서 론
일반적으로 어류의 소화관은 성장에 필요한 영양분의 소화 흡수를 담당하는 중요한 기관으로서 초기 발육단계인 자어에 서 치어기로 이행되는 시기에 형태적, 생리적 변화 중 내부영 양에서 외부영양으로 영양원의 치환 및 섭취와 관련된 소화 기관의 발달은 자치어의 생존과 성장에 크게 영향을 미친다 (Han et al., 2007).
성장에 따른 소화관 분화와 발달을 규명하는 것은 각 성장
시기별로 어류의 소화기능에 관한 정보를 얻을 수 있고, 종묘 생산과정에서 초기 대량 폐사를 줄여 높은 생존율을 얻는 데 중요한 기초자료가 된다(Kim, 2000).
어류의 소화관 발달에 관한 국내 연구로는 감성돔의 소화관 구조 및 조직학적 특징(Lee and Chin, 1999), 자주복의 소화관 발달(Kim et al., 2003), 넙치 자치어의 소화기관 형태발달(Han et al., 2007), 고등어 자치어의 소화기관 발달 및 조직학적 연 구(Park, 2013) 등이 수행되었고, 국외 연구로는 Paralabrax maculatofasciatus 자어의 소화관 발달(Pena et al., 2003), Pagel
lus erythrinus 자어의 소화관 발달(Micale et al., 2006), Miich
thys miiuy 자어의 소화기관 발달(Lian et al., 2007), 흰동가리
—9 — http://www.fishkorea.or.kr
능성어 (Epinephelus septemfasciatus) 자어의 소화기관 구조 및 조직학적 특징
박종연·김나리1·박재민2·명정인·조재권
*
국립수산과학원 양식관리과, 1울주군 축수산과, 2경상북도 토속어류산업화센터
The Structure of Digestive Tract and Histological Features of the Larvae in Sevenband Grouper, Epinephe
lus septemfasciatus by Jong Youn Park, Na Ri Kim1, Jae Min Park2, Jeong In Myeong and Jae Kwon Cho* (Aquacul
ture Management Division, Aquaculture Research Institute, NIFS, Busan 46083, Korea, 1Uljugun Metropolitan City Livestock and Marine Industry Division, Ulju 44663, Korea, 2GyeongsangbukDo Native Fish Business Center, Uiseong 37366, Korea)
ABSTRACT
Histological and morphological development of the digestive tract of sevenband grouper were observed from after hatching to 60 days. Fishes were fed with rotifer (Brachionus rotundiformis) and chlorella (Chlorella ellipsoidea) after hatching from 2 to 20 days rotifer and brine shrimp (Artemia
salina) in after 20 days rotifer, brine shrimp and semi-dry artificial diet in after 23 days. Histological andmorphological development of ten larvae was observed by paraffin embedding method after fixing in 10% neutral buffered formalin. Sevenband grouper RLG showed characteristics of carnivorous fish by average 0.87. Larvae after hatching can’t open the mouth and anus digestive tract was observed in a straight line following yolk sac. Larvae was observed feeding activity by opening the mouth and anus. Metamorphosis started 8 days after hatching. Esophagus divided four layer, and goblet cell was observed in esophagus, mid intestine and rectum. Larvae started cannibalism and it was caused by difference of growth. The inside of stomach was differentiated to cardiac orifice, body of stomach, pyloric stomach, and pyloric caeca. Goblet cell was observed all intestine. Gastric gland differentiated after hatching 28 days in stomach. Secretion of gastric juice was found at stomach and mucosal fold pyloric caeca. Even thought the inside of stomach expended and the number of gastric gland increased consistently and goblet cell in intestine and mucosa became longer, histochemical changes follow couldn’t be found during transforming juveniles 38 days after hatching.
