447
Copyright © 2015 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
동물의장관에서식하고있는고유한장내미생물의대부분은 숙주에유익한영향을미치거나무해하다
.
일반적으로장내미 생물은장관으로침입하는병원체로부터숙주를보호하며,
섭 취된음식으로부터영양성분과에너지를얻고정상적인면역 기능을수행하는데도움을주는것으로알려져있다(Sonnen- burg et al., 2005; Candela et al., 2008; Fukuda et al., 2011;
Olszak et al., 2012; Yatsunenko et al., 2012).
최근장내미생물 이숙주의체중증가또는비만에미치는영향을이해하기위한 연구가많이진행되고있다(Ley et al., 2006; Angelakis et al.,
2012).
또한차세대염기서열분석방법을이용한장내미생물생태계의특징과이해를위한연구가매우활발히수행되고있 지만
(Eckburg et al., 2005),
아직도많은부분이이해되지않고있다
.
사실어류에서도장내미생물의중요성이점점인식되고 있지만,
사람이나다른포유동물에비해서는그와관련된연구 가매우드문편이다(Gatesoupe, 2010).
다른동물과마찬가지 로장내마이크로바이옴이어류에미치는영향을이해하기위 해서는어떤종류의고유한미생물이어류의장관내에서식하 는지를이해하는것이우선시되어야할것이다.
그럼에도불구 하고우리나라최대양식어종인넙치의장내세균의다양성에 관한연구는매우제한적이다(Sugita et al., 2002; Sugita and Ito, 2006; Kim and Kim, 2013).
이번연구의목적은배양되 는넙치장내미생물다양성이해를위한최적의배양조건을탐 색하고,
그조건에따른미생물다양성을비교하며16S rRNA gene clone library
를제작하여비배양의존적방법을이용한미 생물의다양성을이해하고자하였다.
넙치(Paralichthys olivaceus) 장관의 배양 및 비배양 방법에 의한 세균의 다양성
김아란·김도형*
부경대학교 수산생명의학과
Diversity of Cultured and Uncultured Bacteria in the Gut of Olive Flounder Paralichthys olivaceus
Ahran Kim and Do-Hyung Kim*
Department of Aquatic Life Medicine, Pukyong National University, Busan 48513, Korea
We determined the optimal culture conditions for obtaining the maximum number of intestinal bacteria from the olive flounder Paralichthys olivaceus , and studied bacterial diversity using both culture-dependent and culture-independent methods. Using six culture conditions, mean bacterial numbers were greater than 10
6per gram of gut mucus, regard- less of the medium. However, the bacterial diversity, based on colony morphology, appeared much higher on Marine agar (MA) and Zobell 2216 agar than on other media. We found eight and 17 cultured bacterial phylotypes with 99%
minimum similarity in gut mucus grown on MA and tryptic soy agar, respectively. Furthermore, we used genomic DNA extracted from gut mucus to generate 78 random clones, which were grouped into 25 phylotypes. Of these, six were affiliated with Firmicutes, Actinobacteria, and Verrucomicrobia, and were not found using our culture- dependent methods. Consequently, we believe that Marine agar and Zobell 2216 agar are optimal media for culturing diverse intestinal microbes; we also discovered several novel sequences not previously recognized as part of the gut microbiota of olive flounder.
Key words: Olive flounder, Microbial diversity, 16S rRNA gene, Fish disease, Intestine
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2015.0447
Korean J Fish Aquat Sci 48(4) 447-453, August 2015 Received 9 July 2015; Revised 31 July 2015; Accepted 4 August 2015
*Corresponding author: Tel: +82. 51. 629. 5945 Fax: +82. 51. 629. 5938
E-mail address: [email protected]
재료 및 방법
넙치 장관으로부터 배양되는 세균의 분리 및 계수
이번연구에서는배합사료만급여한양식넙치(Paralichthys olivaceus) (n=10;
평균체중=100 g)
를사용하였다.
배양되는 세균의분리를위해서넙치5
마리로부터무균적으로장관을적 출해세로방향으로길게잘라배설물과유사한장내용물을제 거한후장점액만을1.5 mL microcentrifuge tube
에모았다.
