http://dx.doi.org/10.14405/kjvr.2015.55.2.89
89
<원례보저>
Mycobacterium avium subsp. paratuberculosis 감염 초기 개체 검출을 위한 항원 탐색 및 특성 분석
박홍태1·박현의1·신민경1,2·조용일3·유한상1,4,*
1서울대학교 수의과대학, 2Dairy and Swine Research and Development Centre, Agriculture and Agri-Food Canada, Canada,
3농촌진흥청 국립축산과학원, 4서울대학교 그린바이오과학기술연구원 (접수: 2015년 4월 18일, 게재승인: 2015년 5월 11일)
Discovery of antigens for early detection of Mycobacterium avium subsp.
paratuberculosis and analysis of characteristics using bioinformatics tools
Hong-Tae Park
1, Hyun-Eui Park
1, Min-Kyoung Shin
1,2, Yong-Il Cho
3, Han Sang Yoo
1,4,*
1Department of Infectious Diseases, College of Veterinary Medicine, Seoul National University, Seoul 151-742, Korea
2Dairy and Swine Research and Development Centre, Agriculture and Agri-food Canada, Sherbrooke, QC J1M 1Z3, Canada
3Department of Animal Resources Development, National Institute of Animal Science, Rural Development Administration, Cheonan 331-801, Korea
4Institute of Green Bio Science and Technology, Seoul National University, Pyeongchang 232-916, Korea
(Received: April 18, 2015; Accepted: May 11, 2015)
Abstract : Johne’s disease, caused by Mycobacterium avium subsp. paratuberculosis (MAP), is one of the most widespread and economically important diseases in cattle. Current diagnostic methods are based on the detection of anti-MAP antibodies in serum or isolation of the causative agent. However, these techniques are often not applicable for cases of subclinical infection due to relatively low sensitivity. Therefore, finding new antigen candidates that strongly react with the host immune system had been attempted. To effectively detect infection during the subclinical stage, several antigen candidates were selected based on previous researches. Characteristics of the selected antigen candidates were analyzed using bioinformatics-based prediction tools. A total of nine antigens were selected (MAP0862, MAP3817c, MAP2077c, MAP0860c, MAP3954, MAP3155c, MAP1204, MAP1087, and MAP2963c) that have MAP- specific and/or high immune responses to infected animals. Using a transmembrane prediction tool, five of the nine antigen candidates were predicted to be membrane protein (MAP3817c, MAP3954, MAP3155c, MAP1087, and MAP1204). Some of the predicted protein structures identified using the I-TASSER server shared similarities with known proteins found in the Protein Data Bank database (MAP0862, MAP1204, and MAP2077c). In future studies, the characteristics and diagnostic efficiency of the selected antigen candidates will be evaluated.
Keywords : antigen, bioinformatics, discovery, Johne’s disease
서 론
요네병(Johne’s disease)은 Mycobacterium avium subsp.
paratuberculosis(MAP) 균의 감염 때문에 소, 양, 산양 등의 반추류에서 발생하는 만성 소모성 질병이다 [24, 25]. 만성적 인 장염이 특징으로, 장에서의 영양분 흡수 억제로 인한 증 체율 감소, 산유량 감소 등이 나타나는데, 이로 인해 축산농
가에서의 경제적인 피해가 큰 질병 중 하나이다 [8, 25]. 요 네병은 대부분 낙농국가의 젖소 농장에서 감염률이 50%에 이를 정도로 매우 중요한 질병이다 [18, 20]. 또한 MAP는 학술적으로 논란의 여지는 있지만 사람에서의 크론병(Crohn’s disease)과도 연관이 있는 것으로 생각되고 있어 공중보건학 적으로도 중요하게 생각되는 병원체이다 [1, 16, 22].
요네병의 임상 증상은 최초 감염 후 약 2년 또는 그 이상
*Corresponding author
Tel: +82-2-880-1263, Fax: +82-2-874-2738 E-mail: [email protected]
의 잠복기를 거친 후 발현된다 [14]. 임상 증상이 나타나기 전 증상이 없는 상태에서 감염된 개체는 분변을 통해 MAP 를 지속해서 배출하게 되는데, 이를 통해 축사 환경을 오염 시키고 fecal-oral route를 통해 다른 개체의 감염을 유발한 다. 소에서 대부분의 요네병 감염은 이러한 전임상형(pre- clinical)의 형태로 나타나기 때문에 임상 증상을 기초로 한 진단만으로는 요네병을 근절할 수 없다 [6]. 그러므로 전임 상형 상태의 개체를 효율적으로 진단할 방법을 찾는 것이 요 네병의 근절에서 가장 중요하다.
