세포벽 (1,3)-β- D -Glucan Polymer 합성의 저해로 인한 황금(Scutellaria baicalensis)의 항바이오필름 활성
김연희*
세명대학교한의학과
Received : September 25, 2012 / Revised : October 19, 2012 / Accepted : November 2, 2012
서 론
Candida species는기회감염균으로치명적인질환에걸
렸거나면역력이약화된환자에게표재성전신질환을유발 한다[23]. 비록 C. krusei, C. tropicalis, C. glabrata, C.
parapsilosis, C. guilliermondii와같은 non-albicans species 가지속적으로발견되고있으나, C. albicans는아직도가장 널리발견되는진균성병원균으로기록되고있다[15].
C. albicans의독성은 adhesin을포함한숙주조직에대한 부착분자, 단세포성효모형과필라멘트성균사형으로의상 호 형태변화, 아스파르트산 단백질 분해효소나 phos-
pholipase와같은가수분해효소의분비, 환경변화에대한반
응성과 같은 다양한 요인에 의해 결정된다[4, 10]. C.
albicans의가장중요한독성요인중하나는생물학적표면
과맥관내카테타를포함한비생물학적표면모두에바이오 필름이라는소군락을형성하여[8, 24] 숙주를감염시키는능 력이다. 최근의연구에따르면 C. albicans에의해형성된대 부분의질환이바이오필름의성장과관련이있음을보여준
다[2]. 진균성바이오필름은캔디다의은신처로작용하여숙
주의면역체계를회피하고다른병원체로부터의경쟁적압 력을견뎌내며항진균제의치료에대한내성을유발하여[5]
전통적인항진균제사용에대한문제점을심화시키고있다. 진균의세포벽은세포건조중량의 15-30%를차지하는데, 이 는세포의상당한에너지를세포벽합성에사용함을의미한
다[13]. 세포벽은강한팽압을견딜수있게하며세포내의
안정된삼투압을유지할수있게한다. 또한세포형태를유 지시키고형태변화를가능하게하며물리적인손상에대한 방어를돕는다. 이밖에세포벽은부착이나건조에대한보호 기능, 세포벽의외피단백질에대한부착장소로작용한다[13].
C. albicans의 세포벽은 mannoprotein(16-23%), (1,3)-과 (1,6)-β-D-glucan(47-60%), 단백질(16-25%), 지질(1-7%), 키
틴(0.6-9%)으로조성된다. 이들중글루칸은세포의형태와
Antibiofilm Activity of Scutellaria baicalensis through the Inhibition of Synthesis of the Cell Wall (1, 3)-β-D-Glucan Polymer. Kim, Younhee*. Department of Oriental Medicine, Semyung University, Jecheon, Chungbuk 390-711, Korea Candida biofilms are self-organized microbial communities growing on the surfaces of host tissues and medical devices. These biofilms have been displaying increasing resistance against conventional antifungal agents. The roots of Scutellaria baicalensis have been widely used for medicinal purpose throughout East Asia. The aim of the present study was to evaluate the effect of S. baicalensis aqueous extract upon the preformed biofilms of 10 clinical C. albicans isolates, and assess the mechanism of the antibiofilm activity. Its effect on preformed biofilm was judged using an XTT reduction assay and the metabolic activity of all tested strains were reduced (57.7 ± 17.3%) at MIC values. The S. baicalenis extract inhibited (1, 3)-β-D-glucan synthase activ- ity. The effect of S. baicalensis on the morphology of C. albicans was related to the changes in growth caused by inhibiting glu- can synthesis; most cells were round and swollen, and cell walls were densely stained or ruptured. The anticandidal activity was fungicidal, and the extract also arrested C. albicans cells at G0/G1. The data suggest that S. baicalensis has multiple fatal effects on target fungi, which ultimately result in cell wall disruption and killing by inhibiting (1, 3)-β-D-glucan synthesis. There- fore, S. baicalensis holds great promise for use in treating and eliminating biofilm-associated Candida infections.
Keywords: Antifungal activity, biofilm, Candida albicans, cell wall, (1,3)-β-D-glucan synthase, Scutellaria baicalensis
*Corresponding author
Tel: +82-43-649-1346, Fax: +82-43-649-1702 E-mail: [email protected]
경도를유지하는구조적중요성을나타낸다[1, 3]. (1,3)-β-D- glucan synthase는 Candida 속과 Aspergillus 속을포함하 는진균류의세포벽중한성분인 (1,3)-β-glucan polymer의 형성을 촉매하는 효소로 C. albicans, Neurospora crassa, Saccharomyces cerevisiae와같은효모의세포막에결합되어 발견된다[1, 7, 28]. 이효소는 UDP-glucose를기질로사용 하여 직선형의 polymer 형성을 촉매하며 caspofungin, anidulafungin, micafungin과같은항진균제의표적으로작 용한다[16].
Candida 감염에대한치료는진균의진핵세포적특성때
문에쉽지않다. 지난수십년간진균의치료는진균세포막 의 ergosterol에결합하는 amphotericin B와같은살진균성
polyene에 의존해왔다. 최근에는 비교적 사용이 용이한
azoles 계열의정진균제가진균의감염에대한치료제로널
리사용되고있다[21]. 그러나제한된약제의지속적인사용
으로인하여내성균주가출현하고일부환자에서는치료량 에서의심각한부작용과관련된다양한문제들이발생하고
있다[18]. 따라서독성이낮고높은치료효과를지닌고유한
항진균제의개발이시급한실정이다.
