397
Copyright © 2018 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
1920
년대항생물질이발견된이래로항생제는질병의예방이 나치료목적으로사용되어져왔으나,
항생제의오·
남용에의하 여발생한항생제내성균은질병치료의어려움및약물잔류에 의한안전성문제등을야기하였고,
최근사회적문제로서대두 되고있다.
특히,
동물용항생제는감염성질환의예방이나치 료목적외에도사료와혼합하여가축이나어류의성장촉진을 목적으로도사용되어졌으며(Lee et al., 2010a),
이와같은무분 별하고광범위한항생제사용으로인하여내성균의출현이라는 문제가발생하고있다.
Animal and Plant Quarantine Agency (APQA, 2009)
에따 르면erythromycin (Em)
은어류양식산업에서6.3%
의비중을 차지하며, oxytetracycline
에이어높은사용률이확인되었다. Em
은macrolide
계항생제로서,
세균의50S ribosome
에가역적으로결합하여세균의
ribonucleic acid (RNA)
의존성단백 합성을억제함으로서작용한다(Hansen et al., 1999). Em
내성 기전은크게두가지로밝혀져있으며,
내성유전자또한내성기 작에따라구분된다.
첫번째는methylase
를통해세균의ribo- some
에항균제가결합하는부위를변화시키는기작으로, erm (erythromycin ribosome methylation)
유전자와연관되어있으 며,
두번째로내재성막단백질에의해일어나는efflux pump
에의한것으로mef (A) (macrolide efflux), msr (A) (macrolide and streptogramine B), vag (virginiamycin factor)
유전자와관 련이있다(Lee and Kim, 2013).
내성기작의특성에따라Em
내 성유전자도그종류를달리하는데rRNA methylase
의경우는erm (A), erm (B), erm (C)
등이존재하는것으로알려져있으 며, efflux
내성기작을가지는유전자는mef (A)
가대표적인것 으로보고되었다(Sapkota et al., 2006).
최근들어
macrolide
계항생제역시사용량이증가함에따라제주지역 양식 넙치(Paralichthys olivaceus)에서 분리한 어병세균 내 Erythromycin 내성 유전자 분석
이다원·전려진·김승민
1·정준범*
제주대학교 해양의생명과학부, 1주식회사 우진비앤지
Analysis of Erythromycin Resistance Gene in Pathogenic Bacteria Iso- lates from Cultured Olive flounder Paralichthys olivaceus in Jeju
Da Won Lee, Lyu Jin Jun, Seung Min Kim
1
and Joon Bum Jeong*School of Marine Biomedical Sciences, Jeju National University, Jeju 63243, Korea
1Central Research Institute, Woo Gene B&G, Hwaseong 48513, Korea
We determined the resistance rates of pathogenic bacteria isolated from cultured olive flounder Paralichthys oliva- ceus to erythromycin (Em), antibiotic typically used in aquaculture and analyzed the genotypes of resistant bacteria using polymerase chain reaction (PCR). We isolated and utilized 160 isolates of Streptococcus parauberis , 1 of S.
iniae , 66 of Edwardsiella tarda , 56 of Vibrio sp. and 23 of unidentified bacteria from presumed infected olive flounder from Jeju Island from March 2016 to October 2017. Of the 306 isolated strains, Em-resistant strains included 33 of S. parauberis , 39 of E. tarda and 2 of Vibrio sp. We conducted PCR to assess the resistance determination of Em- resistant strains. Five different types of Em-resistance genes were detected in the 74 Em-resistant strains: erm (A), erm (B), erm (C), mef (A) and mef (E); erm (A) and erm (B) were detected in 1 (3%) and 24 (72.7%) S. parauberis isolates, respectively. In E. tarda , erm (B) was detected in five isolates (12.8 %) and no Em-resistance genes were detected in the two Vibrio sp. isolates.
Key words: Erythromycin, Olive flounder, Resistance gene, Jeju, erm (B)
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
https://doi.org/10.5657/KFAS.2018.0397 Korean J Fish Aquat Sci 51(4) 397-403, August 2018
Received 2 July 2018; Revised 5 August 2018; Accepted 14 August 2018
*Corresponding author: Tel: +82. 64. 754. 3426 Fax: +82. 64. 756. 3493
E-mail address: [email protected]
이들항생제에대한내성을가지는세균비율이증가하고있다
(Lee and Kim, 2013).
제주지역에서도분리한어병세균내항 생제내성률을비교한결과,
균주별로차이가있으나Em
에대 해높은내성률이확인되었다(Lee et al., 2016). 2016
년도Ko- rean statistical information service (KOSIS)
통계에따르면,
수 산식품수요증가에따른수산양식량은증가하고있으며,
항생 제사용량역시증가하고있는추세이다.