요네병 진단에서 ‘gold-standard’는 분변 시료에서 MAP를 직접 분리 배양하는 것이다 [23]. 원인체를 직접 분리하면 진단의 특이도는 100%이나, MAP가 매우 느리게 자라는 균 이고 분변 시료에서 분리 배양 시 다양한 decontamination 과정을 거치게 되어 결과를 확인하기까지 시간이 매우 오래 걸리며(8~16주), 진단의 민감도도 낮은 편이다 [26, 29]. 최 근 연구 중 배양법을 이용한 진단에서 민감도가 75%인 보 고가 있었으나 시간이 오래 걸리고 상대적으로 비싸다는 단 점이 여전히 존재한다 [15, 24, 29]. 배양법의 이러한 한계 때문에 혈청학적 진단법 중 하나인 enzyme-linked immuno- sorbent assay(ELISA) 법이 요네병의 진단에 널리 사용되고 있다 [18]. ELISA 법이 쉽고 빠르게 결과를 확인할 수 있 다는 점과 더불어 상대적으로 진단 비용이 저렴하다는 장점 이 있기 때문이다 [28]. 그러나 상용화된 ELISA 키트의 진 단 양성률이 무증상의 분변 배양 양성 개체에서 30% 이하 로 나타났다 [10]. 따라서, 무증상으로 균을 배설하는 개체를 효율적으로 찾기 위해서는 감염 초기에 숙주의 면역체계에 감작되는 MAP 특이 항원의 발굴이 필요하다.
본 연구에서는 이미 발표된 논문을 참고하여 요네병 감염 초기에 반응성이 높을 것으로 예상되는 몇 가지 항원 단백 질 후보를 발굴하고 생물정보학 기반 예측프로그램을 이용 하여 그 후보 단백질의 특성을 예상해 보았다. 또한 이를 통 해 실제 ELISA와 같은 진단법에 사용될 가능성을 제시하고 자 하였다.
재료 및 방법
항원 후보 물질 선정
혈청학적 진단법으로 사용될 수 있는 여러 가지 MAP 항 원 단백질을 찾기 위해 기존에 발표된 논문의 연구 결과를 활용하여 몇 가지 효과적일 것으로 예상되는 단백질을 선정 하였다. 감염 초기 반응성이 높은 항원 후보 단백질을 찾기 위해 PubMed(National Center for Biotechnology Information, USA)에서 MAP의 항원 단백질 발굴에 대한 연구 결과를 검 색하였다. 검색된 논문들을 토대로 MAP에 특이적인 단백질 정보 및 여러 가지 test assay에서의 민감도와 특이도에 대한 결과가 밝혀진 단백질을 정리하고, 1) MAP 특이적인 항원 이며, 2) 감염 초기에 반응성이 높고, 3) 항원-항체 반응성이 높은 순으로 우선순위를 정하여 감염 초기 혈청학적 진단에 효율적일 것으로 보이는 항원 후보 단백질을 선정하였다.
항원 후보 단백질의 특성 분석
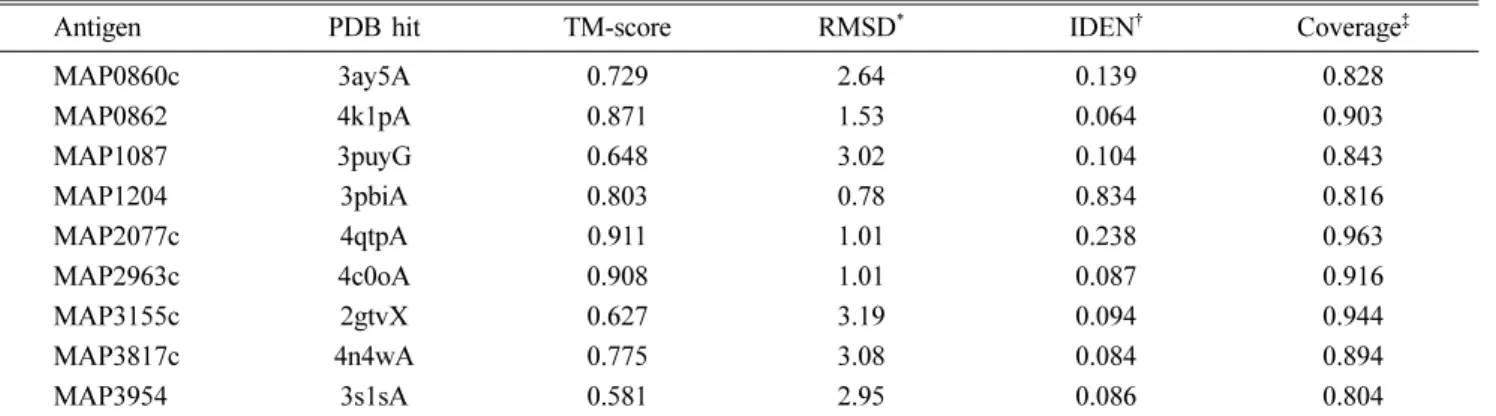
선정된 항원 후보 단백질은 모두 구조와 기능이 밝혀지지 않은 단백질이었기 때문에, 각 항원 후보 단백질의 특성을 파악하기 위해 TMHMM Server ver. 2.0 [13]을 이용하여 단백질에 transmembrane helices가 존재하는지 살펴봄으로써 이들 단백질의 location을 예측하였다. 또한, I-TASSER server [31]를 이용하여 단백질의 3차원 구조를 예측해 봄으 로써 각 단백질의 기능을 추측하였다. 이 프로그램을 이용하 여 이미 구조를 알고 있는 단백질과의 sequence identity 분 석 및 threading aligned region(folding recognition) 분석이 수행되었고, 유사한 folding을 가질 것으로 예상되는 단백질 과의 threading alignment scoring(Normalized Z-score)을 통해 가장 가능성이 큰 3차원 구조가 모델링 되었다. 최종적 으로 5가지의 가능한 모델이 순위(C-score)에 따라 구분되었 고, 그중 가장 순위가 높은 각각의 단백질 예상 구조를 얻었 다. C-score가 높은 몇 가지 단백질은 TM-align tool을 통 해 Protein Data Bank(PDB) database로부터 구조적으로 유 사한 단백질을 TM-score에 따라 10가지로 정리하였다.
