Corresponding author: Seung-Soo Baek, Department of Physical Educa- tion, College of Arts and Physical Education, Sang Myung University, #7 Hongji-dong, Jongno-gu, Seoul 110-743, Korea
Tel: +82-2-2287-5133, E-mail: [email protected] Received November 24, 2010, Revised December 5, 2010 Accepted December 10, 2010
This work was supported by the Korea Research Foundation Grant funded by the Korea Government (MOEHRD) (KRF-2007-332-G00090).
Short-Term Repeated Treadmill Exercise More Potently Increases Cell Proliferation and Brain-Derived Neurotrophic Factor Expression in the
Hippocampus of Rats
Departments of *Biochemistry and Molecular Biology,
†Physiology, College of Medicine, Kyung Hee University, Seoul,
‡School of Sport Science, Sungkyunkwan University, Suwon,
§
Department of Ophthalmology, Chungju Hospital, Konkuk University College of Medicine, Chungju,
∥
Department of Physical Education, College of Arts and Physical Education, Sang Myung University, Seoul, Korea
Ji-Heon Jung*, Hyung-Hwan Baik*, Tae-Woon Kim
†,‡, Il-Gyu Ko
†, Eun-Sang Ji
†, Mal-Soon Shin
†, Bo-Kyun Kim
†, Jin-Hee Seo
†, Dong-Hee Kim
§, Seung-Soo Baek
∥, Young-Ho Jin
†Physical exercise enhances cell proliferation and/or neurogenesis in the hippocampal dentate gyrus. Brain-derived neurotrophic factor (BDNF) is implicated in the modulation of neurotransmitters turnover, neurite genesis, and neuronal survival. BDNF binds to the tyrosine kinas B (TrK). In this study, we compared the effects of the one bout of long-duration treadmill exercise and the repeated short-duration treadmill exercise on cell proliferation and expressions of BDNF and TrKB in the hippocampus. Adult male Sprague-Dawley rats, weighing 200±5 g, were randomly divided into three groups (n=8 in each group): control group, one hour-one bout-treadmill exercise group, and ten minutes-six bouts-treadmill exercise group. The rats in the one hour-one bout-treadmill exercise group were forced to run on a motorized treadmill for 60 min once a day (at 16:00). The rats in the ten minutes-six bouts-treadmill exercise group were forced to run on a treadmill once 10 minutes per six times for a day (at 10:00, 11:00, 14:00, 15:00, 19:00, 20:00). Treadmill exercise was continued for 7 consecutive days. The rats in the control group were left without running. Both types of treadmill exercise increased cell proliferation and enhanced expressions of BDNF and TrkB in the hippocampus. Cell proliferation and BDNF expression was more potently increased by the ten minutes-six bouts treadmill exercise than the one hour-one bout treadmill exercise. These results showed that repeated bouts of short-duration treadmill exercise may achieve more useful results for the brain functions than one-bout long-duration treadmill exercise. (Korean J Str Res 2010;18:295∼303)
Key Words: Treadmill exercise, Brain-Derived neurotrophic factor, Cell proliferation, Hippocampus, Rats
INTRODUCTION
Physical exercise is known to benefits not only to the
peripheral adaptive processes, such as cardiovascular and muscu-
loskeletal adaptation, but also to the brain functions (Meeusen,
2005). The hippocampus is the main brain area for cognitive
function, and the physical exercise enhances brains functions, especially in elderly who are vulnerable to the age-induced or brain diseases-induced decline of cognition (Churchill et al., 2002;
Colcombe et al., 2003; Kim SE et al., 2010). Previous studies re- ported that the hippocampus plays an important role in learning ability and memory capability (Sze et al., 1997; Eichenbaum, 2004). It has recently been firmly established that new cells are continuously produced in the hippocampal dentate gyrus of adult mammals, including humans (Eriksson et al., 1998; Gould et al., 1999; van Praag et al., 1999; Kim SH et al., 2002). Hippocampal neurogenesis after birth is closely correlated with learning ability and memory capability (Gould et al., 1999; van Praag et al., 1999; Shors et al., 2001; Kim H et al., 2007), while stresses tend to inhibit it (Heine et al., 2004). Physical exercise, specifically, has been shown to enhance cell proliferation and/or neurogenesis in the dentate gyrus (van Praag et al., 1999; Kim YP et al., 2003).