이 를멸균생리식염수에균질화한후단계희석(10
-2과10
-3)
하여, 0.5%
의NaCl
이첨가된brain heart infusion agar [BHIA(0.5)], 1.5%
의NaCl
이첨가된BHIA [BHIA(1.5)], 1.5%
의NaCl
과0.3%
의yeast extract
가첨가된tryptic soy agar (TNY), Marine agar (MA), Zobell 2216 agar (ZA)
와thiosulfate citrate bile salts sucrose agar (TCBS)
에100 μL
를도말한후25℃
에서배 양하였다. 7
일후에배지에자란세균집락을계수하였고,
집락 의성상과그람염색을통해배양되는 세균의다양성을 측정 하였다.
구분된각각의세균들을순수분리하여tryptone soya broth (TSB; Oxoid)
에배양후glycerol
을첨가하여사용하기 전까지-70℃
에보관하였다.
세균의 genomic DNA 분리
Marine agar
와TNY (1.5%
의NaCl
과0.3%
의yeast extract
가 첨가된TSA)
에서분리된세균들을25℃
에서24
시간동안배 양한이후3,000 g
에10
분동안원심분리한후genomic DNA
를추출하는데사용하였고, 16S rRNA gene clone library
제작 을위해5
마리의넙치의장점액으로부터DNA
를분리하였다. DNA
분리에는DNeasy
®Blood and Tissue kit (Qiagen)
가사 용되었으며제조사의실험방법을따랐다.
Polymerase chain reaction (PCR)
각분리균주와장점액으로부터추출한
DNA
를Table 1
에제 시된프라이머를이용하여증폭시켰다. PCR
의조건은다음과 같이수행하였다; 94℃
에서3
분동안initial denaturation
을수 행하고, 94℃
에서30
초, 56℃
에서30
초, 72℃
에서1
분의과정 을30
회반복한후, 72℃
에서7
분간final extension
하였다.
증 폭산물은ethidium bromide (1 ng/mL)
가첨가된1% (w/v) agarose gel
상에서전기영동하여분석하였다.
16S rRNA clone library 제작
장점액으로부터추출한
DNA
를9F
와1492R primer (Table 1)
를 사용하여PCR
로 증폭시켰다.
정제된PCR product
를pGEM
®-T vector (Promega)
와E. coli XL1-blue
를이용하여 클로닝하였다. DNA
농도는insert:vector
가3:1
의 비율이 되 도록32.5 ng/μL
로조정하였다. 2× Rapid Ligation Buffer 5 μL, pGEM
®-Teasy Vector (50 ng) 1 μL, T4 DNA Ligase (3 Weiss units/μL) 1 μL, PCR product 2 μL
와11 μL
의멸균Mil- liQ water
를넣은혼합물을4℃
에overnight
하였다. Positive control
로insert DNA
를2 μL
를넣어사용하였다.
각ligation reaction
의2 μL
를멸균microcentrifuge tube
에넣고, 50
또는100 μL (positive control
의경우)
의competent cell
을tube
에옮 겨20
분동안얼음위에꽂아둔후, 42℃
로맞춰진항온수조에1
분동안heat shock
을주었다.
이후즉시얼음위로옮겨2
분동 안반응을정지시켰고, 950
또는900 μL
의SOC
배지를첨가 한후37℃
에서1.5
시간동안진탕배양하였다(~150 rpm).
이후 세포들을1,000 g
에10
분동안원심분리하여집균하였고, 200 μ L
의SOC
배지에재현탁하여그중100 μL
를ampicillin (
최 종농도100 μg/μL, Sigma), 100 μL
의100 mM IPTG
와20 μL
의X-gal
이미리도말된LB (Luria-Bertani) agar
에도말하였 고2
반복으로수행하였다. 37℃,
암실에서24
시간동안배양한 후자란흰색집락만을selection
하여24
시간LB broth
에배양 한이후plasmid DNA
를AccuPrep
®Plasmid mini extraction kit (Bioneer, Daejeon, Korea)
를사용하여제조사의절차에따 라분리하였다.
염기서열 분석
배양된세균과
insertion
을가진plasmid DNA
의정제된PCR product
의sequencing reaction
은이전에서술되었던것과같 이Big Dye Terminator cycle sequencing kit
와ABI Prism 310 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA)
를 사용하여 수행하였다(Han et al., 2011). DNA se- quence
의quality
확인을 위하여Bioedit (version 7.0.9.0)
과Genetyx version 7.0.3 packed program (Software Develop- ment Co., Tokyo, Japan)
을사용하였다.