그에따라다양한균 주에서항생제내성양상에대한많은연구가이루어지고있으 나,
국내양식어류에서분리한Em
내성균및내성유전자에대 한연구는거의이루어지지않고있다(Han et al., 2012; Sung et al., 2013; Lee et al., 2016).
이에본연구에서제주지역에서 분리한어병세균내macrolide
계항생제인Em
에대해내성수 준을확인하고,
내성균주들이가지는내성유전자에대해분석 하고자하였다.
재료 및 방법
균주의 채집 및 분리
실험균주는
2016
년부터2017
년제주지역넙치양식장의병어로부터 분리하여 실험에 사용하였다
.
채집한 균주들의 동 정을 위해 시료의 내부 장기를증균배지인tryptic soy agar (TSA, Difco., USA)
및brain heart infusion agar (BHIA, Dif- co., USA)
와선택배지인thiosulfate citrate bile salts sucrose (TCBS) agar (Difco., USA), salmonella shigella (SS) agar (MB cell, Korea), glutamate starch phenol-red agar (GSP, Sigma-Aldrich, Korea), blood agar (KOMED, Korea)
배지에 도말하고, 27℃
에서18-24
시간배양하여균의집락형성및형 태를확인하였다.
분리된균주는추가적인실험에사용되기전 까지-70℃
에서보관하였다.
균주의 동정
분리한균주중선택배지
SS
배지상에서검은색집락을형 성하는균주를Edwardsiella tarda
균주로추정하였으며,
비브 리오선택배지인TCBS
배지상에서초록색또는노란색집락 을형성하는균주를Vibrio sp.
균주로추정하였다.
각균주들 의정확한균동정은Higene™ genemic DNA prep kit (BIO- FACT, Korea)
를사용하여균주들의genomic DNA
를추출한 후, Table 1
에제시한primer sets
를사용하여PCR
을통해확 인하였다.
Table 1. Primer sets used for the identification of bacteria in this study
Target pathogen Oligonucleotide sequences (5'-3') Product size (bp) Reference Streptococcus iniae AAGAGACGCAGTGTCAAAAG
220
Woo et al., 2006 CGTTTCTTATCTTGTTACTC
Streptococcus parauberis TCCAGTCTTTCGACCTTCTT CAAAGAGATGTTCGGCTTG 107
Lactococcus garvieae AAGCAGTCTTTTGATGCAAG ACTGTGCGCCCTTATTAACT 307
Edwardsiella tarda CGGTAAAGTTGAGTTTACGGGTG
415 Sakai et al., 2007 TGTAACCGTGTTGGCGTAAG
Vibrio genus GTCARATTGAAAARCARTTYGGTAAAGG 689
Kim et al., 2015 ACYTTRATRCGNGTTTCRTTRCC
Vibrio alginolyticus ACGGCATTGGAAATTGCGACTG TACCCGTCTCACGAGCCCAAG 199
Vibrio anguillarum GTTCATAGCATCAATGAGGAG 519 Demircan and Candan, 2006
GAGCAGACAATATGTTGGATG
Vibrio harveyi GTGATGAAGAAGCTTATCGCGATT
601 Kim et al., 2014 CGCCTTCTTCAGTTAACGCAGGA
Vibrio parahaemolyticus AGCTTATTGGCGGTTTCTGTCGG 297 Kim et al., 2015
CKCAACACCAAGAAAAGCCGTC Pseudomonas anguilliseptica GACCTCGCGCCATTA
438 Blanco et al., 2002 CTCAGCAGTTTTGAAAG
Flexibacter maritimus TGTAGCTTGCTACAGATGA 400 Cepeda et al., 2003
AAATACCTACTCGTAGGTACG
항생제 감수성 검사
Em
내성균분리및균주내항생제계열에따른내성비율확인 을위하여,
항생제감수성시험을clinical and laboratory stan- dards institute (CLSI, 2017)
기준에따라실시하였으며,
그방 법은다음과같다.
분리균의증균을위하여tryptic soy broth (TSB, Difco., USA)
를사용하였으며, 27℃
에서18-24
시간배 양하였다.
배양액은10
5-10
6으로희석하여실험에사용하였으 며,
멸균한면봉을이용해mueller hinton agar (MHA, Difco., USA)
평판에도말한후,
항생제디스크를부착시키고27℃
에 서24
시간배양하였다.