Newly generated cells in the dentate gyrus of hippocampus are implicated in the cognitive function and repair of brain (van Praag et al., 2002; Dong et al., 2004; Kim H et al., 2007). Dong et al.(2004) demonstrated that cell proliferation in the dentate gyrus was decreased in the animal model of Alzheimer’s disease.
Brain-derived neurotrophic factor (BDNF) is a member of the nerve growth factor family of neurotrophins, and it has been detected in many areas of the brain (Friedman et al., 1998).
BDNF is also been implicated in the modulation of neuro- transmitters turnover, neurite genesis, and neuronal survival (No- nomura et al., 1992; Ghosh et al., 1994). BDNF is known to upregulate the induction of long-term potentiation (LTP) (Figurov et al., 1996), enhance synaptic transmission (Schinder et al., 2000), and also increase neuronal plasticity in the central nervous system (CNS) (Mizuno et al., 2000). Many studies indicated that exercise increases the expression of BDNF mRNA in the rat’s hippocampus (Russo-Neustadt et al., 2000; Johnson et al., 2003;
Kim H et al., 2007). In this respect, BDNF has been tentatively implicated in brain plasticity associated with physical exercise (Cotman et al., 2002). BDNF binds to the tyrosine kinas (TrK) B (Chao, 2003). Inhibition of BDNF or down regulation TrkB caused memory impairment, decreased synaptic innervations, and reduced levels of synaptic vesicle proteins (Cotman et al., 2002;
Liu et al., 2008).
It was suggested that the influence of exercise on cell proli-
feration and neurogenesis is modulated by both circadian phase and the amount of daily exercise (Holmes et al., 2004). There is no indication on the threshold duration of exercise to achieve training effects. In the guidelines of American College of Sports Medicine (ACSM), at least 20 min duration of aerobic exercise was recommended. These guidelines are based on the VO2max and heart ratemax. The relationship between exercise and brain function have well been recognized, however, it is hard to take enough time performing exercise in the busy lifestyle. For this reason, evaluation of minimal time to achieve the beneficial effects of exercise on brain functions has been required. In the present study, we compared the effects of one bout long-duration of treadmill exercise and repeated short-duration of treadmill exercise on brain functions. For this, cell proliferation in the hippocampal dentate gyrus was evaluated by immunohistochemistry for 5-bromo-2'-deoxyuridine (BrdU) and Ki67. The expressions of BDNF and TrkB in the hippocampus were determined by Western blot analysis.
MATERIALS AND METHODS
1. Animals and treatments
Adult male Sprague-Dawley rats, weighing 200±5 g (7 weeks old), were obtained from commercial breeder (Orient Co., Seoul, Korea) for the experiment. The experimental procedures were performed in accordance with the animal care guidelines of the National Institutes of Health (NIH) and the Korean Academy of Medical Sciences. The animals were housed under controlled temperature (23±2
oC) and lighting (08:00 to 20:00 h) conditions with food and water available ad libitum. The rats were randomly divided into three groups as follows (n=8 in each group): the control group, the one hour-one bout-treadmill exercise group, and the ten minutes-six bouts-treadmill exercise group. All rats received 50 mg/kg BrdU (Sigma Chemical Co., St. Louis, MO, USA) at 09:00 intraperitoneally once a day in first three experimental days.
2. Exercise protocols
The rats in one hour-one bout-treadmill exercise group was
forced to run on a motorized treadmill for 60 min once a day
(at 16:00), and the rats in the ten minutes-six bouts-treadmill
exercise group forced to run on a treadmill once 10 minutes per six times for a day (at 10:00, 11:00, 14:00, 15:00, 19:00, 20:00).
This treadmill exercise was continued for 7 consecutive days. The exercise load in both groups consisted of running at a speed of 8 m/min with a 0° inclination. The rats in the control group were left without running.
3. Tissue preparation
The rats were sacrificed immediately after last exercise. The rats were anesthetized using Zoletil 50
Ⓡ(10 mg/kg, i.p.; Vibac Laboratories, Carros, France), transcardially perfused with 50 mM phosphate-buffered saline (PBS), and fixed with a freshly prepared solution consisting of 4% paraformaldehyde in 100 mM phos- phate buffer (PB, pH 7.4). The brains were dissected and post-fixed in the same fixative method overnight, and transferred to a 30% sucrose solution for cryoprotection. Forty μm thick coronal sections were made using a freezing microtome (Leica, Nussloch, Germany). On average 10 slice sections in the hippo- campal region were collected from each rat. The sections obtained 2.5∼2.7 mm posterior from the bregma were used for immuno- histochemistry.