결 과
항원 후보 물질 선정
PubMed를 이용하여 검색한 여러 논문을 참고하여 몇 가 지 항원 후보 단백질에 대한 특성을 나열하였다(Table 1).
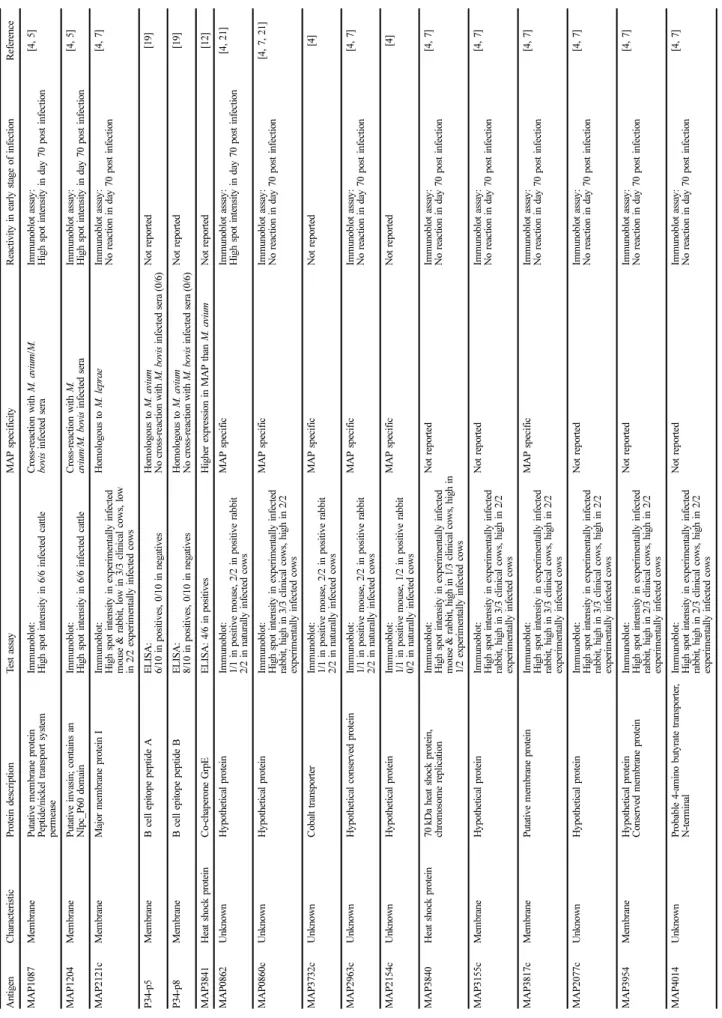
항원성이 높을 것으로 예상되는 분비 단백질과 세포벽 또는 세포막 단백질이 검색되었고, 그중 실험동물 또는 감염된 소 의 혈청과의 반응성이 확인된 단백질에서 field test 결과가 없는 단백질을 정리하였다. 또한 특성이 밝혀지지 않은 단백 질 중에서도 감염 초기 반응성이 높거나 MAP 특이적인 단 백질을 정리하였다. 이들 항원 후보 단백질 중 감염 초기에 반응성이 높은 단백질, MAP에 특이적으로 존재하는 단백질, test assay에서 높은 민감도를 나타낸 단백질의 순으로 우선 순위를 정하여 총 9가지의 항원 후보 물질을 선정하였다 (Table 2).