또한, 16S ribosomal DNA clone library
의염기서열은online tool
인DECIPHER (Wright et al., 2012)
를 사용하여chimeric construct
의유무 를 확인하였고, 3′
과5′
말단에있는부정확한 염기서열또한Table 1. Primers used for PCR and sequencing in this study
Primer designation Sequence (5’→3’) Use(s) Reference(s)
9F GAGTTTGATCCTGGCTCAG
Cloning Lane (1991)
1492R GG(C/T)TACCTTGTTACGACTT
63F CAGGCCTAACACATGCAAGTC
Sequencing Marchesi et al. (1998)
1387R GGGCGG(A/T)GTGTACAAGGC
제거하였다
. 1%
이하의염기서열상동성을가진경우동일한phylotype (operational taxonomic units)
으로 간주하여 분석 을진행하였고,
각phylotype
과가장근접한표준균주를결정 하기위해RDP SEQMATCH program (http://rdp.cme.msu.
edu/index.jsp) (Cole et al., 2013)
을이용하였다.
계통수제작에 는ClustalX (version 1.83)
와MEGA6(Saitou and Nei, 1987;
Tamura et al., 2011)
프로그램이이용되었으며, Kimura-2-pa- rameter distances
에기초하여neighbor-joining tree
를만들었 으며1,000
번의re-sampling
을통해bootstrap value
를결정하 였다.
결과 및 고찰
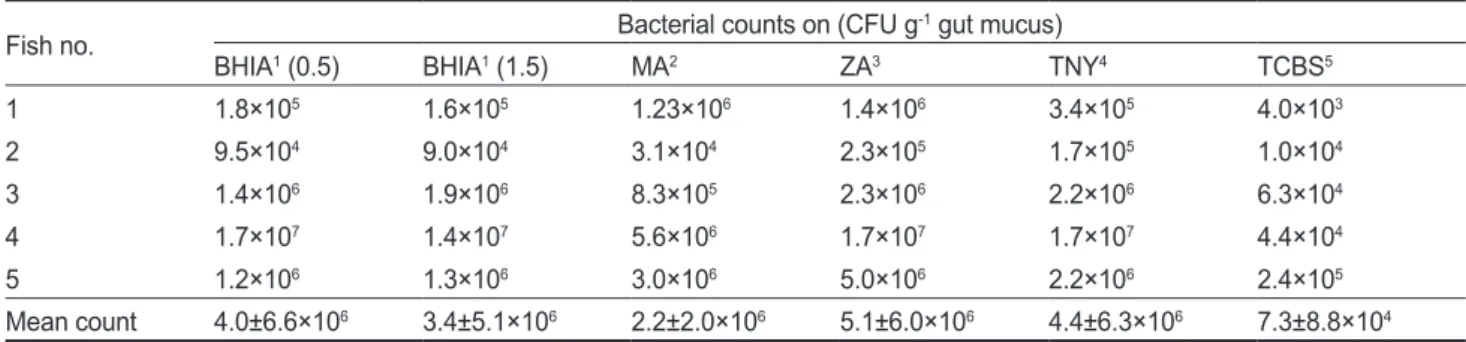
넙치 장관의 배양되는 세균의 수
넙치장점액분리후단계희석하여
BHIA (0.5), BHIA (1.5), TNY, MA, ZA, TCBS
에배양한결과를Table 2
에나타내었 다. TCBS
를제외한모든배지에서10
4-10
7CFU/g gut mucus
의세균이배양되었으며, TCBS
에서는7.3±8.8×10
4CFU/g gut mucus
으로나타나배양되는세균중에서Vibrio
과에속 하는세균이많이존재하고있음을확인할수있었다(Table 2).
이결과는이전넙치의연구결과
(Kim and Kim, 2013)
와매 우유사하였다. Senegales sole Solea senegalensis
의장내세균 은MA
에서2.3×10
5-6.7×10
6CFU/g, TCBS
에서5.0×10
3- 3.9×10
5CFU/g
의수로배양되는것으로나타났으며(Martin- Antonio et al., 2007),
그외grouper Epinephelus coioides (Sun et al., 2009)
와rainbow trout Oncorhynchus mykiss (Kim et al., 2007)
도약10
6CFU/g gut content
로나타나어종이나서 식지와관계없이배양되는세균의수는유사한것으로나타났 다.