4. BrdU immunohistochemistry
To detect newly generated cells in the dentate gyrus, BrdU- specific immunohistochemistry was performed. The sections were first permeabilized by incubation in 0.5% Triton X-100 in PBS for 20 min, then pretreated in 50% formamide-2×standard saline citrate (SSC) at 65
oC for 2 h, denatured in 2 N HCl at 37
oC for 30 min, and rinsed twice in 100 mM sodium borate (pH 8.5). Afterwards, the sections were incubated overnight at 4
oC with BrdU-specific mouse monoclonal antibody (1 : 600; Roche, Mannheim, Germany). The sections were then washed three times with PBS and incubated with biotinylated mouse secondary antibody (1 : 200; Vector Laboratories, Burlingame, CA, USA) for 1 h. The sections were then incubated for another 1 h with an avidin- peroxidase complex (1 : 100; Vector Laboratories). For visualization, sections were incubated in 50 mM Tris-HCl (pH 7.6) containing 0.03% diaminobenzidine (DAB), 40 mg/ml nickel chloride, and 0.03% hydrogen peroxide for 5 min. The sections were mounted onto gelatin-coated slides, air-dried overnight at room temperature, and coverslips were mounted using Permount
Ⓡ.
5. Ki67 immunohistochemistry
To detect newly generated cells in the dentate gyrus, Ki67- specific immunohistochemistry was performed. The sections were first permeabilized by incubation in 0.5% Triton X-100 in PBS for 20 min, then pretreated in 50% formamide-2×standard saline citrate (SSC) at 65
oC for 2 h, denatured in 2 N HCl at 37
oC for 30 min, and rinsed twice in 100 mM sodium borate (pH 8.5).
Afterwards, the sections were incubated overnight at 4
oC with mouse anti-Ki67 antibody (1 : 200; Novocastra Laboratories, Newcastle, UK). The sections were then washed three times with PBS and incubated with biotinylated mouse secondary antibody (1 : 200; Vector Laboratories) for 1 h. The sections were then incubated for another 1 h with an avidin-peroxidase complex (1 : 100; Vector Laboratories). For visualization, sections were incubated in 50 mM Tris-HCl (pH 7.6) containing 0.03%
DAB, 40 mg/ml nickel chloride, and 0.03% hydrogen peroxide for 5 min. The sections were mounted onto gelatin-coated slides, air- dried overnight at room temperature, and coverslips were mounted using Permount
Ⓡ.
6. BDNF and TrkB Western blot analysis
The hippocampal tissues were collected, and then were immediately frozen at −70
oC. The hippocampal tissues were homogenized on ice, and lysed in a lysis buffer containing 50 mM HEPES (pH 7.5), 150 mM NaCl, 10% glycerol, 1% Triton X-100, 1 mM PMSF, 1 mM EGTA, 1.5 mM MgCl
2·6H
2O, 1 mM sodium orthovanadate, and 100 mM sodium flouride. Protein content was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein (30 μg) was sepa- rated on SDS-polyacrylamide gels and transferred onto a nitro- cellulose membrane. Mouse actin antibody (1 : 500; Santa Cruz Biotechnology), rabbit BDNF antibody (1 : 1,000; Santa Cruz Biotechnology) and rabbit TrkB (1 : 1,000; Santa Cruz Biotecho- logy) were used as the primary antibodies. Horseradish peroxi- dase-conjugated anti-rabbit antibody for BDNF (1 : 2,000; Vector Laboratories) and TrkB (1 : 3,000; Vector Laboratories) were used as the secondary antibodies.
Experiments were performed in normal laboratory conditions and at room temperature, except for the transferred membranes.