항원 후보 단백질의 특성 분석
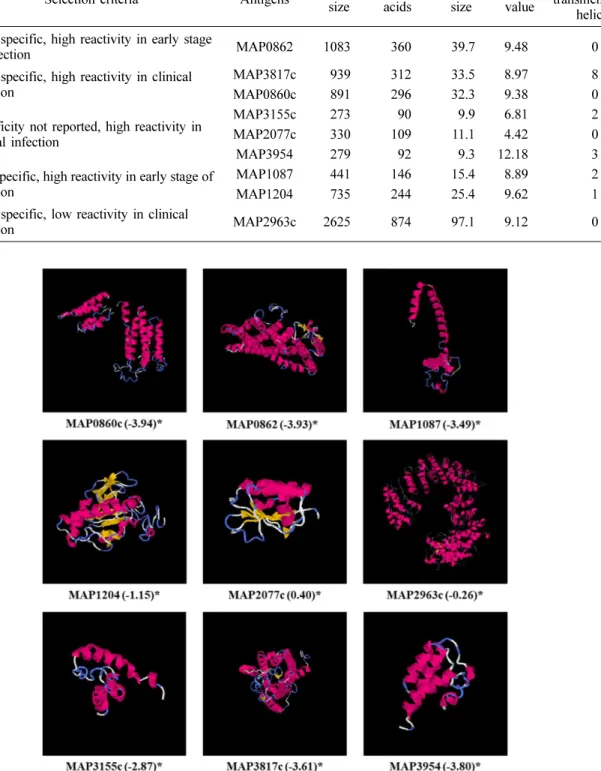
Transmembrane 예측 프로그램에서 9가지 항원 후보 단백 질 중 MAP1087, MAP1204, MAP3155c, MAP3954, MAP3817c 5가지 단백질은 transmembrane helices를 가진 것으로 보여 MAP의 세포벽 또는 세포막에 존재할 것으로 예 측되었다(Table 2). 이 중 MAP1087, MAP1204, MAP3817c 는 이미 putative membrane protein으로 알려진 단백질로 본 연구의 예측 결과와 일치하였다. 나머지 4개의 단백질은 transmembrane helices를 가지지 않는 것으로 예측되었다. I- TASSER server 예측 프로그램을 통한 각 항원 후보 단백질 의 3차원 구조 모델링 결과, 각각에서 가장 가능성이 큰 모 형의 C-score는 −3.94에서 0.40 사이의 값을 나타내었다 (Fig. 1). C-score는 −5에서 2 사이의 숫자로 표시되고 이 숫자가 높을수록 예측 모형의 신뢰도가 높다. MAP2077c의
Ta bl e 1 . Ch ar ac te ris tics o f se ve ra l an ti ge n ca n d id at es e x is tin g in My co ba ct er ium a viu m su bs p. pa ra tube rc ul os is (MA P ) An tigen C haracter istic Pr ot ei n d es cr ipt ion T est ass ay MAP specif icity R eactivity in earl y s tage of inf ec ti on Re fe re n ce MAP 1 0 8 7 M em br an e P u tative m em b ra ne p ro tein Pep tide/nickel tr an spor t sy st em per m ease
Im m uno blot: H igh spo t in tensity in 6/6 in fected ca ttl e Cr oss- re actio n w ith M. a viu m /M. bo vi s in fected sera Im m u n oblot assay : High sp ot intensit y in d ay 70 p o st inf ec ti on [4 , 5 ] MAP 1 2 0 4 M em br an e P u tative invasin; cont ai ns a n N lpc _P 60 do m ai n Im m uno blot: H igh spo t in tensity in 6/6 in fected ca ttl e Cr oss- re actio n w ith M. aviu m/M . b ovis inf ec ted ser a Im m u n oblot assay : High sp ot intensit y in d ay 70 p o st inf ec ti on [4 , 5 ] MAP 2 1 21c Mem b ra n e Ma jo r m em b ra n e pr otein I Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d m o u se & rabb it, low in 3/ 3 clin ica l cow s, low in 2 /2 ex perim entally in fected co ws
H o m olog ous to M. lepr ae Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ] P 34- p5 Mem b ra n e B cell e p itope peptide A E LI SA : 6 /10 in posi tives, 0/1 0 in n eg atives H o m olog ous to M. aviu m N o cr oss- re action wi th M. bo vi s i n fected sera ( 0/6 ) Not r ep o rt ed [1 9] P 34- p8 Mem b ra n e B cell e p itope peptide B E LI SA : 8 /10 in posi tives, 0/1 0 in n eg atives H o m olog ous to M. aviu m N o cr oss- re action wi th M. bo vi s i n fected sera ( 0/6 ) Not r ep o rt ed [1 9] MAP 3 8 4 1 H eat shock pr otein Co- ch ap er o n e Gr pE E L IS A : 4/6 in posi tives H igher exp ressi on in M A P th an M . avium N o t re po rt ed [1 2] MAP 0 8 6 2 U n know n H y p othetical pr otein Im m uno blot: 1 /1 in p o siti ve m o u se, 2 /2 in posit ive r abbit 2 /2 in n atur al ly i n fe cted cows MAP specif ic Im m u n oblot assay : High sp ot intensit y in d ay 70 p o st inf ec ti on [4 , 21] MAP 0 8 60c Un know n H y p othetical pr otein Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d ra bbit , high in 3/3 clini ca l cows, h igh in 2/2 exp er im entally in fected co ws
MAP specif ic Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7, 21 ] MAP 3 7 32c Un know n C obalt tr an spor ter Im m uno blot: 1 /1 in p o siti ve m o u se, 2 /2 in posit ive r abbit 2 /2 in n atur al ly i n fe cted cows M A P s p ec ifi c N o t re po rt ed [4 ] MAP 2 9 63c Un know n H y p othetical c o nser ved p ro tein Im m uno blot: 1 /1 in p o siti ve m o u se, 2 /2 in posit ive r abbit 2 /2 in n atur al ly i n fe cted cows
MAP specif ic Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ] MAP 2 1 54c Un know n H y p othetical pr otein Im m uno blot: 1 /1 in p o siti ve m o u se, 1 /2 in posit ive r abbit 0 /2 in n atur al ly i n fe cted cows M A P s p ec ifi c N o t re po rt ed [4 ] MAP 3 8 4 0 H eat shock pr otein 70 kDa hea t sh ock pro tein, chr o m o som e repli ca tio n Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d m o u se & rabb it, hig h in 1 /3 clinical cows , hig h in 1 /2 e x perim ent al ly in fected cows