그러나이번연구결과와같이이전연구에서도넙치의개체 에따라배양되는세균의수에큰차이가났는데일부개체에 서10
8CFU/g
이상의세균이분리되기도하였다(Sugita et al., 2002; Sugita and Ito, 2006). Sugita and Ito (2006)
의연구에서 는배지의구성성분이장내세균의배양에영향을미칠수있음 을의미하나,
이번연구에서는TCBS
를제외한다양한배지에서배양되는세균의수에큰차이가나지않아넙치장내세균의 배양에배지성분이큰영향을끼치지는않은것으로사료된다
. 넙치 장관의 배양되는 세균 종
넙치의장내세균의다양성에 관한이전연구가존재하지만 매우드물다
(Sugita and Ito, 2006; Kim and Kim, 2013).
이 번연구에서는6
종류의배지를이용하여넙치장내세균의다 양성연구에최적인배지를탐색하고자하였으며,
그중4
종류 의배지에서관찰된결과를Table 3
에나타내었다.
분별된세 균집락의종류는Marine agar
와Zobell 2216 agar
에서가장 많았으며,
개체에따라서로1
또는2
개정도의차이밖에나지 않아그다양성은유사하였다. BHIA
와TNY
배지에서는MA
와ZA
와달리다양한세균집락이배양되지않았다.
유전적으 로99%
이상의상동성을가지는경우동일한phylotype
으로 간주하였을때, TNY
와MA
에서각각8
과17
개의phylotype
이나타났으며그결과를Fig. 1
에계통수로나타내었다. TNY
배지에서분리된8
개의phylotype
은모두Proteobacteria
그룹 에포함되었으며,
이중6
개가Vibrionaceae,
나머지는Rhodo- bacteraceae
와Flavobacteriaceae
에각각속하였다(Fig. 1a).
이 에반해MA
배지에서는TNY
에서보다더다양한세균이분리 되었다(Fig. 1b). 17
개의분리된세균중Proteobacteria
그룹 에속하는phylotype
은12
개로우점적이었으며, Vibrionaceae
와Pseudoalteromonadaceae
에각각4
개와2
개의phylotype
이Table 3. Differential colony counts appeared on varying media Fish no. Marine agar Zobell 2216
agar Brain heart
infusion agar1 Tryptic soy agar2
1 10 10 5 6
2 2 3 1 2
3 6 5 3 3
4 7 6 2 2
5 10 8 2 2
1supplemented with 1.5% NaCl; 2 supplemented with 1.5% NaCl and 0.3% yeast extract
Table 2. Microbial colony counts of gut microbes grown on varying culture media
Fish no. Bacterial counts on (CFU g-1 gut mucus)
BHIA1 (0.5) BHIA1 (1.5) MA2 ZA3 TNY4 TCBS5
1 1.8×105 1.6×105 1.23×106 1.4×106 3.4×105 4.0×103
2 9.5×104 9.0×104 3.1×104 2.3×105 1.7×105 1.0×104
3 1.4×106 1.9×106 8.3×105 2.3×106 2.2×106 6.3×104
4 1.7×107 1.4×107 5.6×106 1.7×107 1.7×107 4.4×104
5 1.2×106 1.3×106 3.0×106 5.0×106 2.2×106 2.4×105
Mean count 4.0±6.6×106 3.4±5.1×106 2.2±2.0×106 5.1±6.0×106 4.4±6.3×106 7.3±8.8×104
1brain heart infusion agar supplemented with 0.5% or 1.5% NaCl; 2marine agar; 3Zobell 2216 agar; 4tryptic soy agar supplemented with 1.5%
NaCl and 0.3% yeast extract; 5thiosulfate citrate bile salts sucrose agar
포함되었고
, Shewanellaceae, Colwelliaceae, Francisellaceae, Sphingomonadaceae, Rhodobacteraceae
에각각하나의phy- lotype
이속하였다.
나머지5
개의phylotype
은TNY
에서배양 된세균에서는관찰되지않은Bacteroidetes
그룹에속하였으 며,
이중4
개와1
개의phylotype
이각각Flavobacteriaceae
와Flammeovirgaceae
에포함되었다.
일반적으로TNY
와BHIA, nutrient agar
는기본영양배지로세균배양시많이사용되지 만,
배양되는 장내세균의다양성연구에는Marine agar
또는Zobell 2216 agar
가가장적합한것을확인하였다.
