Corresponding author: Chang-Ju Kim, Department of Physiology, College of Medicine, Kyung Hee University, 1, Hoeigi-dong, Dongdaemoon-gu, Seoul 130-701, Korea
Tel: +82-2-961-0407, E-mail: [email protected] Received August 15, 2011, Revised September 1, 2011 Accepted September 20, 2011
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2009-352-G00056).
Various Types of Stress Deteriorate Spatial Memory through Inhibition of Cell Proliferation in the Hippocampus of Rats
Departments of *Biochemistry and Molecular Biology,
†Physiology, College of Medicine, Kyung Hee University,
‡
Department of Physical Education, Korea Military Academy, Seoul, Korea
Jee-Sang Yoon*, Hyung-Hwan Baik*, Su-Shin Lee
†, Yu-Mi Heo
†, Bo-Kyun Kim
†, Mal-Soon Shin
†, Sang-Won Lee
‡, Chang-Ju Kim
†Stress is defined as physical, physiological, and psychological modifications that disrupt the homeostasis and internal balance of the body. Stress is closely associated with neuronal changes such as impaired neurogenesis, neuronal atrophy, and neuronal apoptosis in the hippocampus. In the present study, we investigated the effects of various types of stress (restraint stress, noise stress, cold stress) on spatial memory, cell proliferation, and expression of brain-derived neurotrophic factor (BDNF) and its receptor tyrosine kinase B (TrkB) in the hippocampus of rats. The rats in the restraint stress group were restrained in a transparent cylinder.
The rats in the noise stress group were exposed to a 120 dB supersonic machine sound. The rats in the cold stress group were exposed to a cold chamber at 4
oC. The rats in the control group were left undisturbed. Application of each stress was conducted for 60 min 2 times a day, and continued for 10 days. In the present results, all three types of stress induced spatial memory impairment. All three types of stress suppressed cell proliferation and inhibited expression of BDNF and TrkB in the hippocampus.
The strength of the deteriorating and suppressing effects of stresses had the following rank order: restraint stress>noise stress>
cold stress. Thus, impairment of spatial memory by stresses may be due to the inhibition of cell proliferation through suppressing BDNF expression in the hippocampus. (Korean J Str Res 2011;19:243∼251)
Key Words: Restraint stress, Noise stress, Cold stress, Spatial memory, Hippocampus
INTRODUCTION
Stress is defined as physical, physiological, and psychological modifications that disrupt the homeostasis and internal balance of
the body. Stress is known to be implicated in several neurodegenerative and psychiatric diseases including depression (Sung YH et al., 2010), stroke (Manolescu et al., 2011), and Alzheimer's disease (Clark et al., 2010). Stress causes structural changes and neuronal damage, especially in the hippocampus (Smith et al., 1995), which is a region that is important in learning ability and memory function. Stress is closely associated with neuronal changes such as impaired neurogenesis, neuronal atrophy, and neuronal apoptosis in the hippocampus (McEwen et al., 2001; Sung YH et al., 2010).
Neurogenesis is associated with hippocampal function,
including learning ability and memory function, and neurogenesis is modulated by a variety of factors including stress, an enriched environment, learning, serotonin, N-methyl-D-aspartate (NMDA) receptor antagonists, and stroke (Kempermann et al., 1997; Fuchs et al., 2000; Ko IG et al., 2009). Stress inhibits cell proliferation of new granular cells in the hippocampal dentate gyrus (Fuchs et al., 2001; Tanapat et al., 2001).
The neurotrophic factors include nerve growth factor (NGF), neurotrophin-3 (NT-3), neurotrophin-4/5 (NT-4/5), and brain-de- rived neurotrophic factor (BDNF). Among these, BDNF acts as a modulator of long-term potentiation (LTP), synaptic plasticity, and neuronal plasticity in the adult central nervous system (Martin et al., 2000; Lee E et al., 2009). BDNF is found in many areas of the brain (Smith et al., 1996; Givalois et al., 2001;
Alonso et al., 2002). BDNF and its receptor, tyrosine kinase B (TrkB), are crucial for hippocampal-dependent learning ability and memory function (Patterson et al., 1996). In addition, BDNF promotes survival and growth of neurons in the hippocampus (Lowenstein et al., 1996; Bartrup et al., 1997). BDNF is known as a key molecule in stress, because BDNF expression is decreased by exposure to stress (Smith et al., 1995; Adlard et al., 2004).
Stress is known to reduce neurogenesis through the suppression of BDNF expression in the hippocampus (Lucassen et al., 2010).