N ot r epor ted Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ] MAP 3 1 55c Mem b ra n e Hy p othetical pr otein Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d ra bbit , high in 3/3 clini ca l cows, h igh in 2/2 exp er im entally in fected co ws N ot r epor ted Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ] MAP 3 8 17c Mem b ra n e Pu tative m em b ra ne p ro tein Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d ra bbit , high in 3/3 clini ca l cows, h igh in 2/2 exp er im entally in fected co ws
MAP specif ic Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ] MAP 2 0 77c Un know n H y p othetical pr otein Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d ra bbit , high in 3/3 clini ca l cows, h igh in 2/2 exp er im entally in fected co ws N ot r epor ted Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ] MAP 3 9 5 4 M em br an e H y p othetical pr otein Conser ved m em b ra ne pr otein Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d ra bbit , high in 2/3 clini ca l cows, h igh in 2/2 exp er im entally in fected co ws
N ot r epor ted Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ] MAP 4 0 1 4 U n know n P ro b able 4- am ino bu ty ra te tr anspo rter , N- term in al Im m uno blot: H igh spo t in tensity in exper im entally inf ecte d ra bbit , high in 2/3 clini ca l cows, h igh in 2/2 exp er im entally in fected co ws
N ot r epor ted Im m u n oblot assay : No reaction in day 70 po st i n fe ctio n [4 , 7 ]
경우 TM-align을 통한 유사 단백질 분석에서 가장 유사한 구조를 가질 것으로 예상되는 단백질은 MAP의 MAP0380 유전자가 coding하고 있는 anti-sigma factor antagonist protein(PDB ID: 4qtp, TM-score: 0.911)으로 분석되었고, 두 번째로 유사한 단백질은 Bacillus의 anti-sigma F factor protein(PDB ID: 1th8, TM-score: 0.852)였다. 나머지 유사
성이 높은 단백질은 모두 anti-sigma factor antagonist의 기 능을 가지는 단백질이었다. MAP1204의 경우 가장 비슷한 구조로 예상되는 단백질은 Mycobacterium(M.) tuberculosis의 RipB protein (PDB ID: 3 pbi, TM-score: 0.803)으로 분석 되었고 이 단백질과 MAP1204의 sequence identity는 83.4%로 나타났다. 또한 3순위로 유사한 구조로 예상되는 단 Table 2. List of selected antigen candidates and the selection criteria
Rank Selection criteria Antigens Gene
size
Amino acids
Protein size
pI value
Predicted transmembrane
helices 1 MAP specific, high reactivity in early stage
of infection MAP0862 1083 360 39.7 9.48 0
2 MAP specific, high reactivity in clinical infection
MAP3817c 939 312 33.5 8.97 8
MAP0860c 891 296 32.3 9.38 0
3 Specificity not reported, high reactivity in clinical infection
MAP3155c 273 90 9.9 6.81 2
MAP2077c 330 109 11.1 4.42 0
MAP3954 279 92 9.3 12.18 3
4 Non-specific, high reactivity in early stage of infection
MAP1087 441 146 15.4 8.89 2
MAP1204 735 244 25.4 9.62 1
5 MAP specific, low reactivity in clinical
infection MAP2963c 2625 874 97.1 9.12 0
Fig. 1. Three-dimensional structure prediction using I-TASSER server. *C-score.
백질은 MAP1204의 p60 domain으로 나타났는데(PDB ID:
3i86, TM-score: 0.547), sequence identity가 100%인데 반 해 TM-score는 상대적으로 낮았다. 그 외 단백질들의 TM- align 결과가 Table 3에 정리되었다.
고 찰
요네병에 감염된 개체는 대부분이 증상이 없는 상태에서 분변으로 균을 배설하기 때문에 이러한 무증상 감염우를 찾 는 것이 요네병의 제어에 있어서 가장 중요한 요소이다. 혈 청학적 진단 방법인 ELISA 법은 진단의 신속성과 편의성, 비용적 측면에서 장점을 가지고 또한 많은 개체를 스크리닝 하는 데 적합한 진단법이다. 무증상 감염우를 ELISA 법으로 효율적으로 진단하기 위해서는 MAP 감염 초기에 숙주동물 의 면역체계와 반응하는 항원을 찾는 것이 중요하다. 혈청학 적 진단에 사용할 수 있는 항원은 민감도와 특이도의 두 가 지 측면에서 분석할 수 있다. 첫째로, 민감도에서 항원 후보 물질은 항원성(antigenicity)을 가지고 있고 MAP에 감염된 동물의 면역체계에 감작될 수 있어야 한다. 분비 단백질이나 세포막 또는 세포벽을 구성하고 있는 단백질이 숙주동물의 면역체계에 감작될 확률이 높다 [9]. 둘째로, 특이도에서 항 원 후보물질은 MAP에 특이적으로 존재하는 단백질이어야 한다. MAP의 whole genome sequencing 정보를 이용하여 다른 Mycobacterium 속균의 genome과 비교 분석함으로써 MAP만이 특이적으로 가지고 있는 open reading frame(ORF) site를 찾을 수 있다 [2, 21].