16S rRNA gene clone library의 염기서열 분석으 로 확인된 세균 종
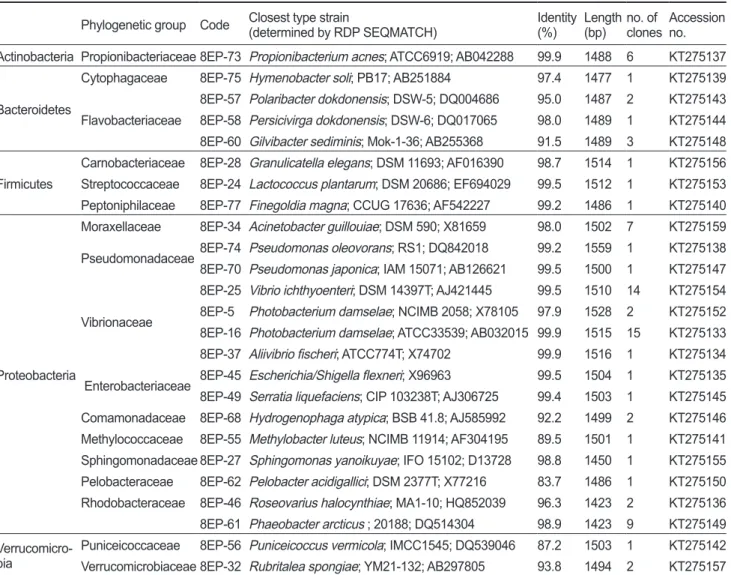
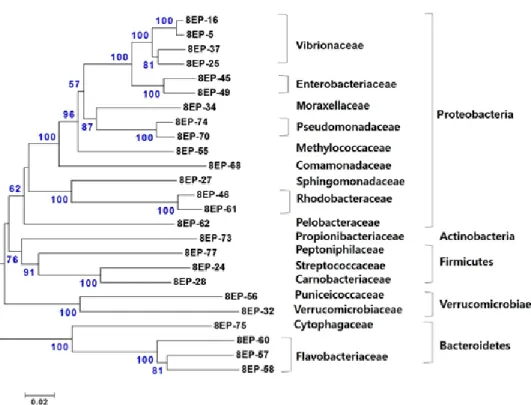
이번연구에서는총
78
개clone
의염기서열을분석하여25
개 의phylotype
을얻었으며가장근접한세균종(
기존의염기서열 이밝혀진표준균주)
의결과를Table 4
에나타내었다.
결론적으 로clone library
에서도출된많은염기서열은이전에어류의장 내세균으로는알려진적이없는새로운세균종의서열임이드러났다
.
이25
개의phylotype
은Actinobacteria, Bacteroidetes, Firmicutes, Proteobacteria
및Verrucomicrobia
에속하였으며, phylotype
의대부분(
약76%)
이Proteobacteria
로가장많은비 율을차지하였다(Fig. 3). Proteobacteria
그룹중에서는Vibrio- naceae
가가장우점적(
약55%)
이었으며,
다음으로Rhodobac- teraceae, Moraxellaceae
가각각약19%
와12%
의비율을차지 하였다(Fig. 3).
이외에도Enterobacteriaceae, Pseudomonada- ceae, Moraxellaceae, Methylococcaceae, Comamonadaceae, Sphingomonadaceae
에 속하는 세균들이 분리되었다(Fig. 2 and Table 4).
양식산과자연산넙치장관의세균다양성을조 사한 연구에서각각38
개와47
개의phylotype
이분리되었고Firmicutes, Proteobacteria, Actinobacteria
와Bacteroidetes
그 룹에속하였다(Kim and Kim, 2013).
이중Proteobacteria
가양 식산어류에서60.5%,
자연산어류에서는51.1%
로가장우점 적이었으며,
이번연구에서처럼Vibrionaceae
가가장높은비 율을차지하였다(Kim and Kim, 2013).
Fig. 1. Neighbor-joining phylogenetic tree showing the relationship of 8 and 17 16S rRNA gene sequences of cultured bacteria grown on tryptic soy agar (supplemented with 1.5% NaCl and 0.3% yeast extract) (a) and on marine agar (b), respectively, retrieved from gut mucus of olive flounder Paralichthys olivaceus. Bootstrap values based on 1,000 re-samplings display the significance of the interior nodes, and are shown at branch points; only values displaying >50% are given. The scale bar represents a 2% estimated difference in nucleotide sequences.