Key words: Digestive, Epinephelus septemfasciatus, juvenile, larvae, sevenband grouper
* Corresponding author: Jae Kwon Cho Tel: 82517202420, Fax: 82517202419, Email: [email protected] ISSN: 12258598(Print), 22883371(online)
Accepted: March 25, 2016
의 소화기관 발달(Umur and Chris, 2008), Ompok bimaculatus 자어의 소화기관 발달(Pradhan et al., 2012), 흉기흑점바리 자 어의 소화관 점액세포(Li et al., 2012) 등이 수행되었다.
능성어류는 분류학적으로 농어목(Perciformes) 바리과(Ser
ranidae)에 속하는 어류로서 전 세계에 62속 450여 종으로 알 려져 있으며, 우리나라에는 능성어 Epinephelus septemfasciatus 를 비롯한 붉바리 E. akaara, 자바리 E. bruneus 등 12속 27종 이 서식하고 있다(Kim et al., 2005).
능성어는 우리나라 남해안을 비롯한 제주도 연안 그리고 아 열대와 열대지방의 암초 및 산호초가 발달한 지역에 주로 서 식하고 있으며(Lee and Go, 2003), 능성어에 대한 연구로는 성전환 유도(Lee et al., 1996), 난소 발달과 난모세포의 성숙 (Shein et al., 2004), 성숙과 배란유도(Song et al., 2008), 신경 괴사 바이러스와 예방접종(Oh et al., 2012) 등 많은 연구가 수 행되었으나 바리과 어류의 소화관에 대한 연구는 부족한 실정 이다.
최근 능성어의 종묘생산 연구가 활발히 진행되면서 생산과 정 중에 일어나는 문제점으로 어미관리, 기형발생 및 먹이생 물 등이 초기 폐사 원인으로 대두되고 있다(Park et al., 2015).
따라서 본 연구에서는 능성어의 자어 시기에 나타나는 소화 기관 구조 및 조직학적 특징을 관찰하여 능성어의 소화기능에 관한 정보와 종묘 생산과정에 있어 적절한 먹이 투여시기 및 높은 생존율을 얻기 위한 기초자료로 이용하고자 한다.
재료 및 방법
1. 친어관리 및 배란유도
친어는 2012년 6월 전라남도 여수시 삼산면 거문리 소재의 해상가두리(7×7m)에서 8년간 양성된 성숙한 암컷과 수컷 5 마리(전장 67.5~83.5cm, 평균 74.8±5.3cm)를 선별하여 배 란유도에 사용하였다. 성숙이 확인된 친어는 성선자극호르몬 HCG(human chorionic gonadotropin)를 어체 중 1kg당 500IU 주사하였으며, 복부를 압박하여 채란 및 채정 후 인공수정 시 켰다.
2. 자어 사육관리
자어는 폴리프로필렌(PP, Polypropylene) 재질의 원형 수조 (100×50×80cm)에서 지수식으로 사육하였고, 수온은 21.0~
23.0℃(평균 22.0±1.0℃), 염분농도는 32.0~33.2‰(평균 33.0‰) 범위를 유지하였다. 먹이공급은 부화 후 2일부터 20일 까지 S타입의 로티퍼 Brachionus rotundiformis와 해산클로렐 라 Chlorella ellipsoidea를 혼합 공급하였고, 20일부터 로티퍼+
알테미아 Artemia salina를 혼합 공급하였으며, 23일부터 로티
퍼+알테미아+배합사료를 혼합 공급하였다. 먹이 종류별 급 이 밀도는 해산클로렐라 mL당 500~750만/cell, 로티퍼 19~
20개체, 알테미아는 10~11개체를 유지시켰다.
