743 0.434 0.394
Quant1 0.737 0.035 -0.331
Quant3 0.562 0.339 0.456
Quant4 0.174 0.91 0.059
Quali6 0.046 0.859 0.113
Quant5 0.568 0.735 0.167
Quali5 -0.189 0.732 -0.047
Quant9 0.244 0.398 0.346
Quant12 0.011 -0.105 -0.897
Quant11 0.131 0.434 0.609
Quant10 -0.145 -0.326 0.517
Quali9 0.021 0.051 0.127
Quali7 0.142 0.113 0.046
Quali2 0.069 0.107 -0.06
Quant6 0.074 0.073 -0.016
Quali3 -0.19 -0.075 0.115
Quali4 0.341 0.233 -0.131
Eigenvalue 4.772 3.737 2.189
Explanatory
Power 22.723 17.797 10.425
Total
Explanatory Power 22.723 40.52 50.945
Factor Extraction Method: Principal Component Analysis.
Rotation Method: Varimax with Kaiser normalization
Appendix 1. Factor analysis result of 12 quantitative and 9 qualitative characteristics