However, the effects of various types of stress (restraint stress, noise stress, cold stress) on spatial memory in relation to cell proliferation have not been compared, and the exact underlying mechanisms of stress on memory function are unclear. In the present study, we investigated the effects of these kinds of stresses on spatial memory, cell proliferation, and expression of BDNF and TrkB in the hippocampus of rats. To this end, we used an 8-arm maze test for spatial memory, immunohistochemistry for 5-bromo-2'-deoxyuridine (BrdU) and Ki-67, and western blot analysis for BDNF and TrkB.
MATERIALS AND METHODS
1. Animals and treatments
Experimental procedures were performed in accordance with the guidelines of the National Institute of Health (NIH) and the Korean Academy of Medical Sciences. Male Sprague-Dawley rats weighing 200±10 g were used for the experiment. Food and
water were made available ad libitum. The rats were randomly divided into four groups (n=8 in each group): a control group, a restraint stress group, a noise stress group, and a cold stress group. All rats received 50 mg/kg BrdU (Sigma Chemical Co., St.
Louis, MO, USA) intraperitoneally 30 min before the start of first stress exposure, once a day for 5 consecutive days. The animals were sacrificed immediately after the completion of the behavior test.
2. Stress exposure
We applied each stress twice per day according to the previous studies (Endo et al., 1999; Adlard et al., 2004; Heine et al., 2005). The rats in the restraint stress group were restrained in a transparent Plexiglas cylinder (21 cm long, 6 cm diameter). The rats in the noise stress group were exposed to a 120 dB supersonic machine sound. The rats in the cold stress group were exposed to a cold chamber that was maintained at 4°C. The rats in the control group were left undisturbed. Application of each stress was conducted for 60 min, 2 times a day, and continued for 10 days.
3. Radial-8-arm maze test
Spatial memory was tested using a radial-8-arm maze apparatus as previously described method (Kim SE et al., 2010;
Ko IG et al., 2011). The radial-8-arm maze apparatus consisted of a central octagonal plate (30 cm in diameter) and eight radiating arms (each 50 cm in length and 10 cm in width). The apparatus was placed 1 m above the floor. A small receptacle filled with water (3 cm in diameter and 1 cm in depth) was located at the end of the arms. The rats were trained three times before the spatial learning test. The rats deprived of water for 24 h were allowed to explore for water and to drink for 5 min. On 9 days after each stress exposure, radial-8-arm maze test was performed. The time spent seeking water at the end of the arms was recorded. The test was terminated when a rat found water in all eight arms or over 5 min elapsed. Re-entering a previously visited arm was counted as an error. In addition, the number of correct choice before the first error was counted.
4. Tissue preparation
For brain tissue preparation, animals were fully anesthetized
with Zoletil 50
Ⓡ(10 mg/kg, i.p.; Vibac Laboratories, Carros, France), transcardially perfused with 50 mM phosphate-buffered saline (PBS), and fixed with freshly prepared solution consisting of 4% paraformaldehyde (PFA) in 100 mM phosphate buffer (PB, pH 7.4). Brains were then removed, post-fixed in the same fixative overnight, and transferred into a 30% sucrose solution for cryoprotection. Coronal sections of 40 μm thickness were made using a freezing microtome (Leica, Nussloch, Germany).
5. BrdU immunohistochemistry
For the detection of newly generated cells in the dentate gyrus, BrdU incorporation, which has been generally used as an indicator of DNA synthesis, was visualized using an immunohi- stochemical method as previously described method (Kim SE et al., 2010; Jung SY et al., 2011). Briefly, the sections were permeabilized by incubation with 0.5% Triton X-100 in PBS for 20 min, treated with 50% formamide-2× standard saline citrate (SSC) at 65
oC for 2 h, denatured in 2 N HCl at 37
oC for 30 min, and rinsed twice in 100 mM sodium borate (pH 8.5).
Subsequently, the sections were incubated overnight at 4
oC with BrdU-specific mouse monoclonal antibody (1 : 600; Roche, Mannheim, Germany). The sections were then washed three times with PBS and incubated for 1 h with a biotinylated mouse secondary antibody (1 : 200; Vector Laboratories, Burlingame, CA, USA). Then, the sections were incubated for another 1 h with avidin–peroxidase complex (1 : 100; Vector Laboratories). For the visualization of BrdU, the sections were incubated with 50 mM Tris-HCl buffer (pH 7.6) containing 0.02% 3,3'-diamino- benzidine (DAB), nickel chloride (40 mg/ml) and 0.03% H
2O
2for 5 min. After BrdU-specific staining, counter-staining was performed on the same sections using a mouse anti-neuronal nuclei (NeuN) antibody (1 : 300; Chemicon International, Teme- cula, CA, USA). The sections were washed three times with PBS, incubated for 1 h with a biotinylated anti-mouse secondary antibody, and processed with VECTASTAIN
ⓇABC kits. For staining, the sections were reacted with 0.02% DAB and 0.03%
H
2O
2in 50 mM Tris-HCl buffer (pH 7.6) for 5 min after which the sections were mounted onto gelatin-coated slides. The slides were air dried overnight at room temperature, and coverslips were mounted using Permount
Ⓡ.