3. 소화관 외부형태 및 조직학적 관찰
소화관의 외부형태 관찰을 위해 자어를 부화 직후부터 20일 까지 1일 1회, 치어로 이행하는 시기까지 2일 1회 채집하였고, 10마리를 임의로 선택하여 10% 중성 포르말린에 고정 후 만 능투영기(Nikon JP V12B, Japan)를 이용, 전장을 0.01mm까 지 측정하였으며, 입체해부현미경(NIKON NM40, Japan) 하 에서 디지털카메라(Nikon Coolpix 995, Japan)로 관찰 및 사진 촬영을 하였다. 치어기에 달한 개체는 복부를 절개하여 10%
중성 포르말린에 고정하였고, 식도에서 항문 말단까지의 소화 관 길이를 0.1cm까지 측정하였다. 자치어의 형태 발달 단계는 Kendall et al.(1984)에 따라 구분하였고, 자치어의 외부 형태 는 Han and Kim(1997)에 따랐다.
조직학적 관찰을 위해 자어 6마리를 임의로 선택하여 10%
중성 포르말린에 고정 후 Bouin’s solution(Picric acid:forma
lin:Acetic acid=15:5:1)에 일정시간 재고정하였으며, 전장 10.0mm 이상은 두부와 미부를 제거한 뒤 실험에 사용하였다.
고정된 조직은 파라핀 포매법에 의한 블럭을 제작한 뒤 파라 핀 절편법에 의해 4~5μm로 연속 절편하였다. 이때 절편방향 은 Cross와 Sagittal을 실시하였으며, 각 Mayer’s haematoxylin
eosin(HE) 대비염색과 Periodic acidSchiff(ABPAS, pH 2.5) 반응을 실시하여 광학현미경 관찰용 조직표본을 제작하였다. 조직표본은 image measurement system(FOCUS technology, 2005)이 부착된 광학현미경(Olympus, CX31, Japan)으로 관찰 하였다.
A
B
Pc Pa
St
Sp Ai
Pi
Mi
An
Li ep
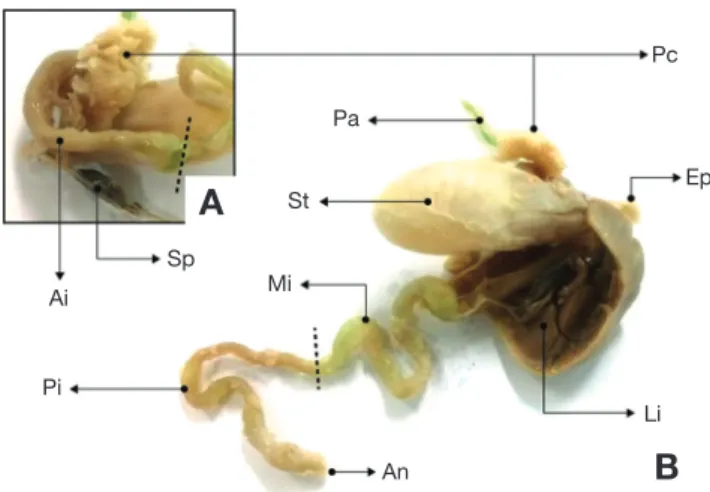
Fig. 1. Morphological observations of the alimentary tract in seven
band grouper, Epinephelus septemfasciatus. A: Ai, Anterior intestine;
Pc, Pyloric stomach; Sp, Spleen; B: An, Anus; Ep, Esophagus; Li, Liver; Mi, Mid intestine; Pi, Posterior intestine; St, Stomach.
결 과
1. 소화관의 외부형태
능성어 소화관의 외부형태는 식도, 위, 장 및 항문이 일련의
관을 이루었고, 간은 다른 소화기관에 비하여 컸으며, 다른 장 기를 둘러싸고 있는 형태로 관찰되었다. 아래쪽에는 비장과 이자가 위치하였고, 위의 구조는 분문위, 위체부 및 유문부로 나누어지며, 유문부에는 30~32개 내외의 유문수가 많은 분지 를 가진 맹낭을 형성하였다(Fig. 1A). 소화관의 외부는 지방 Fig. 2. Histological observations is cross section of the stomach in sevenband grouper. A: Cross section of the stomach in sevenband grouper. B, C:
Cross section of the stomach of mucosa. Cm: Circular muscle layer, Lm: Longitudinal muscle layer, M: Mucosa, Sg: Secretory granule.