최근보편적인미생물통제전략에대한대체요법으로다 양한식물유래화합물에대한관심이집중되고있는데그이 유는이들화합물들이질병과감염에대한치료및예방을위 해오랜역사동안민간요법으로널리사용되어안전하다고 생각되기때문이다[9]. 황금(Scutellaria baicalensis Georgi)은 한국, 중국, 일본, 몽고원산지로수천년동안한의학에서사
용되어왔다. 3-4년된식물의뿌리부분이약재로사용되며주
로열, 감기, 설사, 알레르기성비염, 천식의치료에사용되고 있다. 이식물의주성분은 baicalin, baicalein, wogonin으로메 탄올, 에탄올, 아세톤에잘녹지만물이나벤젠에는잘녹지 않는다. 이들은항염증제, 항산화제로작용한다[12, 29].
본연구는 S. baicalensis의수용성추출물이캔디다증환 자들로부터분리된임상 Candida균주에대해우수한항바 이오필름 능력을 나타내며, 이러한 능력이 C. albicans의 (1,3)-β-D-glucan synthase 활성의저해를통한세포벽합성 억제와이로인한살진균활성그리고 G0/G1에서 S로의세 포주기의진행을억제하는다중효과와관련이있음을 XTT 환원분석법, (1,3)-β-D-glucan synthase 활성, FUN-1/Calcofluor white 이중염색, neutral red 염색, time-kill assay, 세포주기 분석을통해연구하였다.
재료 및 방법
식물추출물
건조된황금(S. baicalensis)의뿌리를 jchanbang.com에서 구매하였다. 30 g의황금을 600 ml의증류수에넣어 1시간
동안불린후 2시간동안끓였다. 2,000 g로 20분간원심분 리한후상등액을진공농축기로농축시키고동결건조하여 황금의수용성추출물 14.1 g을얻었다. 수용성추출물(이하 S. baicalensis)은 50 mg/ml의 농도로 증류수에 녹인 후 0.45µm filter (Avantec, Japan)로 여과하여 사용 전까지 -20oC에보관하였다.
균주 및 배양
Candida albicans (KCCM 50235, ATCC 18804), C.
glabrata (KCCM 50044, ATCC 2001), C. tropicalis (KCCM 50075, ATCC 750), C. krusei (KCCM 11426, ATCC 32196)는한국미생물보존센터(Korean Culture Center of Microorganisms, KCCM)에서분양받아사용하였다. 바이오 필름실험에사용된 10 C. albicans 균주는캔디다증환자로 부터분리하였으며[22] 분리균주는−70oC에보관하였고필요 시 YM agar (Becton Dickinson, USA) 또는 YM 액체배지에 서계대배양후사용하였다.
최소 억제농도(Minimum Inhibitory Concentration, MIC) MIC는 CLSI M27-A3 미량희석법[6]에준하여 resazurin 을포함시킨색변화측정법[17]을병행하여사용하였다.
모든분석은 3회반복하였고 MIC는두가지기준에따라 결정하였다: MIC는 3회의분석중최소한 2회일치하거나 3회 중 가장 높은 값으로 결정하였다. 양성 대조군으로 amphotericin B (Sigma, USA)를포함시켰다.
바이오필름 형성과 S. baicalensis의 항바이오필름 활성 측정 캔디다증환자에서분리한 10 C. albicans 균주에대한대 수기의 배양액을 5% glucose가 첨가된 Yeast Nitrogen Base (YNB/Glc, Becton Dickinson, USA)에 최종 농도 1×107CFU/ml로 희석하였다. Flat-bottomed 96 well plate (Becton Dickinson, USA)에희석액을 0.1 ml씩넣고 37oC에서 3시간배양하여 C. albicans가바이오필름을형성 하게하였다. 부유상태의효모를제거하고 YNB/Glc로헹군 후음성대조군을포함하여 S. baicalensis가포함된 0.2 ml 의 YNB/Glc를각 well에넣어초기단계의바이오필름에대 한 S. baicalensis 추출물의저해효과를조사하였다. 습도가 유지되는조건에서 24시간동안 37oC에서반응시킨후바이
오필름의생존력을 XTT 환원분석법[25]에따라측정하였다.
모든실험은 4개씩 3회시행하였다. 데이터는 3회실험중의 하나를나타낸다.
C. albicans의 폴리스티렌 표면 부착에 대한 S. bai- calensis의 영향
대수기의 C. albicans ATCC 18804 배양액을 YNB/Glc에
최종농도 1×107CFU/ml로희석하였다. Flat-bottomed 96 well plate에배양액 0.1 ml을넣고 37oC에서 3시간배양하 여 C. albicans가바이오필름을형성하게하였다. 부유상태 의효모를조심스럽게제거하고 YNB/Glc로헹군후음성대 조군을포함하여 S. baicalensis가포함된 0.2 ml의 YNB/Glc 를각 well에넣어 3시간과 24시간후에폴리스티렌바닥에 부착된 C. albicans를위상차현미경(Olympus, Japan)을통 해분석하였다.