이번 연구에서는
Vibrio ichthyoenteri
와Photobacterium damselae
와같은어류병원체가매우높은비율(
각각14/78
개 와15/78
개의clone)
로분리되었다.
일반적으로이러한병원체 들은어류의장관에서분리•
배양된다는이전의연구결과(Su- gita and Ito, 2006)
와일치함을보여주며,
어류의장관에존재 하다가면역력이약해진틈을타서체내에침투하는것으로생 각된다.
Actinobacteria
에속하는6
개의clone
은모두Propionibacte- riaceae
의Propionibacterium acnes
로확인되었고, Firmicutes
그룹에서는3
개의clone
이 각각Carnobacteriaceae, Strepto- coccaceae, Peptoniphilaceae
에속하였다.
그외Bacteroidetes
에는Cytophagaceae
와Flavobacteriaceae, Verrucomicrobia
에 는Puniceicoccaceae
와Verrucomicrobiaceae
에속하는세균이 확인되었다.
이상의결과를종합해본결과넙치의배양되는세균의다양
성연구에는
Marine agar
또는Zobell 2216 agar
가가장적합 한것으로나타났다.
그러나배양의존적인방법은특별한배양 환경및조건이요구되어배양이까다로운미생물이거나절대 혐기성미생물의배양이배제되기때문에한계가있다(Asfie et al., 2003).
이번연구에서는5
마리의넙치를배양되는세균과 또다른5
마리의넙치를clone library
제작에사용하여배양의 존적인방법과비배양의존적인방법의결과를직접적으로비 교하는것은의미가없다.
그렇지만clone library
에서더많은phylotype
이도출되었고이들중많은염기서열이이전연구결과에서드러나지않았던새로운종임이밝혀졌으므로최신의 배양비의존적방법은미생물의다양성연구에매우유용한것 임을다시한번확인할수있었다
.
최근들어차세대염기서열 분석방법(next-generation sequencing)
을적용한다양한연구 가진행되고있어이기술을적용한어류의장내세균다양성및 기능성연구가필요할것으로생각된다.
Table 4. Representative sequences of a clone library isolated from the intestinal mucus of farmed olive flounder Paralichthys olivaceus Phylogenetic group Code Closest type strain
(determined by RDP SEQMATCH) Identity (%) Length
(bp) no. of
clones Accession no.
Actinobacteria Propionibacteriaceae 8EP-73 Propionibacterium acnes; ATCC6919; AB042288 99.9 1488 6 KT275137
Bacteroidetes
Cytophagaceae 8EP-75 Hymenobacter soli; PB17; AB251884 97.4 1477 1 KT275139
Flavobacteriaceae
8EP-57 Polaribacter dokdonensis; DSW-5; DQ004686 95.0 1487 2 KT275143 8EP-58 Persicivirga dokdonensis; DSW-6; DQ017065 98.0 1489 1 KT275144 8EP-60 Gilvibacter sediminis; Mok-1-36; AB255368 91.5 1489 3 KT275148
Firmicutes
Carnobacteriaceae 8EP-28 Granulicatella elegans; DSM 11693; AF016390 98.7 1514 1 KT275156 Streptococcaceae 8EP-24 Lactococcus plantarum; DSM 20686; EF694029 99.5 1512 1 KT275153 Peptoniphilaceae 8EP-77 Finegoldia magna; CCUG 17636; AF542227 99.2 1486 1 KT275140
Proteobacteria
Moraxellaceae 8EP-34 Acinetobacter guillouiae; DSM 590; X81659 98.0 1502 7 KT275159 Pseudomonadaceae8EP-74 Pseudomonas oleovorans; RS1; DQ842018 99.2 1559 1 KT275138 8EP-70 Pseudomonas japonica; IAM 15071; AB126621 99.5 1500 1 KT275147
Vibrionaceae
8EP-25 Vibrio ichthyoenteri; DSM 14397T; AJ421445 99.5 1510 14 KT275154 8EP-5 Photobacterium damselae; NCIMB 2058; X78105 97.9 1528 2 KT275152 8EP-16 Photobacterium damselae; ATCC33539; AB032015 99.9 1515 15 KT275133 8EP-37 Aliivibrio fischeri; ATCC774T; X74702 99.9 1516 1 KT275134 Enterobacteriaceae 8EP-45 Escherichia/Shigella flexneri; X96963 99.5 1504 1 KT275135 8EP-49 Serratia liquefaciens; CIP 103238T; AJ306725 99.4 1503 1 KT275145 Comamonadaceae 8EP-68 Hydrogenophaga atypica; BSB 41.8; AJ585992 92.2 1499 2 KT275146 Methylococcaceae 8EP-55 Methylobacter luteus; NCIMB 11914; AF304195 89.5 1501 1 KT275141 Sphingomonadaceae 8EP-27 Sphingomonas yanoikuyae; IFO 15102; D13728 98.8 1450 1 KT275155 Pelobacteraceae 8EP-62 Pelobacter acidigallici; DSM 2377T; X77216 83.7 1486 1 KT275150 Rhodobacteraceae 8EP-46 Roseovarius halocynthiae; MA1-10; HQ852039 96.3 1423 2 KT275136 8EP-61 Phaeobacter arcticus ; 20188; DQ514304 98.9 1423 9 KT275149 Verrucomicro-