A B
C
Fig. 3. Histological observations is cross section of the anterior, mid, posterior intestine in sevenband grouper. Cm: Circular muscle layer, Lm:
Longitudinal muscle layer, Gc: Goblet cell.
Foregut Midgut Hindgut
Muscle layer
Mucosal fold
Striated border
덩어리가 전체적으로 싸여 있었다(Fig. 1B). 장은 나선의 꼬임 형태를 보였고, RLG(relative length of gut)는 평균 0.87±0.2 (n=10)로 관찰되었다.
2. 소화관의 조직학적 형태 1) 위
위의 점막은 주름에 분지가 형성되지 않고 규칙적이었으며, 내강 쪽으로 뻗어진 형태였다(Fig. 2A). 점막하층을 구성하는 결합조직은 식도의 점막하층보다 치밀한 구조였으며, 장막가 지로 얇은 막의 형태로 위를 감싸고 있었다. 근육층은 점막 하 층과 장막 사이에 위치하여 민무늬근 섬유로 이루어져 있었고, 구조상 외부로부터 종주근층, 환상근층으로 구성되어 있었으 며, 바깥쪽의 종주근층은 내강의 환상근층보다 더 얇았다(Fig.
2A). 위의 점막상피부분과 점막하층부분에는 PAS(periodic
acidSchiff stain) 양성반응을 보이는 당단백질 계통의 분비과 립이 다수 관찰되었다(Fig. 2B, C).
2) 장
장의 점막주름은 위의 점막주름과 유사한 형태로 관찰되었 으나 외부형태 및 점막주름의 형태와 길이 근육층의 두께, 상 피층의 발달 정도 등 내부구조에 의해서 크게 전장부, 중장부, 후장부로 구분할 수 있었다(Fig. 3).
전장부의 경우 점막주름은 불규칙한 형태로서 분지의 형성 이 뚜렷하였고, 점막주름의 길이는 장 전체 중 가장 짧았으며, 상피세포층에 배상세포의 분포도 적었다. 근육층은 종주근층 이 환상근층보다 얇게 관찰되었고, 후장부의 경우 점막주름의 길이와 배상세포의 분포도가 장 전체에서 가장 길고, 많았으 며, 중장부의 경우 근육층, 점막주름의 길이 및 배상세포의 분 포도 등이 전장부와 후장부의 중간적인 성향을 나타내었다.
3. 소화관의 발달
자치어의 형태발달 단계에서 전기 자어기는 전장 1.74~
2.45mm, 후기 자어기는 2.54~17.3mm까지 치어기는 전장 21.3~42.6mm까지 나누어 조사하였으며, 각 단계별 소화기관 형태학 및 조직학적 발달 양상은 다음과 같다.
1) 전기 자어기
부화 직후의 전기 자어는 전장 1.28~1.95mm(평균 1.74±
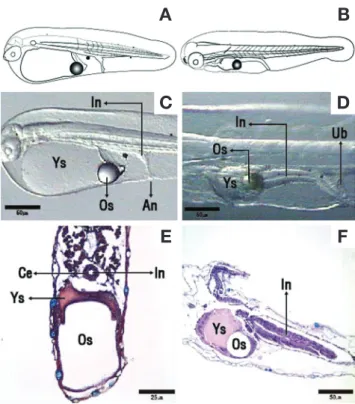
0.24mm, n=10)로 입과 항문은 열려 있지 않았고, 1개의 유구 를 가지고 있었으며, 지느러미는 막상으로 되어 머리 뒷부분의 등쪽 정중앙선을 따라 꼬리와 난황을 지나 머리 아랫부분까지 연결되어 있었다(Fig. 4A). 소화관은 난황을 따라 일직선상으 로 관찰되었다(Fig. 4C). 내강은 좁고, 점막주름은 아직 형성되 지 않았으며, 상피층은 입방상피로 관찰되었다(Fig. 4E).