Microsomal membrane의 준비
(1,3)-β-D-glucan synthase의 활성 측정을 위한 효모의 microsomal membrane은 Shedlefzky[27]와 Kurtz[16]의 방법을변형하였으며원심분리이후의과정은 4oC에서행하 였다: 1 L의 YM에 C. albicans ATCC 18804를 접종하여 37oC에서 16시간동안진탕배양하였다. 배양액을 1,900 g 로 원심분리하여 캔디다를 회수한 후 10 ml의 breakage buffer [50 mM Tris.Cl (pH 7.4), 1 mM EGTA, 4 mM DTT, 5µM leupeptin, 20µM PMSF]에 현탁시킨 후 3 ml의 0.5 mm glass bead를 넣어 70초씩 12회 Bead beater (BioSpec, USA)를작동시켜세포를파쇄시켰다. 세포의파 쇄정도는위상차현미경으로관찰하였다. 3,000 g로 10분 간원심분리한후상등액을 100,000 g로 1시간원심분리하 여 membrane 침전물을얻었다. Membrane 침전물은 33%
의 glycerol이 포함된 breakage buffer에 녹인 후 -27oC에 보관하였다. Membrane 침전물의단백질정량은제조사의 방법에따라 Bradford 분석법(Bio Rad, USA)으로측정하 였다.
(1,3)-β-D-glucan synthase 활성
(1,3)-β-D-glucan synthase 활성측정을위한반응은 Kurtz [16]의 방법을 약간 변형하였다: 최종 50µl의 부피로 reaction buffer [50 mM Tris.Cl (pH 7.4), 4 mM EDTA, 20µM GTP, 0.5% Brij 35, 6.6% glycerol, 2 mM UDP-glucose], 100µg의캔디다 membrane protein과 S. baicalensis를혼 합하여 25oC에서 30분간반응시켰다. 6 N NaOH를 10µl 넣 어반응을정지시킨후생성된 glucan을 80oC로 30분간용해 시켰다. 210µl의 aniline blue mix [40:21:59의 0.1% aniline blue, 1 N HCl, 1 M glycine/NaOH buffer (pH 9.5)]를넣은 후 50oC에서 30분간 반응시킨 다음 fluorescence reader (Bio-Tek, USA)를 사용하여 excitation 400 nm, emission 460 nm에서 50의감도로형광의양을측정하였다. reaction buffer, stop solution, aniline blue mix의 형광값을 background로하여 (1,3)-β-D-glucan synthase 활성을다음 의식에따라계산하였다.
(1,3)-β-D-glucan synthase activity (%)
= (fluorescence400/460 of S. baicalensis-treated sample- fluorescence400/460 of background)/(fluorescence400/460 of non-treated sample-fluorescence400/460 of background)
×100
FUN-1과 Calcofluor White를 사용한 이중 염색 대수기의 C. albicans ATCC 18804를 YNB/Glc에 최종 농도 2×106CFU/ml로희석하였다. 음성대조군을포함하 여 MIC의 S. baicalensis를처리한후 37oC, 200 rpm에서 3시간동안진탕배양하였다. 1 ml의배양액을 5,000 g로 1분 간원심분리하여회수하고 Phosphate buffered saline (PBS)으 로헹군후 50µl의 PBS로균질하게녹였다. 20µM의 FUN- 1 (Invitrogen, USA)과 25µM의 Calcofluor white (Sigma, USA)를넣어상온의암소에서 1시간이상염색한후 triple pass filter가장착된형광현미경(Olympus, Japan)으로관찰 하였다.
Neutral red 염색에 의한 분석
대수기의 C. albicans ATCC 18804 배양액 (2×106CFU/
ml)에 MIC의 S. baicalensis를처리한후 37oC, 200 rpm에서 진탕배양하였다. 4시간후 5,000 g로 1분간원심분리하여 회수한후 PBS로헹구었다. 같은양의 5 mg/ml neutral red 로혼합한후위상차현미경으로관찰하였다.
생존률의 분석
대수기의 C. albicans ATCC 18804를 YNB/Glc에최종농 도 2×107 CFU/ml로희석하였다. MIC의 S. baicalensis를 처리한후 37oC에서 200 rpm으로 4시간동안진탕배양하 였다. 1 ml의 배양액을 5,000 g로 1분간 원심분리하여 Candida를회수한후 phosphate-buffered saline (PBS)로헹 구었다. MuseTM Count and Viability kit (Merck Millipore, Germany)를제조사의방법에따라처리한후 MuseTM Cell
Analyzer를사용하여생존률을측정하였다. 음성대조군을약
95%의생존률으로조정한후 S. baicalensis 의효과를분석 하였다.
Time-kill curves
대수기의 C. albicans ATCC 18804 배양액을 YM 배지에 2×104CFU/ml로희석한후음성대조군을포함하여 MIC 의 S. baicalensis를넣은후 37oC, 200 rpm으로진탕배양 하였다. 정해진시간마다 100µl의배양액을새로운튜브에 옮긴후 20초이상교반하여출아직후의결합된효모를분 리하였다. 증류수로적당하게희석하고희석액 100µl를 YM
agar plate에도말하여 37oC에서 24시간배양한후집락의갯 수를기록하였다. 결과에나타난집락의수는 quadruplicate 로관찰된 3회의실험중하나를나타낸다.