항생제디스크는모두Liofilchem®
에 서구입하였으며, amoxicillin (10 μg, AML), amoxicillin/cla- vulanic acid (30 μg, AUG), ampicillin (10 μg, AMP), chlor- amphenicol (10 μg, C), ciprofloxacin (5 μg, CIP), doxycycline (30 μg, DXT), enrofloxacin (5 μg, ENR), erythromycin (15 μg, E), gentamycin (10 μg, CN), kanamycin (30 μg, K), mi- nocycline (30 μg, MN), nalidixic acid (30 μg, NA), neomycin (30 μg, N), norfloxacin (10 μg, NOR), ofloxacin (5 μg, OFX), oxolinic acid (2 μg, OA), oxytetracycline (30 μg, OT), peni- cillin (10 μg, P), streptomycin (10 μg, S), sulfadiazine (300 μg, SUZ), tetracycline (30 μg, TE)
으로총21
종을사용하였으 며,
배양후증식억제대를확인하여내성균을분리하였다.
또한, Em
내성균의내성수준은broth dilution method
를적용한min- imum inhibitory concentration (MIC) test
를통해확인하였고, Lee et al. (2010b)
의방법과유사하게실시하여다음과같다.
항생물질의최고농도가100 μg/mL
가되도록하여최종농도 가0.78 μg/mL
가되도록mueller hinton broth (MHB, Difco., USA)
로1/2
씩단계희석하였으며,
균배양액의농도는10
5-10
6 이되도록희석하여준비하였다.
균주들의MIC
값은plate
를27℃
에서24
시간배양하여균의증식여부에따른액체배지의 혼탁도를확인하여균이자라지않은최소농도로결정하였다.
Erythromycin 내성유전자 검출
분리된
Em
내성균주들은erm (A), erm (B), erm (C), mef (A)
및mef (E)
를후보유전자로하여PCR
을통해존재여부 를살펴보았으며,
각내성관련유전자검출을위한primer
서 열은Table 2
에나타내었다. PCR
분석을위하여1 µM
의각primer, 2.5 µM
의각dNTP, 10x G-Taq Buffer, 2.5 U G-Taq DNA polymerase (Gene Pro Themal Cycler Cosmo)
및tem- plate DNA
를첨가한후, distilled water
로PCR
혼합물의최종volume
이20 μL
가되도록하였다. PCR
반응은94℃
에서3
분 간pre-denaturation
실시한후, 94℃ 30
초denaturation, 55℃
30
초annealing, 72℃ 30
초extenstion
반응을1 cycle
로하여30 cycles
한후, 72℃
에서7
분간post-extension
시켰다.
증폭 산물은1% agarose gel
상에서전기영동시킨후, UV
검출기(UVP, USA)
에서band
의크기를확인하여내성유전자의종류 를확인하였다.
결 과
세균의 분리 및 동정
2016
년3
월부터2017
년10
월까지제주지역넙치양식장에서 질병에감염된것으로추정되는넙치로부터총306
균주를분리 하여실험에사용하였다.
선택배지인SS, TCBS, GSP, Blood agar
를사용하여분리균을구분한결과, Streptococcus sp. 161
균주, E. tarda 66
균주, Vibrio sp. 56
균주,
그리고종이확인되지 않은23
균주를분리하였다. PCR
을통해연쇄구균병의원인균 을확인한결과, S. parauberis
가160
균주, S. iniae
가1
균주로확 인되었으며, Lactococcus garvieae
는검출되지않았다. E. tarda
균주는선택배지인SS
배지상에서검은색집락이형성된66
균 주를채집하였으며,
정확한동정을위해실시한PCR
분석에서 도동일한결과를보였다. Vibrioceae
의동정역시PCR
분석을 통해실시되었으며, 56
균주의Vibrio sp.
중V. alginolyticus
가Table 2. Primers and expected sizes of PCR products of various erm / mef genes
Gene Primer Oligonucleotide sequences (5’ to 3’) Product size (bp) Reference
erm (A) Em (A)F GTTCAAGAACAATCAATACAGAG 421
Jun, 2010
Em (A)R GCATCAGGAAAAGGACATTTTAC
erm (B) Em (B)F GAAAAGGTACTCAACCAAAT
Em (B)R AGTAACGGTACTTAAATTGT 639
erm (C) Em (C)F GCTAATATTGTTTAAATCGTCAATTCC 572
Em (C)R GGATCAGGAAAAGGACATTTTAC
mef (A) mef-F ATGGAAAAATACAACAATTGG
mefA-R GTAGTACAGCCATTCCTTC 646
mef (E) mef-F ATGGAAAAATACAAACAATTGG 812
mefE-R CCTATCAACATTCCAGATGC
PCR, polymerase chain reaction; erm, erythromycin ribosome methylation; mef, macrolide efflux.