6. Ki-67 immunohistochemistry
For the detection of Ki-67-positive cells in the dentate gyrus, immunohistochemistry was performed as previously described method (Sung YH et al., 2010). The sections were then incubated in 50 mM PBS for 10 min, washed 3 times using in 50 mM PBS, and then incubated in 3% H
2O
2for 30 min. After that, the sections were incubated overnight with mouse anti-Ki-67 antibody (1 : 500; Novocastra Laboratories, Newcastle, UK). Next day, we incubated the sections for 1 h with biotinylated anti-mouse secondary antibody (Vector Laboratories), and subsequently incubated them with an avidin-biotin-peroxidase complex (Vector Laboratories) for 1 h at room temperature.
Immunoreactivity was visualized by incubating the sections in a solution consisting of 0.05% DAB and 0.01% H
2O
2in 50 mM Tris-HCl buffer (pH 7.6) for approximately 3 min. The sections were then mounted on gelatin-coated glass slides, and coverslips were mounted using Permount
Ⓡ.
7. Western blot analysis
Western blot analysis was conducted for the determination of BDNF and TrKB expression as previously described method (Kim SE et al., 2010). Hippocampal tissues were collected and then immediately frozen at −70
oC. Hippocampal tissues were homogenized on ice, and lysed in a lysis buffer containing 50 mM HEPES (pH 7.5), 150 mM NaCl, 10% glycerol, 1% Triton X-100, 1 mM PMSF, 1 mM EGTA, 1.5 mM MgCl
2ㆍ6H
2O, 1 mM sodium orthovanadate, and 100 mM sodium fluoride. Protein content was measured using a Bio-Rad colorimetric protein assay kit (Hercules, CA, USA). Protein (30 μg) was separated on SDS-polyacrylamide gels and transferred onto a nitrocellulose membrane. Mouse beta actin antibody (1 : 500; Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit BDNF and TrkB antibody (1 : 1,000; Santa Cruz Biotechnology) were used as the primary antibodies. Horseradish peroxidase-conjugated anti-rabbit antibody for BDNF and TrkB (1 : 3,000; Vector Laboratories) were used as the secondary antibodies.
Experiments were performed under normal laboratory
conditions and at room temperature, except for the transfer to
membranes. The transfer was performed at 4°C with a cold pack
and pre-chilled buffer. Band detection was performed using
enhanced chemiluminescence (ECL) detection kits (Santa Cruz Biotechnology).
8. Data analysis
The area of the hippocampal dentate gyrus was measured using Image-Pro Plus software (Media Cybernetics, Silver Spring, MD, USA). The numbers of BrdU-positive and Ki-67-positive cells in the hippocampus were counted hemilaterally. The data are expressed as the number of cells per mm
2of the hippocampus.
To compare the relative expressions of BDNF and TrKB, the detected bands were calculated densitometrically using Image-Pro Plus software (Media Cybernetics).
Statistical analysis was performed using one-way ANOVA followed by Duncan's post-hoc test. The results are presented as the mean±standard error of the mean (S.E.M.). Significance was set as p<0.05.
RESULTS
1. Effects of various types of stress on spatial memory
To verify the effects of various types of stress on spatial memory, the radial-8-arm maze test was performed. All rats completed eight successful performances within 106.00±12.35 sec in the normal group, 253.28±14.13 sec in the restraint stress group, 226.62±22.91 sec in the noise stress group, and 154.42±14.04 sec in the cold stress group (Fig. 1A).
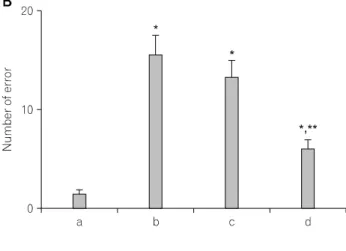
The number of errors made before eight successful perfor- mances was 1.28±0.47 in the normal group, 15.57±1.95 in the restraint stress group, 13.25±1.72 in the noise stress group, and 6.00±0.87 in the cold stress group (Fig. 1B).
The number of correct choices before the first error in the control group was 7.42±0.20 in the normal group, 3.28±0.42 in the restraint stress group, 3.62±0.37 in the noise stress group, and 5.57±0.42 in the cold stress group (Fig. 1C).