S. baicalensis에 의한 세포주기 변화
대수기의 C. albicans ATCC 18804 배양액(1×108CFU /ml)에 S. baicalensis를처리한후 37oC에서 200 rpm으로 진탕 배양하였다. 1, 2, 3시간 후 각각 1 ml의 배양액을 5,000 g로 1분간원심분리하여세포를회수한후 PBS로헹 구었다. 1 ml의 4oC ethanol/PBS (7:3)를넣고 -20oC에서 15 분이상고정하여세포막을 permeabilization시켰다. 0.5 ml 의 PBS로헹군후 0.5 ml의 PBS를넣고최종농도 200µg/
ml RNase A (Sigma, USA)를넣었다. 50oC에서 1시간반응 시킨후, DNA 염색을위해 50µg의 propidium iodide 를넣 고 30분간어두운곳에놓아둔후 MuseTM Cell Analyzer로 분석하였다.
통계분석
모든실험은 2 또는 3회반복하였으며통계분석은 t-test 로, 두그룹간의차이는 Paired t-test로 SigmaPlot 10.0을 이용하여분석하였다. p값이 0.05 미만일때통계적으로유 의한것으로보았다.
결과 및 고찰
임상적으로중요한캔디다인 C. albicans, C. glabrata, C.
krusei, C. tropicalis에대하여 S. baicalensis 수용성추출
물의항진균활성을 resazurin을적용한미량희석법에의해
분석하였다(Table 1). 식물추출물은다양한화합물을포함 하므로단일성분보다높은시너지항진균효과를내는장 점을가지지만, 많은식물추출물이진한색을띠거나실험 을할때불용성침전물을형성하는경우가많아결과에대 한 해석을 방해하는 경우가 많다. 따라서 본 실험에서는 resazurin을포함시켜탁도에의존하는 CLSI A27-3의기존 방법을보완하였다. resazurin은무독성의염료로서살아있
는세포에의해 resorufin으로환원되어무형광의파란색에
서형광의분홍색으로변하므로[20], 색변화가나타나지않 는최저농도가 MIC이며보다객관적인결과를보여준다. S. baicalensis의 C. albicans ATCC 18804에대한 MIC는 1.56 mg/ml이었고, 분석에이용한다른 Candida sp. 보다높 은 MIC 값을나타냈다. S. baicalensis의 10가지 C. albicans 임상분리균주에대한 MIC 또한표준 C. albicans로사용된 C. albicans ATCC 18804와모두동일한 1.56 mg/ml을나 타냈다(Table 1).
임상 C. albicans 분리균주들이 3시간동안형성한각바
이오필름에대해 S. baicalensis의바이오필름제거효과를조 사하였다. 24시간동안 S. baicalensis에노출된바이오필름 은 MIC에서 57.7±17.3%, 2MIC에서 79.7±8.2%가제거되 었고, Paired-t test로분석한대조군과의차이는대부분 95%
또는 99% 수준에서유의성을나타냈다(Table 2).
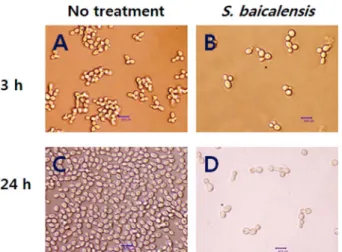
C. albicans의 폴리스티렌 표면 부착에 대한 S. baical-
ensis의영향을알아보기위해 3시간동안형성된바이오필
름에 S. baicalensis를처리한후의바이오필름변화를위상 차현미경을통해분석하였다. 3시간과 24시간후관찰된 S.
baicalensis의바이오필름제거효과는우수했으며전형적인
Table 1. Inhibitory activity of S. baicalensis extract against Candida spp.
Candida species
Concentration S. baicalensis
(mg/ml)
amphotericin B (µg/ml)
C. albicans ATCC 18804 1.56 0.25
C. glabrata ATCC 2001 0.05 0.13
C. krusei ATCC 750 0.20 0.25
C. tropicalis ATCC 32196 0.20 0.13
C. albicans (clinical, n=10) 1.56 NDa The MIC endpoint is based on visual determination using tur- bidity levels according to CLSI M27-A3 protocol and the colori- metric determination using an oxidation-reduction indicator resazurin [17]. NDa: not determined.
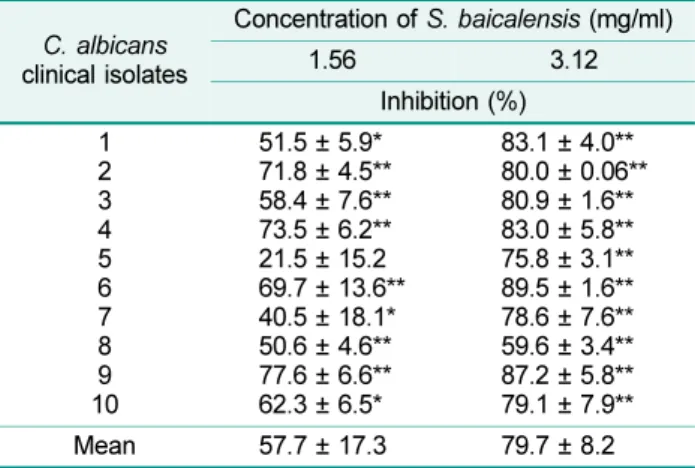
Table 2. The effect of the S. baicalensis extract on the metabolic activity of preformed C. albicans biofilms, expressed as percentage inhibition.