4
균주, V. harveyi
가2
균주로확인되었다. 50
개의Vibrio
균주 는본연구에서동정을위해사용한primer sets
로는확인되지 않아V. alginolyticus, V. anguillarum, V. harveyi, V. parahae- molyticus
외다른Vibrio
균주로추정하였다.
또한, 306
균주중23
균주는본연구에서동정을위해사용한선택배지와primer sets
로는확인할수없어미동정균주로분류하였다.
제주도내양식장지역분포에따른균주발생차이확인을위 해시험균주들의지역별분포를확인하였으며
,
제주시에속해 있는양식현장(
한경,
애월,
조천,
구좌등)
에서전체306
균주중 총99
균주,
서귀포시에위치하고있는양식현장(
대정,
남원,
표 선,
성산등)
에서S. parauberis 119
균주, S. iniae 1
균주, E. tarda 46
균주, Vibrio sp. 38
균주,
미동정균주3
균주로207
균주가분 리되었음을확인하였다.
Erythromycin 내성균주 분리
Em
내성균주는약제감수성실험에서Em
에대해증식억제대 를형성하지않는균으로구분하였다.
분리균주별Em
내성균 은S. parauberis
가33
균주, E. tarda
는39
균주, Vibrio sp.
로추 정되는균주는9
균주로확인되었다.
내성균주들의제주도내양 식지역별분포를살펴보았을때,
제주시내한림,
애월,
조천,
구 좌등의양식장에서분리된99
균주중내성균주는S. parauberis 5
균주, E. tarda 10
균주, Vibrio sp. 1
균주로총16
균주(16.2%)
가확인되었으며,
성산,
남원,
대정등의서귀포시에서분리된207
균주중내성균주는S. parauberis 28
균주, E. tarda 29
균주, Vibrio sp. 8
균주로총65
균주(31.4%)
가분리되어지역에따른 내성균의출현에차이를보였으며,
내성균주는양식현장이밀 집되어있는서귀포시지역에많이분포하고있음을알수있 었다.
또한,
조사시기에따른내성균의비율을분석하였으며, 2016
년도에분리한163
균주중49
내성균주(30.1%), 2017
년 도에분리한143
균주중32
내성균주(22.4%)
로Em
내성균의비율을확인할수있었다
.
Minimum inhibitory concentration (MIC)
약제감수성검사결과에의하여
Em
에내성을가지는것이확 인된81
내성균들을대상으로MIC test
를실시하여내성수준을 확인하였다. S. parauberis 33
균주중2
균주가100 μg/mL
이상, 16
균주가50 μg/mL, 10
균주는25 μg/mL
그리고5
균주가12.5 μg/mL
이하의MIC
값을나타내었다(Fig. 1). E. tarda
내성균 주에서는5
균주가50 μg/mL,
나머지34
균주가12.5 μg/mL
이 하의값을나타내었다. Vibrio sp. 9
균주는E. tarda
균주와유 사하게7
균주가12.5 μg/mL
이하의값을나타내었으며, 2
균주 가25 μg/mL
의MIC
값을나타내었다.
본실험에서분리한S.
parauberis
균주는그람음성균인E. tarda, Vibrio sp.
에비하여,
Em
에대해넓은범위의MIC
값을가질뿐만아니라비교적높은내성수준의값을가지는것을확인하였다
. 약제감수성
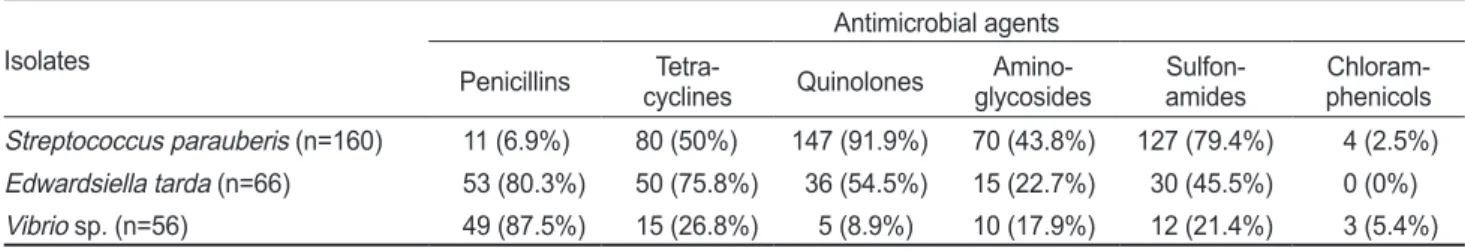
제주지역넙치양식장내병어로부터분리한
S. parauberis, E.
tarda
및Vibrio sp.