Fig. 1. The effects of various types of stress on spatial memory. (A)
The time for eight successful performances. (B) The number of errors
made before eight successful performances. (C) The number of correct
choices made before the first error. (a) Normal group, (b) restraint
stress group, (c) noise stress group, (d) cold stress group. The data are
mean±standard error of the mean (S.E.M.). *p<0.05 compared to the
normal group. **p<0.05 compared to the restraint stress group.
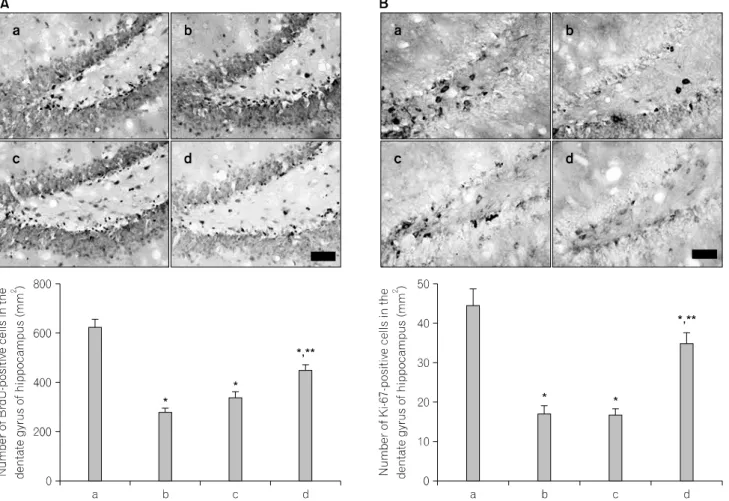
Fig. 2. The effects of various types of stress on cell proliferation in the hippocampus. (A) 5-Bromo-2'-deoxyuridine (BrdU)-positive cells. Upper:
Photomicrographs of BrdU-positive cells in hippocampal dentate gyrus. The scale bar is 50 μm. Lower: Number of BrdU-positive cells in the hippocampal dentate gyrus. (B) Ki-67-positive cells. Upper: Photomicrographs of Ki-67-positive cells in the hippocampal dentate gyrus. The scale bar is 50 μm. Lower: Number of Ki-67-positive cells in the hippocampal dentate gyrus. (a) Normal group, (b) restraint stress group, (c) noise stress group, (d) cold stress group. The data are mean±standard error of the mean (S.E.M.). *p<0.05 compared to the normal group. **p<0.05 compared to the restraint stress group.
The time of performance and the error number were increased and the correct number was decreased in all stress groups relative to the normal group (p<0.05). The deteriorative effects of these stresses on spatial memory appeared to have the following rank order: restraint stress>noise stress>cold stress.
2. Effects of various types of stress on cell proliferation in the hippocampal dentate gyrus
Photomicrographs of BrdU-positive cells in the hippocampal dentate gyrus in each group are presented in Fig. 2A. The number of BrdU-positive cells was 624.32±30.88/mm
2in the normal group, 281.10±14.87/mm
2in the restraint stress group,
338.72±20.86/mm
2in the noise stress group, and 446.91±
19.68/mm
2in the cold stress group.
Photomicrographs of Ki-67-positive cells in the hippocampal dentate gyrus in each group are presented in Fig. 2B. The number of Ki-67-positive cells was 44.44±4.14/mm
2in the normal group, 16.94±1.98/mm
2in the restraint stress group, 16.74±1.53/mm
2in the noise stress group, and 34.93±2.68/mm
2in the cold stress group.
Cell proliferation in the hippocampal dentate gyrus was decreased in all stress groups relative to the normal group (p
<0.05). The suppressing effect of these stresses on cell prolife-
ration in the hippocampal dentate gyrus appeared to have the
following rank order: restraint stress>noise stress>cold stress.
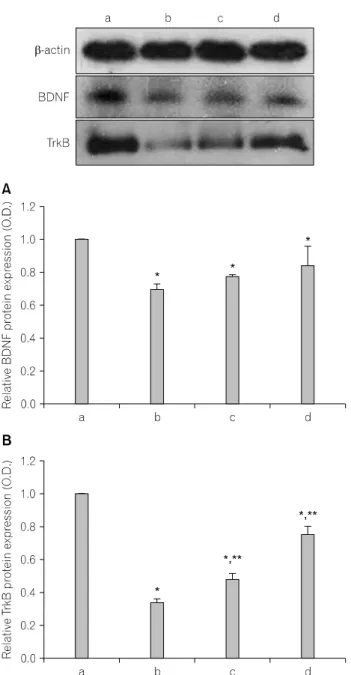
Fig. 3. The effects of various types of stress on the BDNF and TrkB