본 연구에서는 이미 발표된 논문을 참고하여 민감도와 특 이도가 높은 항원 후보물질을 선발하고, 몇 가지 생물정보학 기반 예측프로그램을 이용하여 선발한 항원 후보물질들의 특 성을 분석함으로써 실제 진단에서의 사용 가능성을 예측하 였다. 선행 연구 결과 분석을 통해 총 9가지의 항원 후보 단백질이 선정되었다.
Mikkelsen 등 [17]이 정리한 목록에 의하면, MAP 진단에 사용 가능할 것으로 생각되는 항원 단백질 중 분비 단백질
이 28종, 세포막 또는 세포벽 단백질이 18종 정리되었다. 또 한 Heat shock 단백질로 5종, 기능이 알려지지 않은 단백질 42종이 정리되었다. 정리된 항원 단백질 중, 세포막 또는 세 포벽의 구성 단백질인 MAP1087, MAP1204, MAP2121c는 다른 Mycobacterium 속균과 유사성을 가지지만 실험적으로 감염한 개체에서 높은 양성율을 보였다 [4, 5, 7]. 또 다른 세포막 또는 세포벽 구성 단백질인 P34 단백질의 epitope 분석을 통해 만들어진 P34-p5, P34-p8의 두 재조합 단백질 은 ELISA test에서 높은 양성율을 나타내었고 M. bovis 감 염 혈청과의 교차반응이 관찰되지 않았다 [19]. Heat shock 단백질로 가능성이 큰 것은 MAP3841로 M. avium subsp.
avium 보다 MAP에서 더 높은 수준으로 발현되고 ELISA test에서도 높은 민감도를 나타내었다 [12]. 기능이 알려지지 않 은 단백질 중 항원으로 가능성이 큰 것은 MAP에 특이적으로 존재하는 MAP0860c, MAP0862, MAP2154c, MAP2963c, MAP3732c 등이었다 [21]. Paustian 등 [21]은 2004년에 발 표한 논문에서 MAP의 whole genome sequencing 분석을 통해 13개의 MAP 특이 ORF site를 확인하였다. 이 중 5 개의 ORF에 대한 재조합 단백질을 제조하여 실험동물 및 요네병 임상 증상을 나타내는 동물의 혈청과 immunoblot 분 석을 시행하였는데 모든 혈청과 반응한 것은 MAP0862, MAP3732c, MAP2963c(C-terminal half)였으며 실험동물 혈 청에만 반응한 것은 MAP2154c, 모두와 반응하지 않은 것은 MAP0860c였다 [21]. Bannantine 등 [7]이 2008년에 실시 한 protein array 분석에서 여러 가지 MAP 항원단백질을 분석하였는데 MAP 접종 마우스와 토끼 혈청에 높은 반응성 을 나타낸 단백질은 MAP3840, MAP2121c였고 임상 증상을 나타내는 소 혈청에 높은 반응성을 나타낸 것은 MAP3155c, MAP3817c, MAP2077c, MAP3954, MAP4014였으며, 실험 감염한 소 혈청에 높은 반응성을 나타낸 것은 MAP3155c, MAP2077c, MAP0860c, MAP3817c, MAP3954였다. Ban- nantine 등 [4]이 2008년에 실시한 또 다른 연구에서는 실험 적으로 감염한 소에서 감염 초기 혈청에 가장 강한 반응성 을 나타낸 항원은 MAP0862, MAP1087, MAP1204였다.
Table 3. TM-align analysis of antigen candidates
Antigen PDB hit TM-score RMSD* IDEN† Coverage‡
MAP0860c 3ay5A 0.729 2.64 0.139 0.828
MAP0862 4k1pA 0.871 1.53 0.064 0.903
MAP1087 3puyG 0.648 3.02 0.104 0.843
MAP1204 3pbiA 0.803 0.78 0.834 0.816
MAP2077c 4qtpA 0.911 1.01 0.238 0.963
MAP2963c 4c0oA 0.908 1.01 0.087 0.916
MAP3155c 2gtvX 0.627 3.19 0.094 0.944
MAP3817c 4n4wA 0.775 3.08 0.084 0.894
MAP3954 3s1sA 0.581 2.95 0.086 0.804
*The RMSD between residues that are structurally aligned by TM-align. †The percentage sequence identity in the structurally aligned region.
‡It represents the coverage of the alignment by TM-align and is equal to the number of structurally aligned residues divided by length of the query protein.