균주들에대하여macrolide
계항생제외에 도penicillins, tetracyclines
등을포함한다양한계열의항생 제에대해증식억제대를형성하지않는균주를내성균주로구 분하였으며각균주별내성경향의결과는Table 3
에나타내었 다. S. parauberis
균주는quinolone
계(91.9%), sulfonamide
계(79.4%), tetracycline
계(50%)
항생제순서로높은내성률을보 였으며, E. tarda
균주에서는penicillin
계(80.3%), tetracycline
계(75.8%), quinolone
계(54.5%), sulfonamide
계(45.5%)
항생 제가순서로높은내성비율을나타내어분리된대부분의균주 들이특정계열에국한되지않고다양한계열의항생제에대 해내성을가지는것을확인할수있었다.
그러나, Vibrio sp.
균주에서는
penicillin
계항생제를제외한나머지계열의항생제Fig. 1. MIC value distribution of erythromycin against Streptococcus parauberis, Edwardsiella tarda, Vibrio sp. isolated from olive flounder Paralichthys olivaceus.
0 20 40 60 80 100
more than 100 50 25 less than 12.5
R ate o f r esi sta nt is olates ( %)
MIC value of erythromycin (µg/mL)
Streptococcus parauberis (n=33) Edwardsiella tarda (n=39) Vibrio sp. (n=9)
에대해대체로낮은내성률이관찰되었으며
,
실험에사용된모 든균주는chloramphenicol
계항생제에대해매우낮은내성률 을보였다.
Erythromycin 내성 유전자 분석
Em
에대해내성을가지는내성균으로부터total nucleic acid
를분리한후, PCR
을통해Em
내성유전자를확인하였다. erm (A), erm (B), erm (C), mef (A)
및mef (E)
의검출을위한PCR
은이전Jun (2010)
을참고하여수행하였으며, S. parauberis 33
균주중erm (B)
가24
균주(72.7%)
로확인되었고, 1
균주(3%)
에서erm (A)
가검출되었다(Table 4, Fig. 2). Em
에대해내성 을가지는E. tarda 39
내성균주중5
균주(12.8%)
에서erm (B)
가확인되었으며,
그외다른유전자는발견되지않았다.
또한, Vibrio sp.
균주의경우,
본연구에서검출을시도한5
종류의erm/mef
유전자는검출되지않았다.
고 찰
에드워드병
,
연쇄구균병,
비브리오병,
활주세균병등과같은 세균성 질병은매년 양식현장에서연중 발생하여 피해를유 발하며,
양식현장에서는발생한세균성질병의치료를목적으 로항생제를사용하고있다.
그중Em
의수산용항생제판매 량에근거한사용률은6.3%
로oxytetracycline
에이어국내양 식현장에서많이사용되고있는항생제로알려져있다(APQA,
2009).
이에본연구에서는양식넙치에서분리한어병세균에서Em
내성균주를분리하고, PCR
을통해Em
에대해내성을유발 하는내성유전자를확인하고자하였다.
2016
년부터2017
년까지 제주도 내 넙치 양식장에서S.
parauberis 160
균주, S. iniae 1
균주, E. tarda 66
균주, Vibrio sp.
56
균주및미동정균주23
균주등총306
균주를분리하여실험 에사용하였다.
이전연쇄구균병은L. garvieae, S. parauberis, S. iniae, Enterococcus sp.
등에의해발생된다고보고되었으나(Domeenech et al., 1996; Eldar and Ghittino, 1999),
본연구 에서분리한연쇄구균의세부적인동정결과, S. parauberis
균 주가160
균주로뚜렷하게검출빈도가증가하고있는것을확 인하였다.
이전제주도내연쇄구균병원인체의동정결과, S.
iniae
와S. parauberis
의검출량이비슷하다는보고(Jeong et al., 2006)
와검출률에서차이를보였는데이는이전S. iniae
균주 에대한안정적인백신이개발됨에따라발병률이낮아짐으로 현재S. parauberis
균주에의한연쇄구균병이다발하고있는것 으로보인다.