C. albicans clinical isolates
Concentration of S. baicalensis (mg/ml)
1.56 3.12
Inhibition (%) 1
2 3 4 5 6 7 8 9 10
51.5 ± 5.9*
71.8 ± 4.5**
58.4 ± 7.6**
73.5 ± 6.2**
21.5 ± 15.2 69.7 ± 13.6**
40.5 ± 18.1*
50.6 ± 4.6**
77.6 ± 6.6**
62.3 ± 6.5*
83.1 ± 4.0**
80.0 ± 0.06**
80.9 ± 1.6**
83.0 ± 5.8**
75.8 ± 3.1**
89.5 ± 1.6**
78.6 ± 7.6**
59.6 ± 3.4**
87.2 ± 5.8**
79.1 ± 7.9**
Mean 57.7 ± 17.3 79.7 ± 8.2
Metabolic activity was assessed using the XTT reduction assay measuring the absorbance at 492 nm. Inhibition percentages were calculated as 100× (1-A492 with S. baicalensis/ A492 without S. baicalensis). The differences in the optical density (mean ± standard deviation) of individual biofilms incubated with S.
baicalensis extract and control values were calculated using Paired Student's t-test, and were considered statistically signif- icant when p-value was 0.05. The experiments were performed three times with four replicates. *p≤ 0.05, **p ≤ 0.01.
난원형의대조군에비해실험군에서는둥글고팽창된모습 을나타내는세포가많이관찰되었다(Fig. 1).
S. baicalensis를처리한실험군의세포형이대부분둥글
고안정적으로부착되지않은형태에착안하여효모세포벽의 (1,3)-β-D-glucan polymer 형성에관여하는 (1,3)-β-D-glucan
synthase 활성에미치는 S. baicalensis의효과를분석하였다. Fig. 2에보여지는바와같이 1.56 mg/ml의 S. baicalensis의 존재하에서형성된 (1,3)-β-D-glucan polymer의양은대조군 에 비해 67.4 ± 3.1%로 감소되었으며 3.12 mg/ml의 S.
baicalensis 존재하에서는 42.4 ± 1.0%로 1,3-β-D-glucan polymer의 형성량이 급감하였다. 또한 (1,3)-β-D-glucan synthase의활성은 S. baicalensis의존재하에서농도-의존 적으로감소하였다.
S. baicalensis의 C. albicans 세포에대한형태적변화효 과를관찰하기위해 FUN-1과 Calcofluor white를동시에염 색시켜 triple pass filter가장착된형광현미경을통해관찰 하였다. Calcofluor white는살아있거나죽은세포의세포벽 을형광의파란색으로염색시킨다. FUN-1은세포막을통과
할수있는형광 probe로서대사적으로활발한효모는세포
질에붉은 CIVS(cylindrical intravacuolar structure)를나
타내지만[19] 죽은세포의세포질은옅은담황색의형광으로
염색[11]시키므로효모의대사와생존여부를관찰하기에적
합하다. Fig. 3A에보여지는바와같이대조군의현미경적인
이미지는정상적인출아형태를나타내고있으며붉은색의
세포내 CIVS를보여줌으로써이들세포가대사적으로활발
함을시사한다. S. baicalenis를처리한실험군(Fig. 3B)은대 부분둥글고부풀어확장된풍선형을나타냈으며, 붉은색의 CIVS를함유하는세포는소수로발견되었다. 실험군은또한
C. albicans의밀도가낮았고, 세포 주변의외액을흡수한
Fig. 1. Effect of S. baicalensis on C. albicans adhesion on polystyrene surfaces.
Microscopic view of C. albicans biofilms after treating without (A) and with 1.56 mg/ml of the S. baicalensis (B) on preformed bio- films for 3 h and 24 h, respectively. Bar = 10.0 µm.
Fig. 2. Cell wall (1,3)-β-D-glucan formation is decreased by the S. baicalensis extract.
Reaction mixture containing Tris.Cl (pH 7.4), GTP, Brij 35, UDP- glucose and yeast membrane protein were incubated in the absence and presence of S. baicalensis for 30 min at 25oC. Glu- can produced was solubilized and aniline blue mix was added.
Fluorescence with excitation wavelength at 400 nm and emis- sion wavelength at 460 nm was quantified with a fluorescence plate reader [16]. Error bars represent standard deviations (n = 4).
Fig. 3. FUN-1 and Calcofulor white double staining (A and B) and neutral red staining (C and D) of C. albicans.