부화 후 2일째 전기 자어는 전장 1.75~2.36mm(평균 2.05±
0.21mm)로 입과 항문은 완벽하게 열리지 않았고, 꼬리지느러 미의 분화가 시작되었다(Fig. 4B). 소화관의 내강은 전에 비하 여 넓어져 있었으며(Fig. 4D), 후방부의 상피층을 구성하는 세 포들은 원주형으로 나타났다(Fig. 4F).
부화 후 3~4일째 전기 자어(Fig. 5A)는 육안적인 관찰은 어려웠지만 조직학상으로 난황이 관찰되었으며, 그 옆으로 간 조직이 발달하였다(Fig. 5D). 식도와 장이 구분되었고, 일직선 상이었던 장은 배 쪽으로 90° 기울이며 장과 직장으로 나누어 졌다(Fig. 5B). 장과 직장 사이에는 판막이 발달하였고, 점막층 구상세포의 증식과 함께 점막주름의 형성이 관찰되었다.
2) 후기 자어기
부화 후 5일째 후기 자어는 전장 2.40~2.49mm(평균 2.45±
0.03mm)로 입과 항문이 열리면서 아래턱이 발달하였다(Fig.
5C). 식도에서는 점막주름이 형성되었고, 위의 초기 위체부가 형성되었다. 소화관의 내강은 4일째에 비하여 넓어지고, 점막
C D
E F A B
Fig. 4. Histological observations of the digestive system during the development of sevenband grouper larvae. A: General view of the new hatched larvae; B: Larvae past 2 days after hatching; C: Pos
terior region of the intestine new larvae 1 days after hatching; D:
Posterior region of the intestine new larvae 2 days after hatching; E:
Cross section of the digestive system of new hatched larvae, cuboidal epithelium is shwon in digestive system; F: Sagittal section of the digestive system the new larvae 2 days after hatching. HE, ABPAS.
An: Anus; Ce: Cuboidal epithelium; In: Intestine; Os: Oil globule;
Ub: Urine bladder; Ys: Yolk sac.
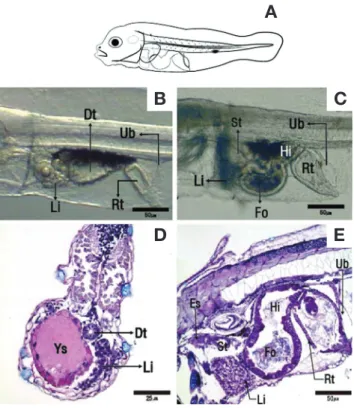
주름을 형성하였으며, 상피층에는 선조연이 형성된 원주세포 들이 관찰되었다(Fig. 5E). 이 시기의 자어를 해부한 결과 몇 몇 자어의 장내에서 로티퍼의 난이 발견되었다.
부화 후 6일째 식도는 근육층이 구분되었으며, 위의 위체부 는 4일째에 비하여 넓어지면서 점막주름을 형성하였고, 공급 한 먹이는 주로 장내에서 발견되었다. 소화관은 배 쪽으로 1회 전 꼬이게 되면서 전장과 후장으로 나누어졌다. 직장은 후장 에 비하여 선조연의 길이가 2배 이상 길어져 두 관의 차이를 관찰할 수 있었다.
부화 후 8일째 후기 자어는 전장 2.48~2.59mm(평균 2.54±
0.05mm)로 유구는 체내에 거의 흡수되었고, 배지느러미에는 양쪽으로 2개의 가시가 생겼으며, 등지느러미에 1개의 가시가 형성되기 시작했다. 이 시기에는 흑색소포와 간이 소화관을 덮기 시작했다.