Transferred membranes were performed at 4
oC with the cold pack
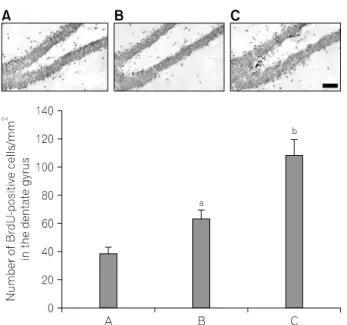
Fig. 1. Effects of treadmill exercise on the BrdU-positive cells in the dentate gyrus. Upper: Photomicrographs of 5-bromo-2’-deoxyuridine (BrdU)-positive cells in the dentate gyrus. Scale bar represents 400 μm.
Lower: Number of BrdU-positive cells in each group. (A) Control group, (B) one hour-one bout-treadmill exercise group, (C) ten minutes-six bouts-treadmill exercise group. The data are represented as the mean±
S.E.M.
ap<0.05 compared to the control group.
bp<0.05 compared to the one hour-one bout-treadmill exercise group.
and pre-chilled buffer. Band detection was performed using the enhanced chemiluminescence (ECL) detection kit (Santa Cruz Biotechnology).
7. Data analysis
The numbers of BrdU-positive and Ki67-positive cells in the dentate gyrus were counted hemilaterally under a light micro- scope (Olympus, Tokyo, Japan), and they were expressed as the numbers of cells per square millimeter in the dentate gyrus. The area of the dentate gyrus was measured by Image-Pro
ⓇPlus image analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA.). For confirming the expressions of BDNF and TrkB, the detected bands were calculated densitometrically using Molecular Analyst
TM, version 1.4.1 (Bio-Rad).
The data were analyzed with one-way ANOVA and then Duncan post-hoc tests. All values are expressed as the mean±
standard error of the mean. The p value<0.05 was considered significant.
RESULTS
1. Effect of treadmill exercise on BrdU-positive cells in the dentate gyrus
Photomicrographs of BrdU-positive cells in the hippocampal dentate gyrus are presented in Fig. 1. The number of BrdU- positive cells was 38.62±4.94/mm
2in the control group, 63.99±5.15/mm
2in the one hour-one bout-treadmill exercise group, 109.31±10.49/mm
2in the ten minutes-six bouts-treadmill exercise group.
These result showed that both types of treadmill exercise increased the number of BrdU-positive cells in the hippocampal dentate gyrus (p<0.05). The number of BrdU-positive cells in the hippocampal dentate gyrus was more potently enhanced in the ten minutes-six bouts-treadmill exercise group compared to that in the one hour-one bout-treadmill exercise group (p<0.05).
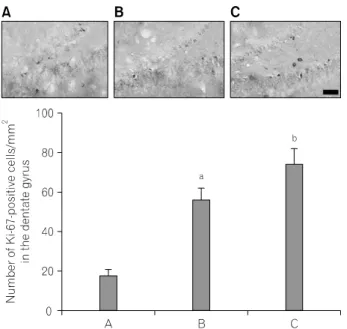
2. Effect of treadmill exercise on Ki67-positive cells in the dentate gyrus
Photomicrographs of Ki67-positive cells in the hippocampal dentate gyrus are presented in Fig. 2. The number of Ki-67- positive cells was 18.47±2.67/mm
2in the control group, 56.81±5.86/mm
2in the one hour-one bout-treadmill exercise group, 75.13±7.51/mm
2in the ten minutes-six bouts-treadmill exercise group.
These result showed that both types of treadmill exercise increased the number of Ki67-positive cells in the hippocampal dentate gyrus (p<0.05). The number of Ki67-positive cells in the hippocampal dentate gyrus was more enhanced in the ten minutes-six bouts-treadmill exercise group compared to that in the one hour-one bout-treadmill exercise group (p<0.05).
3. Effect of treadmill exercise on the expressions of BDNF and TrkB proteins
The BDNF (14 kDa) and TrkB (95-145 kDa) protein expres- sions are presented Fig. 3. When the level of BDNF in the control group was set at 1.00, the level of BDNF was 1.11±0.05 in the one hour-one bout-treadmill exercise group, 1.59±0.08 in the ten minutes-six bouts-treadmill exercise group.
These result showed that both types of treadmill exercise
Fig. 3. Effects of treadmill exercise on expression of BDNF and TrkB proteins in the hippocampus. (A) Control group, (B) one hour-one bout-treadmill exercise group, (C) ten minutes-six bouts-treadmill exercise group. The data are represented as the mean±S.E.M.
ap<0.05 compared to the control group.
b