(A) and (B): C. albicans stained with FUN-1 and Calcofluor white were viewed by fluorescence microscope. (A) Viable C. albicans with characteristic red CIVS inclusions; (B) S. baicalensis- treated swollen round cells with densely stained cell walls. Both live and dead yeast cell walls were stained equally well with Cal- cofluor white. (C) and (D): C. albicans stained with neutral red were viewed by phase contrast microscope. (C) Live yeast cells with red vacuole; (D) Dead yeast cells with red cytoplasm. An arrow indicates ruptured cell debris. Bar = 10.0 µm.
듯팽창된 형태를 나타내는 세포가 많았다. 이들은 또한
Calcofluor white에의해진하게세포벽이염색되는특징을
나타냈다. C. albicans의생존여부와세포형태를관찰하기
위한다른방법으로 C. albicans를 neutral red로염색한후 위상차현미경으로관찰하였다. Neutral red는생체염료로 서죽은세포는세포질전체가붉게염색되나살아있는세 포는세포질내의 vacuole만이붉게염색된다[14]. Fig. 3C 의대조군은정상적인난원형이며세포질내에붉은 vacuole 이관찰되었으나, Fig. 3D에보여지는바와같이실험군에 서는세포질전체가어둡게염색된세포가발견되거나세포
벽이파열되어흩어져있는 cell debris가여러곳에관찰되
었고, 사진은그중하나를나타낸다.
(1,3)-β-D-glucan synthase의활성이저해되면더얇고약 한세포벽이형성될것으로예상되었으나, Fig. 3B의결과처 럼 Calcofluor white로염색된실험군의세포벽은일관성이있 게더욱밝고두꺼우며진하게염색이되는부분이관찰되었 다. 본실험의결과와유사하게 lipopeptide에의해 (1,3)-β-D- glucan synthase의활성이저해된 Aspergillus fumigutus에 서도두꺼운세포벽과비정상적으로팽창된균사, 확장된공 같은형태등이관찰되었다[16]. 이는세포벽의한구성요소 의합성의저해로인해다른구성요소를더많이투입시켜
세포벽의결함을보상하려는메커니즘이작용한것으로보 여지나, 이를밝히기위해서는좀더심화된연구가필요할 것으로생각된다.
S. baicalensis의항진균효과에대한메커니즘을분석하기 위해 S. baicalensis를넣고 4시간동안배양한후 C. albicans 의생존률을조사하였다. 살아있는세포와죽은세포에대한
투과성이다른 두 가지의 DNA 결합 형광염료를 넣은 후
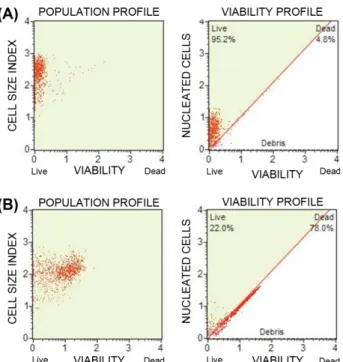
MuseTM Cell analyzer로분석하였고, 대조군의생존률을약 95%로보정한후실험군의생존률을측정하였더니 22%의 생존률을나타내었으며(Fig. 4), 이는 S. baicalensis가급속 하게세포의생존률에영향을준다는사실을시사한다.
S. baicalensis를넣고배양한 C. albicans의시간대별생 존한개체에대한 colony count 분석법을시행하였다. Fig.
5에보여진바와같이 9시간처리후 C. albicans의생존률 은 0.93 ± 0.3%이었고, 24시간 후의 생존률은 0.01% 이하 [26]로이는 S. baicalensis의 C. albicans에대한효과가살 진균성이라는것을의미한다.
C. albicans에 S. baicalensis를처리한후 1, 2, 3시간후 의세포주기를 MuseTM Cell Analyzer로분석하였다. 대조군 은시간이지나면서 G0/G1과 S기에있는세포들의비율이감 소하면서 G2기로진행되는것을관찰할수있었다(Table 3).
그러나 S. baicalensis를처리한실험군에서는 G0/G1의비율 이일정하게유지(G0/G1 arrest) 되어 S기로의세포주기진 행을 억제함을 알 수 있었다. 따라서 C. albicans는 S.
baicalensis로유발된새로운환경에대한적응의방책으로
세포분열의정지를선택한것으로보인다. 이는 C. albicans 의세포벽합성이저해되면서불필요하게 S기로진행하여 세포의많은에너지를소비하는일이없도록미리 S기로의 전진을차단하는경로를선택한것으로생각된다.
(1,3)-β-D-glucan synthase에대한저해제의효과가살진 균성[16]이라는보고처럼 S. baicalensis의효과도 (1,3)-β-D-
glucan synthase를억제하면서살진균활성을나타내었다
(Fig. 5). 본실험의결과에따르면 S. baicalensis는서론에 서언급했듯이캔디다세포벽무게의 47-60%를차지하는중 요한구성요소인 (1,3)-과 (1,6)-β-D-glucan 중하나인 (1,3)-β-
Fig. 4. Effect of the S. baicalensis on cell viability of C.
albicans.
Subcultured C. albicans cells (2× 107 CFU/ml) were treated without (A) and with S. baicalensis (B) and incubated for 4 h with shaking and 37oC. After washing with PBS, the cells were stained by mixing cells with MuseTM Count & Viability reagent and analyzed by MuseTM Cell Analyzer.