또한, Thompson et al. (2004)
은비브리오병이60
종이넘는다양한비브리오균에의해발생되며,
계속해서새로 운종이발견되는추세라보고하였으며,
본연구에서는선택배지인
TCBS
배지상에서노란색또는초록색집락을형성하는것으로구분한후
,
이전Demircan and Candan (2006), Kim et
Table 3. Number of bacterial isolates from cultured fish against various antimicrobial agents
Isolates
Antimicrobial agents Penicillins Tetra-
cyclines Quinolones Amino-
glycosides Sulfon-
amides Chloram- phenicols Streptococcus parauberis (n=160) 11 (6.9%) 80 (50%) 147 (91.9%) 70 (43.8%) 127 (79.4%) 4 (2.5%) Edwardsiella tarda (n=66) 53 (80.3%) 50 (75.8%) 36 (54.5%) 15 (22.7%) 30 (45.5%) 0 (0%) Vibrio sp. (n=56) 49 (87.5%) 15 (26.8%) 5 (8.9%) 10 (17.9%) 12 (21.4%) 3 (5.4%)
Table 4. Detection of various erm / mef genes in erythromycin resistant isolates from cultured fish in Jeju
Resistance isolates Genotypes of erythromycin resistance factor
erm (A) erm (B) erm (C) mef (A) mef (E)
Streptococcus parauberis (n=33) 1 24 0 0 0
Edwardsiella tarda (n=39) 0 5 0 0 0
Vibrio sp. (n=2) 0 0 0 0 0
erm, erythromycin ribosome methylation; mef, macrolide efflux.
Fig. 2. DNA amplification of erm gene in erythromycin resistant isolates of Streptococcus parauberis, Edwardsiella tarda and Vibrio sp. from cultured fish in Jeju. Lane 1, S. parauberis-erm (A); Lane 2-9, S. parauberis-erm (B); Lane 10-14, E. tarda- erm (B); Lane 15 and 16, Vibrio sp.-not detected; M, 100bp DNA ladder.
al. (2014), Kim et al. (2015)
의방법을참고하여각균주의세 부적인동정을실시하였다.
그결과, V. alginolyticus
가4
균주, V. harveyi
가2
균주로확인되었다.
이전제주도내Vibrio sp.
균 주들의세부적종을확인한결과, V. scophthalmi, V. harveyi, V.
anguillarum, Photobacterium damselae
등이검출되어본연구 에서동정을실시하지않았던다양한종류의비브리오균주들 이제주도내에분포하고있음을확인할수있었다(Jo, 2006).
넙치로부터분리한어병세균내
Em
내성균주는약제감수성 을통해증식억제대를CLSI (2017)
에서내성의기준으로보는13 mm
이하로규정하였으나내성을보이는모든균주는증식억제대를형성하지않았다
.
그결과S. parauberis 33
균주, E.
tarda 39
균주, Vibrio sp. 9
균주로총81
균주를내성균주로구분 하였다. 2016
년과2017
년도에분리한어병세균내내성균주는 각각47
균주(30.1%), 32
균주(22.4%)
로Em
내성균발생률이 감소한것을확인하였다.
이에조사기간내균종별발생비율을 조사한결과, E. tarda
균주는2016
년과2017
년에각각42.9%, 56.3%
의 내성균이 확인되었으며, Vibrio sp.
균주도10.2%, 12.5%
로E. tarda
균주와유사하게조사기간동안내성균의출 현이증가한것을확인할수있었다.
그러나S. parauberis
균 주는각년도에대해23
균주(46.9%), 10
균주(31.3%)
로 감소 한것을확인하였다.
각균종별내성균발생양상은조사기간 이짧아규정하기에어려움이있으며,
추후Em
내성균주발생 에대한지속적인모니터링이실시되어져야할것으로판단된 다. Em
내성균주들은MIC test
를통해내성수준을살펴본후,
균주들이지니고있는내성인자의유전형을PCR
을통해분석 하였다. CLSI
판단기준(>8 μg/mL)
에따라내성균주를구분하 였을때, Vibrio sp.
균주내확인된Em
내성균주는총2
균주로,
본연구에서검출을시도한어떠한gene
도확인되지않았다.
이 전Luna et al. (1999)
와Portillo et al. (2000)
은장구균의내성 에주로erm (B)
유전자가관여하며,
일부에서erm (A)
가존재 한다고보고하였다.
본연구에서도S. parauberis
에서erm (B)
가24
균주, erm (A)
가1
균주로확인되어이전의보고와유사한 결과를확인할수있었다. E. tarda
를대상으로Em
에대한내성 비율을조사하고유전자분석을실시한이전의연구는없었으 며,
본연구가국내에서의첫보고라고할수있다. E. tarda
의39
균주가Em
에대해내성을보였지만erm (B)
는5
균주에서 만확인할수있었다(Fig. 2). E. tarda
와Vibrio sp.
의내성수준 이S. parauberis
에서보다낮게나타나는것은Em
과같은항생 제가그람음성균보다그람양성균에높은활성을보인다는보 고와연관성이있을것으로여겨진다(Hiroshi, 1996).