부화 후 11일째 후기 자어는 전장 3.77~4.67mm(평균 4.27±0.33mm)로 꼬리 부분의 척색 말단은 45°로 굽어지기 시작하였고, 등지느러미에는 2개의 가시가 형성되었고, 그중
두 번째 가시가 가장 길게 발달하였으며, 배지느러미에 있는 2 개의 가시도 길게 발달하였다(Fig. 6A). 식도는 장막, 근육층, 점막하층, 점막층의 구분이 가능해지고, PAS 반응을 나타내는 배상세포가 확인되었다(Fig. 6B, C). 장은 전장, 중장, 후장, 직 장으로 나누어지게 되고, 중장부터 직장까지 PAS 반응을 나타 내는 배상세포와 선조연이 관찰되었다(Fig. 6D, E). 이 시기에 수조 전체의 70%가 폐사가 발생하였으며, 그중 60%가 변태를 완료한 개체였다.
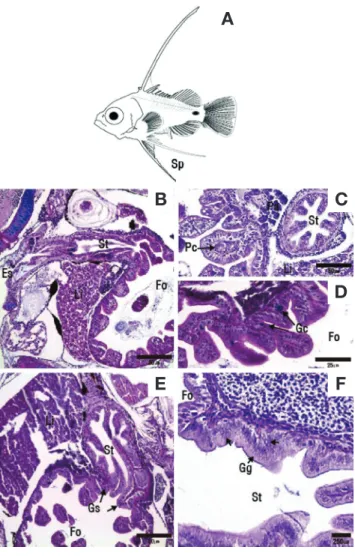
부화 후 19일째 후기 자어는 전장 4.79~5.99mm(평균 5.39±0.41mm)로 공식이 시작되었으며, 그로 인해 개체의 성 장 차이가 관찰되었다. 막으로 된 지느러미가 등지느러미 가 시 3번째 끝에서부터 꼬리를 따라 항문까지 연결되어 있었고, 뒷지느러미가 분화하기 시작하였다(Fig. 7A). 조직학적으로 위는 내부가 팽창되면서 분문위, 위체부, 유문위의 구분이 가 능해졌으나 위선은 관찰되지 않았고(Fig. 7B), 전장과 연결부
B C
D E A
Fig. 5. Sagittal section and cross section of the digestive system during the development of sevenband grouper larvae. A: Larvae past 3~5 days after hatching; B: Incipient intestine larvae past 3 days after hatching; C: Incipient intestine larvae past 5 days after hatching;
D: Cross section of the digestive system of larvae past 3~5 days after hatching; E: Sagittal section of the digestive system of larvae past 5 days after hatching. HE, ABPAS. Dt: Digestive tract; Es: Esophagus;
Fo: foregut; Hi: Hindgut; Li: Liver; Rt: Rectum; St: Stomach; Ub:
Urine bladder; Ys: Yolk sac.
B C
D E A
Fig. 6. Sagittal section and cross section of the digestive system during the development of sevenband grouper larvae. A: Larvae past 11~12 days after hatching; B: Sagittal section of the digestive system of larvae past 11~12 days after hatching; C: Goblet cells shown in esophagus; D: Goblet cells shown in midgut; E: Goblet cells shown in rectum. HE, ABPAS. Es: Esophagus; Fo: Foregut; Gc: Goblet cell; Hi: Hindgut; Li: Liver; Mi: Midgut; Pa: Pancreas; Rt: Rectum;
Sp: Spine; St: Stomach.
위에 초기 장의 형태와 유사한 유문수가 분화되었다(Fig. 7C).
전장에서는 점막층의 선조연 길이가 부화 후 11일째에 비하여 길어졌고, 배상세포가 관찰되어 장 전체에서 배상세포를 관찰 할 수 있었다(Fig. 7D).
부화 후 22일째 위의 위체부가 식도관보다 2배로 팽창되었 다. 이때 위체부 내에서 먹이생물을 관찰할 수 있었고, 위의 유문부에서는 위문괄약근이 형성되었다(Fig. 7E). 부화 후 28 일째 위 내부에는 위선이 분화되었으나 조직화학적으로 위선 의 기능 여부를 확인할 수 있는 분비과립은 관찰되지 않았다 (Fig. 7F).