bia Puniceicoccaceae 8EP-56 Puniceicoccus vermicola; IMCC1545; DQ539046 87.2 1503 1 KT275142 Verrucomicrobiaceae 8EP-32 Rubritalea spongiae; YM21-132; AB297805 93.8 1494 2 KT275157
사 사
이논문은부경대학교자율창의학술연구비
(2014
년)
에의하 여연구되었습니다.
References
Angelakis E, Armougom F, Million M and Raoult D. 2012.
The relationship between gut microbiota and weight gain in humans. Future Microbiol 7, 91-109. http://dx.doi.
org/10.2217/fmb.11.142.
Asfie M, Yoshijima T and Sugita H. 2003. Characterization of the goldfish fecal microflora by the fluorescent in situ hybridization method. Fish Sci 69, 21-26. http://dx.doi.
org/10.1046/j.1444-2906.2003.00583.x.
Candela M, Perna F, Carnevali P, Vitali B, Ciati R, Gionchetti P, Rizzello F, Campieri M and Brigidi P. 2008. Interaction of probiotic Lactobacillus and Bifidobacterium strains with hu- man intestinal epithelial cells: adhesion properties, competi- tion against enteropathogens and modulation of IL-8 pro- duction. Int J Food Microbiol 125, 286-292. http://dx.doi.
org/10.1016/j.ijfoodmicro.2008.04.012.
Cole JR, Wang Q, Fish JA, Chai B, McGarrell DM, Sun Y, Brown CT, Porras-Alfaro A, Kuske CR and Tiedje JM.
Fig. 2. Neighbor-joining phylogenetic tree showing the relationship of 25 16S rRNA gene sequences derived from gut mucus (clone library).
Bootstrap values based on 1,000 re-samplings display the significance of the interior nodes, and are shown at branch points; only values displaying >50% are given. The scale bar represents a 2% estimated difference in nucleotide sequences.
Fig. 3. Clonal frequency of microbial phylogenetic groups in a 16S rRNA gene clone library established using PCR amplification, cloning, and sequencing of 16S rRNA genes extracted from the gut mucus of olive flounder Paralichthys olivaceus. (a) phylum-level microbial composition; (b) family-level microbial composition of Proteobacteria.
2013. Ribosomal Database Project: data and tools for high throughput rRNA analysis. Nucl Acids Res 42,633-642.
http://dx.doi.org/10.1093/nar/gkt1244.
Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE and Relman DA. 2005. Di- versity of the human intestinal microbial flora. Science 308, 1635-1638. http://dx.doi.org/ 10.1126/science.1110591.
Fukuda S, Toh H, Hase K, Oshima K, Nakanishi Y, Yoshimura K, Tobe T, Clarke JM, Toppng DL, Suzuki T, Taylor TD, Itoh K, Kikuchi J, Morita H, Hattori M and Ohno H. 2011.
Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 469, 543-547. http://
dx.doi.org/10.1038/nature09646.
Gatesoupe FJ. 2010. Probiotics and other microbial manipula- tions in fish feeds: prospective health benefits. In: Bioactive foods in promoting health: probiotics and prebiotics. Wat- son W and Preedy V R, eds. Academic Press, San Diego, U.S.A., 541-552.
Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/
NT. Nucleic Acids Symp 41, 95-98.
Han HJ, Kim DY, Kim WS, Kim CS, Jung SJ, Oh MJ and Kim DH. 2011. Atypical Aeromonas salmonicida infection in the black rockfish, Sebastes schlegeli Hilgendorf, in Ko- rea. J Fish Dis 34, 47-55. http://dx.doi.org/10.1111/j.1365- 2761.2010.01217.x.
Kim DH, Brunt J and Austin B. 2007. Microbial diversity of in- testinal contents and mucus in rainbow trout(Oncorhynchus
mykiss). J Appl Microbiol 102, 1654-1664. http://dx.doi.
org/10.1111/j.1365-2672.2006.03185.x.
Kim DH and Kim DY. 2013. Microbial diversity in the intes- tine of olive flounder (Paralichthys olivaceus). Aquacul- ture 414, 103-108. http://dx.doi.org/10.1016/j.aquacul- ture.2013.06.008.
Lane DJ. 1991. 16S/23S rRNA sequencing. Nucleic acid tech- niques in bacterial systematics. Stackebrandt E. and Good- fellow M, eds, Academic Press, Chichester, UK, 115-175.
Ley RE, Turnbaugh PJ, Klein S and Gordon JI. 2006. Microbial ecology: human gut microbes associated with obesity. Na- ture 444, 1022-1023. http://dx.doi.org/10.1038/4441022a.
Marchesi JR, Sato T, Weightman AJ, Martin TA, Fry JC, Hiom SJ and Wade WG. 1998. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl Environ Microbiol 64, 795- Martin-Antonio B, Manchado M, Infante C, Zerolo R, Labella 599.
A, Alonso C and Borrego JJ. 2007. Intestinal microbiota variation in Senegalese sole (Solea senegalensis) under dif- ferent feeding regimes. Aquacult Res 38, 1213-1222. http://
dx.doi.org/10.1111/j.1365-2109.2007.01790.x.
Olszak T, An D, Zeissig S, Vera MP, Richter J, Franke A, Glick- man JN, Siebert R, Baron RM, Kasper DL and Blumberg
KR. 2012. Microbial exposure during early life has persis- tent effects on natural killer T cell function. Science 336, 489-493. http://dx.doi.org/10.1126/science.1219328.
Saitou N and Nei M. 1987. The neighbor-joining method-a new method for reconstructing phylogenetic trees. Mol Biol Evol 4, 406-425.
Sonnenburg J L, Xu J, Leip DD, Chen CH, Westover BP, Weath- erford J, Buhler JD and Gordon JI. 2005. Glycan foraging
in vivo by an intestine-adapted bacterial symbiont. Science
307, 1955-1959. http://dx.doi.org/10.1126/science.1109051.Sugita H, Okano R, Suzuki Y, Iwai D and Kanagawa F. 2002.
Antibacterial abilities of intestinal bacteria from larval and juvenile Japanese flounder against fish pathogens.
Fish Sci 68, 1004-1011. http://dx.doi.org.10.1046/j.1444- 2906.2002.00525.x
Sugita H. and Ito Y. 2006. Identification of intestinal bacteria from Japanese flounder (Paralichthys olivaceus) and their ability to digest chitin. Lett Appl Microbiol 43, 336-342.
http://dx.doi.org/10.1111/j.1472-765X.2006.01943.x.
Sun Y, Yang H, Ling Z, Chang J and Ye J. 2009. Gut microbiota of fast and slow growing grouper Epinephelus coioides. Afr J Microbiol Res 3, 713-720.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M and Ku- mar S. 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28, 2731- 2739. http://dx.doi.org/10.1093/molbev/msr121.
Wright ES, Yilmaz LS and Noguera DR. 2012. DECIPHER, a search-based approach to chimera identification for 16S rRNA sequences. Appl Environ Microbiol 78, 717-725.
http://dx.doi.org/10.1128/AEM.06516-11.
Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominquez-Bello MG, Contreras M, Margris M, Hidalgo G, Baldassano RN, Anokhin AP, Heath AC, Warner B, Reeder J, Kuczynski J, Caporaso JG, Lozupone CA, Lauber C, Clemente JC, Knights D, Knight R and Gordon JI. 2012. Human gut mi- crobiome viewed across age and geography. Nature 486, 222-227. http://dx.doi.org/10.1038/nature11053.