또한,
내 성을가지고있지만erm
및mef
유전자를확인할수없는균주 는본연구에서검출을시도하지않은다른유전형이나새로운 종류의내성인자출현등의결과로해석할수있으며, Lee et al.
(2010a)
의연구에서는Em
내성유전자검출을위해erm
과mef
유전자외
msr
유전자를추가적으로실험을실시하였으나검출되지않았으며
, erm (B)
와mef (A)
가높은비율로확인되었다보고하였다
.
이에본연구에서도그람양성균에대해msr
유전자의
primer
를제작하여존재유무를확인할필요가있다고생각된다
.
또한,
비브리오에서는mef
유전자외mph
유전자의확 인이보고되어있으며추후이에대한확인유무도필요할것으 로판단된다(Nonaka et al., 2015).
국내양식현장에서는외국에비해비교적자유롭게항생제 를투여하고있기때문에기존에존재한다고보고되어진유전 자이외의새로운내성결정인자가발생할가능성이높으며
,
이 를방지하기위한지속적인연구가필요하다.
본연구결과는국 내양식넙치에서분리되는어병세균내다양한계열의항생제 내성비율및Em
내성유전자분포에관한기초자료로활용될수 있을것으로사료된다.
사 사
이논문은
2017
학년도제주대학교교원성과지원사업에의하여연구되었음
.
References
APQA (Animal and Plant Quarantine Agency). 2009. National antibiotic use and monitoring of antimicrobial resistance in 2015 [Internet]. Retrieved from www.qia.go.kr on Jun 18, 2018.
Blanco MM, Gibello A, Vela AI, Moreno MA, Dominguez L and Fernandez-Garayzabal JF. 2002. PCR detection and PFGE DNA macrorestriction analyses of clinical isolates of Pseudomonas anguilliseptica from winter disease out- breaks in sea bream Sparus aurata. Dis Aquat Org 50, 19- 27. https://dx.doi.org/10.3354/dao050019.
Cepeda C, Garcia-Marquez S and Santos Y. 2003. Detection of
Flexibacter maritimus in fish tissue using nested PCR am-
plification. J Fish Dis 26, 65-70. https://dx.doi.org/10.1046/j.1365-2761.2003.00431.x.
CLSI (Clinical and Laboratory Standards Institute). 2017. Per- formance standards for antimicrobial susceptibility testing, 28th edition [Internet]. Retrieved from www.clsi.org on Jun 14, 2018.
Demircan D and Candan A. 2006. Identification of Vibrio an-
guilliarum by PCR (rpoN Gene) associated with Vibriosis
in marine fish in Turkey. Turk J Vet Anim Sci 30, 305-310.Domeenech A, Derenaaandez-Garayzabal JF, Pascual C, Garcia JA, Cutuli MT, Moreno MA, Collins MD and Dominguez L. 1996. Streptococcosis in cultured turbot, Scopthalmus
maximus, associated with Streptococcus parauberis. J Fish
Dis 19, 33-38. https://dx.doi.org/10.1111/j.1365-2761.1996.tb00117.x.
Eldar A and Ghittino C. 1999. Lactococcus garvieae and Strep-
tococcus iniae infections in rainbow trout Oncorhynchus
mykiss: similar, but different diseases. Dis Aquat Org 36,
227-231. https://dx.doi.org/10.3354/dao36227.
Han AR, Yoon YJ and Kim JW. 2012. Antibiotic Resistance and Plasmid Profile of Vibrio parahaemolyticus Strains Isolated from Kyunggi-Incheon Coastal Area. Kor J Microbiol 48, 22-28. https://dx.doi.org/10.7845/kjm.2012.48.1.022.
Hansen LH, Mauvais P and Douthwaite S. 1999. The macro- lide-ketolide antibiotic binding site is formed by struc- tures in domains II and V of 23S ribosomal RNA. Mol Microbiol 31, 623-631. https://dx.doi.org/10.1046/j.1365- 2958.1999.01202.x.
Hiroshi N. 1996. Multidrug Efflux Pumps of Gram-Negative Bacteria. J Bacteriol 178, 5853-5859.
Jeong YU, Kang CY, Kim MJ, Heo MS, Oh DC and Kang BJ.
2006. Characterization of Streptococcosis occurrence and molecular identification of the pathogens of cultured floun- der in Jeju island. J Microbiol 42, 199-204.
Jo MR. 2006. Rapid molecular identification and antibiotic re- sistance of Vibrio spp. Isolated from the farmed olive floun- ders in Jeju island, Korea. Master’s Thesis, Jeju National University, Jeju, Korea.