MAP0862는 MAP에 특이적으로 존재하면서 인공 감염한 소에서 감염 초기에 높은 반응성을 보였는데, 이 단백질의 3 차원 구조 예측 결과 C-score는 상대적으로 낮았지만 TM- align 결과는 Bacillus cereus의 NheA 단백질과 유사성이 예 측되었다 [11]. NheA 단백질은 pore forming toxin의 family 로 알려졌으며 향후 연구를 통해서 MAP0862의 구조를 밝 혀내면 실제 이 단백질의 병원성 인자로서의 가능성을 분석 할 수 있다. 인공 감염한 소에서 감염 초기 높은 반응성을 보인 또 다른 단백질인 MAP1087, MAP1204의 경우 M.
bovis와 M. avium에 감염된 혈청과 교차반응이 보고되었다 [5]. 특히, MAP1204의 경우 M. tuberculosis의 RipB 단백 질과 구조적으로 유사하였고 [3], T-BLASTn 분석 결과 M.
tuberculosis 및 M. bovis에서 84%의 아미노산이 동일하였으 며 M. avium complex에서는 아미노산이 100% 동일하였다.
MAP1087은 M. avium subsp. avium 104 strain과 99%, M. bovis의 부분 서열과 29% 동일함이 보고되었다 [5].
MAP3817c, MAP3155c, MAP3954는 membrane protein으 로 숙주세포의 면역체계에 감작될 확률이 높을 것으로 생각 된다. MAP3155c의 경우 T-BLASTn 검색에서 M. avium subsp. avium 104 strain과 98% 동일하였으며, M. smegmatis 와는 46% 정도 동일함을 보였다(data not shown). MAP3954 는 M. avium subsp. avium 104 strain과 96%, M. bovis 와는 71% 동일하였다(data not shown). 이 두 단백질은 실 험적으로 M. bovis 감염 혈청 또는 M. avium subsp.
avium 감염 혈청과의 반응성이 확인되어야 할 것이다.
MAP2077c의 경우 anti-sigma factor antagonist protein과의 유사성이 예측되어 세포질 내 단백질일 것으로 생각되며, stress와 같은 외부의 자극에 따라 발현이 조절되는 단백질일 것으로 추측한다. MAP0860c의 경우 MAP 특이적인 단백질 이며 임상 증상을 나타내는 소에서의 반응성도 높은 것으로 보고되었는데 [7], I-TASSER server를 이용한 3차원 구조 모형에서 유사한 구조를 가지는 세균 단백질은 찾을 수 없 었다. 이에 따라 MAP0860c 단백질은 기존에 구조와 기능이 알려진 세균 유래 단백질에는 존재하지 않는 새로운 기능을 가진 단백질일 것으로 추측된다.
선정된 항원 후보 단백질은 실험동물이나 인공 감염한 소 또는 제한된 숫자의 임상 증상을 나타내는 소 혈청과의 항 원-항체 반응성이 증명되었기 때문에 실제로 자연 감염이 일 어난 소의 혈청을 이용한 반응성이 증명되어야 할 필요가 있 다. 향후 연구에서, 이들 항원 후보 물질의 재조합 단백질을 생산한 후 MAP에 자연 감염된 소 혈청에 대한 반응성을 확인하고 antigen cocktail을 조합한다면, 이를 바탕으로 ELISA 기법을 확립할 수 있을 것으로 기대된다.
감사의 글
본 연구는 농촌진흥청 연구사업(PJ00897001), BK21플러스 수의창의연구 인력양성 사업단 및 서울대학교 수의과학연구 소의 지원으로 이루어졌습니다.
References
1. Abubakar I, Myhill D, Aliyu SH, Hunter PR. Detection of Mycobacterium avium subspecies paratuberculosis from patients with Crohn’s disease using nucleic acid-based techniques: a systematic review and meta-analysis. Inflamm Bowel Dis 2008, 14, 401-410.
2. Alexander DC, Turenne CY, Behr MA. Insertion and deletion events that define the pathogen Mycobacterium avium subsp. paratuberculosis. J Bacteriol 2009, 191, 1018-1025.
3. Böth D, Schneider G, Schnell R. Peptidoglycan remodeling in Mycobacterium tuberculosis: comparison of structures and catalytic activities of RipA and RipB. J Mol Biol 2011, 413, 247-260.
4. Bannantine JP, Bayles DO, Waters WR, Palmer MV, Stabel JR, Paustian ML. Early antibody response against Mycobacterium avium subspecies paratuberculosis antigens in subclinical cattle. Proteome Sci 2008, 6, 5.
5. Bannantine JP, Paustian ML, Waters WR, Stabel JR, Palmer MV, Li L, Kapur V. Profiling bovine antibody responses to Mycobacterium avium subsp. paratuberculosis infection by using protein arrays. Infect Immun 2008, 76, 739- 749.
6. Bannantine JP, Rosu V, Zanetti S, Rocca S, Ahmed N, Sechi LA. Antigenic profiles of recombinant proteins from Mycobacterium avium subsp. paratuberculosis in sheep with Johne’s disease. Vet Immunol Immunopathol 2008, 122, 116- 125.
7. Bannantine JP, Waters WR, Stabel JR, Palmer MV, Li L, Kapur V, Paustian ML. Development and use of a partial Mycobacterium avium subspecies paratuberculosis protein array. Proteomics 2008, 8, 463-474.
8. Chacon O, Bermudez LE, Barletta RG. Johne’s disease, inflammatory bowel disease, and Mycobacterium paratu- berculosis. Annu Rev Microbiol 2004, 58, 329-363.