Table 3. Effect of the S. baicalensis extract on cell cycle of C. albicans.
No treatment S. baicalensis
1 h 2 h 3 h 1 h 2 h 3 h
G0/G1 49.7 38.1 20.0 49.9 50.5 54.8
S 24.8 27.5 18.8 21.9 22.7 19.6
G2 15.4 23.5 30.6 16.6 16.2 15.8
C. albicans ATCC 18804 were stained with MuseTM Cell Cycle Kit, and acquired on the MuseTM Cell Analyzer. Population per- centages were analyzed with the MuseTM cell cycle software module.
D glucan polymer의합성을억제시켜온전한세포벽형태유 지를방해함으로써세포가정상적인난원형을취하지못하 게하며쉽게세포외액을흡수하면서부풀어오른공모양을 형성하게하는것으로추정된다. 나아가세포는삼투압을견
디지못하면서세포막의파열(Fig. 3D)이일어나세포의죽
음으로연결되는것으로여겨진다. 즉, S. baicalensis 가캔 디다세포의바이오필름형성능력을억제하는것은세포벽 파열에의한세포의죽음, 비정상적인둥근형태로인한부
착능력감소, adhesin과같은부착단백질에대한지지대의
결여등이폴리스티렌에대한부착을억제하는요인으로작 용할것으로생각되며세포주기의 G0/G1 arrest 효과또한 출아의억제를통해바이오필름의증식억제에기여할것 으로보인다.
따라서본연구의결과는 S. baicalensis가바이오필름과 관련된캔디다증의치료를위한새로운항진균제개발에대 한후보물질로서의가능성을가짐을시사한다.
요 약
Candida 바이오필름은숙주조직과의료기기의표면에자
라는자가-조직화된미생물의군락으로전통적인항진균제 에대한저항성이높게나타난다. 황금(Scutellaria baical-
ensis)의뿌리는극동지방에서의료용목적으로널리사용되
어왔다. 본연구의목적은 10 C. albicans 임상분리균주에 의해형성된바이오필름에대한황금의수용성추출물의효 과를평가하고, 항바이오필름활성에대한메커니즘을알아
보는것이다. 바이오필름에대한효과는 XTT 환원분석법을 사용하였으며, 조사된모든균주에대한대사활성은 MIC에 서유의하게감소(57.7±17.3%)하였다. 황금추출물은 (1,3)- β-D-글루칸합성효소의활성을저해하였고 C. albicans의형 태에대한황금의효과는글루칸합성의억제로인한생장 의변화와관련이있었다: 대부분의세포는둥글고팽창되었 으며세포벽이진하게염색되거나파열되었다. 항캔디다활 성은살진균성이었고, 황금은 C. albicans를 G0/G1기에머물 게했다. 데이터는황금이목표가되는균류에다중의치명 적인효과를내며, (1,3)-β-D-글루칸합성효소의활성을저해 함을통해궁극적으로는세포벽의파열과죽음에이르게한 다는것을나타낸다. 따라서황금은바이오필름과관련된캔 디다의감염을치료하고제거하기위한항진균제개발후보 물질로서의가능성을가진다.
References
1. Beaulieu, D., J. Tang, S. B. Yan, J. M. Vessels, J. A. Radding, and T. R. Parr, Jr. 1994. Characterization and cilofungin inhibition of solubilized Aspergillus fumigatus (1,3)-β-D-glucan syn- thase. Antimicrob. Agents Chemother. 38: 937-944.
2. Blankenship, J. R. and A. P. Mitchell. 2006. How to build a bio- film: a fungal perspective. Curr. Opin. Microbiol. 9: 588-594.
3. Calderone, R. A. and P. C. Braun. 1991. Adherence and recep- tor relationships of Candida albicans. Microbiol. Rev. 55: 1-20.
4. Calderone, R. A. and W. A. Fonzi. 2001. Virulence factors of Candida albicans. Trends Microbiol. 9: 327-335.
5. Chandra, J., D. M. Kuhn, P. K. Mukherjee, L. L. Hoyer, T. McCor- Fig. 5. Time-kill curves for the S. baicalensis extract and relative viability of C. albicans.
C. albicans cells (2× 104 CFU/ml) were incubated with shaking in the absence and presence of S. baicalensis (1.56 mg/ml) at 37oC, and a colony count assay was used to determine the candidacidal activity of the S. baicalensis extract against C. albicans. Error bars represent standard deviations (n = 4).
mick, and M. A. Ghannoum. 2001. Biofilm formation by the fun- gal pathogen Candida albicans: development, architecture, and drug resistance. J. Bacteriol. 183: 5385-5394.
6. CLSI. 2008a. Reference method for broth dilution antifungal sus- ceptibility testing of yeasts; approved standard-third edition;
CLSI document M27-A3. Clinical and Laboratory Standards Institute, Wayne.
7. Douglas, C. M., F. Foor, J. A. Marrinan, N. Morin, J. B. Nielsen, A. M. Dahl, P. Mazur, W. Baginsky, W. Li, and M. el-Sherbeini.
1994. The Saccharomyces cerevisiae FKS1 (ETG1) gene en- codes an integral membrane protein which is a subunit of 1, 3- β-D-glucan synthase. Proc. Natl. Acad. Sci. U.S.A. 91: 12907- 12911.