Jun LJ. 2010. Characterization of antibiotic resistant genes car- ried by fish pathogens in Korea. Ph. D. Thesis, Pukyong Na- tional University, Busan, Korea.
Kim HJ, Ryu JO, Lee SY, Kim ES and Kim HY. 2015. Multi- plex PCR for detection of the Vibrio genus and five patho- genic Vibrio species with primer sets designed using com- parative genomics. BMC Microbiol 15, 239. https://dx.doi.
org/10.1186/s12866-015--577-3.
Kim MS, Cho JY and Choi HS. 2014. Identification of Vibrio
harveyi, Vibrio ichthyoenteri and Photobacterium damse- lae isolated from olive flounder Paralichthys olivaceus in
Korea by multiplex PCR developed using the rpoB gene.J Fish Sci 80, 333-339. https://dx.doi.org/10.1007/s12562- 014-0702-5.
Lee SY and Kim YH. 2013. Incidence of Erythromycin Resis- tance Genes, erm(B) and mef(A), in Streptococci Isolated from Dental Plaques of Koreans. Int J Oral Biol 38, 61-65.
https://dx.doi.org/ 10.11620/IJOB.2013.38.2.061.
Lee DW, Jun LJ and Jeong JB. 2016. Distribution of Tetracy- cline Resistance Genes in Pathogenic Bacteria Isolated from Cultured Olive Flounder (Paralichthys olivaceus) in Jeju in 2016. JFMSE 29, 834-846. https://dx.doi.org/10.13000/
JFMSE.2017.29.3.834.
Lee HI, Jung JH, Lee SJ and Choi SS. 2010a. Analysis of Geno- type and Phenotype of Erythromycin Resistance in Entero- cocci spp. Isolated from Raw Milk Samples. Kor J Micro- biol 46, 148-151.
Lee HY, Lim JE, Kim SC, Kim KR, Lee SS, Kwon OK, Yang JE and Ok YS. 2010b. Environmental Monitoring of Selected Veterinary Antibiotics in Soils, Sediments and Water Adja- cent to a Poultry Manure Composting Facility in Gangwon Province, Korea. J Korean Soc Environ Eng 32, 278-286.
Luna VA, Coates P, Eady EA, Cove JH, Nguyen TH and Roberts MC. 1999. A variety of Gram-positive bacteria carry mobile
mef genes. J Antimicro Chemother 44, 19-25. https://dx.doi.
org/10.1093/jac/44.1.19.
Nonaka L, Maruyama F, Suzuki S and Masuda M. 2015. Novel macrolide-resistance genes, mef (C) and mph (G), car- ried by plasmids from Vibrio and Photobacterium isolated from sediment and seawater of a coastal aquaculture site.
Appl Microbiol 61, 1-6. https://onlinelibrary.wiley.com/doi/
abs/10.1111/lam.12414.
Portillo A, Ruiz-Larrea F, Zarazaga M, Alonso A, Martinez JL and Torres C. 2000. Macrolide Resistance Genes in Entero-
coccus spp.. Antimicrob Agents Chemother 44, 967-971.
https://dx.doi.org/10.1128/AAC.44.4.
Sakai T, Iida T, Osatomi K and Kanai K. 2007. Detection of type 1 fimbrial genes in fish pathogenic and non-pathogenic
Edwardsiella tarda strains by PCR. Fish Pathol 42, 115-117.
https://dx.doi.org/ 10.3147/jsfp.42.115.
Sapkota AR, Ojo KK, Roberts MC and Schwab KJ. 2006. An- tibiotic resistance genes in multidrug-resistant Enterococcus spp. And Streptococcus spp. Recovered from the indoor air of a large-scale swine-feeding operation. Lett Appl Micro- biol 43, 534-540. https://dx.doi.org/10.1111/j.1472-765X.
2006.01996.x.
Sung CH, Chon JW, Kwak HS, Kim HS and Seo KH. 2013.
Prevalence and Antimicrobial Resistance of Enterococ-
cus faecalis and Enterococcus faecium Isolated from Beef,
Pork, Chicken and Sashimi. Korean J Food Sci 33, 133-138.https://dx.doi.org/ 10.5851/kosfa.2013.33.1.133.
Thompson FL, Iida T and Swings J. 2004. Biodiversity of Vib- rios. Microbiol Mol Biol Rev 68, 403-431. https://dx.doi.
org/10.1128/MMBR.68.3.
Woo SH, Kim HJ, Lee JS, Kim JW and Park SI. 2006. Patho- genicity and classification of streptococci isolated from cul- tured marine fishes. J Fish Pathol 19, 17-33.