9. Cho D, Collins MT. Comparison of the proteosomes and antigenicities of secreted and cellular proteins produced by Mycobacterium paratuberculosis. Clin Vaccine Immunol 2006, 13, 1155-1161.
10. Collins MT, Wells SJ, Petrini KR, Collins JE, Schultz RD, Whitlock RH. Evaluation of five antibody detection tests for diagnosis of bovine paratuberculosis. Clin Diagn Lab Immunol 2005, 12, 685-692.
11. Ganash M, Phung D, Sedelnikova SE, Lindbäck T, Granum PE, Artymiuk PJ. Structure of the NheA component of the Nhe toxin from Bacillus cereus: implications for function. PLoS One 2013, 8, e74748.
12. Hughes V, Bannantine JP, Denham S, Smith S, Garcia- Sanchez A, Sales J, Paustian ML, Mclean K, Stevenson K.
Immunogenicity of proteome-determined Mycobacterium avium subsp. paratuberculosis-specific proteins in sheep with paratuberculosis. Clin Vaccine Immunol 2008, 15, 1824-1833.
13. Krogh A, Larsson B, von Heijne G, Sonnhammer ELL.
Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J Mol Biol 2001, 305, 567-580.
14. Irenge LM, Walravens K, Govaerts M, Godfroid J, Rosseels V, Huygen K, Gala JL. Development and validation of a triplex real-time PCR for rapid detection and specific identification of M. avium subsp. paratuberculosis in faecal
samples. Vet Microbiol 2009, 136, 166-172.
15. Lambrecht RS, Carriere JF, Collins MT. A model for analyzing growth kinetics of a slowly growing Mycobacterium sp. Appl Environ Microbiol 1988, 54, 910-916.
16. Mendoza JL, Lana R, Díaz-Rubio M. Mycobacterium avium subspecies paratuberculosis and its relationship with Crohn’s disease. World J Gastroenterol 2009, 15, 417-422.
17. Mikkelsen H, Aagaard C, Nielsen SS, Jungersen G. Review of Mycobacterium avium subsp. paratuberculosis antigen candidates with diagnostic potential. Vet Microbiol 2011, 152, 1-20.
18. Nielsen SS, Toft N. A review of prevalences of paratuberculosis in farmed animals in Europe. Prev Vet Med 2009, 88, 1-14.
19. Ostrowski M, Mundo SL, Harris NB, Barletta RG, Lopez OJ. B-cell epitopes in the immunodominant p34 antigen of Mycobacterium avium ssp. paratuberculosis recognized by antibodies from infected cattle. Scand J Immunol 2003, 58, 511-521.
20. Ott SL, Wells SJ, Wagner BA. Herd-level economic losses associated with Johne’s disease on US dairy operations. Prev Vet Med 1999, 40, 179-192.
21. Paustian ML, Amonsin A, Kapur V, Bannantine JP.
Characterization of novel coding sequences specific to Mycobacterium avium subsp. paratuberculosis: implications for diagnosis of Johne’s disease. J Clin Microbiol 2004, 42, 2675-2681.
22. Pierce ES. Where are all the Mycobacterium avium subspecies paratuberculosis in patients with Crohn’s disease?
PLoS Pathog 2009, 5, e1000234.
23. Sockett DC, Conrad TA, Thomas CB, Collins MT.
Evaluation of four serological tests for bovine paratuberculosis. J Clin Microbiol 1992, 30, 1134-1139.
24. Stabel JR. Johne’s disease: a hidden threat. J Dairy Sci 1998, 81, 283-288.
25. Sweeney RW. Pathogenesis of paratuberculosis. Vet Clin North Am Food Anim Pract 2011, 27, 537-546.
26. Sweeney RW. Transmission of paratuberculosis. Vet Clin North Am Food Anim Pract 1996, 12, 305-312.
27. Weber MF, Verhoeff J, van Schaik G, van Maanen C.
Evaluation of Ziehl-Neelsen stained faecal smear and ELISA as tools for surveillance of clinical paratuberculosis in cattle in the Netherlands. Prev Vet Med 2009, 92, 256-266.
28. Weldingh K, Rosenkrands I, Okkels LM, Doherty TM, Andersen P. Assessing the serodiagnostic potential of 35 Mycobacterium tuberculosis proteins and identification of four novel serological antigens. J Clin Microbiol 2005, 43, 57-65.
29. Wells SJ, Collins MT, Faaberg KS, Wees C, Tavornpanich S, Petrini KR, Collins JE, Cernicchiaro N, Whitlock RH.
Evaluation of a rapid fecal PCR test for detection of Mycobacterium avium subsp. paratuberculosis in dairy cattle.
Clin Vaccine Immunol 2006, 13, 1125-1130.
30. Whitlock RH, Wells SJ, Sweeney RW, Van Tiem J. ELISA and fecal culture for paratuberculosis (Johne’s disease):
sensitivity and specificity of each method. Vet Microbiol 2000, 77, 387-398.
31. Zhang Y. I-TASSER server for protein 3D structure prediction. BMC Bioinformatics 2008, 9, 40.