8. Douglas, L. J. 2003. Candida biofilms and their role in infec- tion. Trends Microbiol. 11: 30-36.
9. Guarrera, P. M. 2005. Traditional phytotherapy in central Italy.
Fitotherapia 76: 1-25.
10. Haynes, K. 2001. Virulence in Candida species. Trends Micro- biol. 9: 591-596.
11. Henry-Stanley, M. J., R. M. Garni, and C. L. Wells. 2004.
Adaptation of FUN-1 and Calcofluor white stains to assess the ability of viable and nonviable yeast to adhere to and be inter- nalized by cultured mammalian cells. J. Microbiol. Methods 59: 289-292.
12. Huang, W. H., Lee, A. R., and C. H. Yang. 2006. Antioxidative and anti-inflammatory activities of polyhydroxyflavonoids of Scutellaria baicalensis GEORGI. Biosci. Biotechnol. Biochem.
70: 2371-2380.
13. Klis, F. M., A. F. J. Ram., and P. W. J. De Groot. 2007. A mole- cular and genomic view of the fungal cell wall. The Mycota 8:
97-120.
14. Kolotila, M. P., C. W. Smith, and A. L. Rogers. 1987. Candi- dacidal activity of macrophages from three mouse strains as demonstrated by a new method: neutral red staining. Med.
Mycol. 25: 283-290
15. Krcmery, V. and A. J. Barnes. 2002. Non-albicans Candida spp.
causing fungaemia: pathogenicity and antifungal resistance.
J. Hosp. Infect. 50: 243-260.
16. Kurtz, M. B., C. Douglas, J. Marrinan, K. Nollstadt, J. Onishi, S. Dreikorn, J. Milligan, S. Mandala, J. Thompson, and J. M.
Balkovec. 1994. Increased antifungal activity of L-733,560, a water-soluble, semisynthetic pneumocandin, is due to enhanc- ed inhibition of cell wall synthesis. Antimicrob. Agents Chemo- ther. 38: 2750-2757.
17. Liu, M., V. Seidel, D. R. Katerere, and A. I. Gray. 2007. Colori- metric broth microdilution for the antifungal screening of plant extracts against yeast. Methods 42: 325-329.
18. Mahmoud, A. G. and B. R. Louis. 1999. Antifungal agents:
mode of action, mechanism of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol.
Rev. 12: 501-517.
19. Millard, P. J., B. L. Roth, H. -P. Thi, S. T. Yue, and R. P.
Haugland. 1997. Development of the FUN-1 family of fluores- cent probes for vacuole labeling and viability testing of yeast.
Appl. Environ. Microbiol. 63: 2897-2905.
20. O'Brien, J., J. Wilson, T. Orton, and F. Pognan. 2000. Investi- gation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem.
267: 5421-5426.
21. Oliver, B. G., P. M. Silver, C. Marie, S. J. Hoot, S. E. Leyde, and T. C. White. 2008. Tetracycline alters drug susceptibility in Candida albicans and other pathogenic fungi. Microbiol. 154:
960-970.
22. Park, S. J., S. J. Choi, W. S. Shin, H. M. Lee, K. S. Lee, and K. H. Lee. 2009. Relationship between biofilm formation abil- ity and virulence of Candida albicans. J. Bacteriol. Virol. 39:
119-124.
23. Pfaller, M. A. and D. J. Diekema. 2007. Epidemiology of in- vasive candidiasis: a persistent public health problem. Clin.
Microbiol. Rev. 20: 133-163.
24. Ramage, G., J. P. Martinez, and J. L. Lopez-Ribot. 2006. Can- dida biofilms on implanted biomaterials: a clinically significant problem. FEMS Yeast Res. 6: 979-986.
25. Ramage, G., K. Vande-Walle, B. L. Wickes, and J. L. Lopez-Ribot.
2001. Standardized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob. Agents Chemother. 45: 2475-2479.
26. Roling, E. E., M. E. Klepser, A. Wasson, R. E. Lewes, E. J.
Ernst, and M. A. Pfaller. 2002. Antifungal activities of flucona- zole, caspofungin (MK0991), and anidulafungin (LY303366) alone and in combination against Candida spp. and Cryoto- coccus neoformans via time-kill methods. Diagn. Microbial.
Infec. Dis. 43: 13-17.
27. Shedletzky, E., C. Unger, and D. P. Delmer. 1997. A microtiter- based fluorescence assay for (1,3)-β-glucan synthases. Anal.
Biochem. 249: 88-93.
28. Tang, J. and T. R. Parr, Jr. 1991. W-1 solubilization and kinet- ics of inhibition by cilofungin of Candida albicans (1,3)-β-D-glucan synthase. Antimicrob. Agents Chemother. 35: 99-103.
29. Yoon, S. B., Y. J. Lee, S. K. Park, H. C. Kim, H. Bae, H. M. Kim, S. G. Ko, H. Y. Choi, M. S. Oh, and W. Park. 2009. Anti-inflam- matory effects of Scutellaria baicalensis water extract on LPS- activated RAW 264.7 macrophages. J. Ethnopharmacol. 125:
286-290.