부화 후 38일째 후기 자어는 전장 12.2~17.1mm(평균 14.4±1.85mm)로 등지느러미의 두 번째 가시와 배지느러미 첫 번째 가시가 길게 발달하였다(Fig. 8A). 조직학적으로 위는 위체부의 넓이가 전장보다 넓어졌으며, 위액과 소화되고 있는 먹이생물들이 발견되어 이 시기에는 성어의 위 기능과 유사하 였다(Fig. 8B). 위선을 빼곡히 가진 위 점막이 두텁게 증가하 였으며, 분비과립이 형성되어 위액의 분비를 확인할 수 있었 다(Fig. 8C). 위의 유문부에 위치한 유문수는 분지를 형성하여 조밀하게 증가하고, 내강에는 점막주름이 발달하면서 유문수 또한 성어와 유사한 형태를 가지고 있었다(Fig. 8D, E). 부화 후 38일부터 치어기로 이행하는 동안 위 내강은 넓어지고, 위
B C
D E A
Fig. 8. Sagittal section and cross section of the digestive system during the development of sevenband grouper larvae. A: Larvae past 30~39 days after hatching; B: Sagittal section of the digestive system of larvae past 30~39 days after hatching; C: Increased gastric gland shown in stomach; D: Pyloric caeca shown in stomach; E: Sagittal section of the pyloric caeca mucosa. HE, ABPAS. Es: Esophagus;
Fo: Foregut; Gg: Gastric gland; Li: Liver; Mi: Midgut; Pc: Pyloric stomach; Sp: Spleen; St: Stomach.
B C
E F A
D
Fig. 7. Morphological observations and sagittal section and cross section of the digestive system during the development of sevenband grouper larvae. A: Larvae past 19~23 days after hatching; B: Sag
ittal section of the digestive system of larvae past 19~23 days after hatching; C: Pyloric caeca shown in stomach; D: Goblet cells shown in foregut; E: Sagittal section of the digestive system of larvae past 25~28 days after hatching; F: Gastric gland shown in stomach. HE, ABPAS. Es: Esophagus; Fo: Foregut; Gc: Goblet cell; Gg: Gastric gland; Gs: Gastric outlet sphincter; Li: Liver; Pa: Pancreas; Pc: Py
loric stomach; St: Stomach.
선의 숫자와 장내 점액세포 및 점막주름은 지속적으로 증가하 거나 길어졌으며, 조직화학적인 변화는 관찰되지 않았다.
고 찰
어류의 소화관은 다른 척추동물과 같이 소화된 먹이로부터 성장에 필요한 대사물질과 에너지를 얻는 과정인 소화와 흡 수 기능을 담당하고 있다(Tanaka, 1969). 장의 길이는 식성과 밀접한 관계가 있으며, Takeuchi(1991)는 어류의 전장에 대한 상대소화관 길이의 비(RLG)를 식성과 비교하였다. 육식성 어 류의 소화관 RLG는 평균 1~2.0으로 뱀장어 Anguilla japon
ica 0.46, Gobio gobio는 0.68로 보고되었고, 초식성 어류의 경 우 RLG가 평균 5.0 이상으로 초식성 어류는 육식성어류에 비 하여 소화관의 길이가 길며(Ferraris et al., 1987), Sarotherodon mossambicys(Takeuchi, 1991)는 RLG가 5.28, 능성어의 RLG 는 0.87로 육식성 어류에 가까웠다.
경골어류의 소화계 형성 과정은 두 번의 중요한 분화기가 있는데 첫 번째 시기는 난황 흡수 말기로 섭이를 시작하는 시 기이고, 두 번째 시기는 위선이 완성되어 유문수가 분화하는 시기 즉, 치어로 이행되기 직전으로 보고 있다(Tanaka, 1971).
해산어류의 경우 농어 Lateolabrax japonicus, 참돔 Pagrus major, 감성돔 Acanthopagrus schlegeli은 각각 부화 후 4일째 에 초기 소화기관이 형성되며(Tanaka, 1973), 바리과 어류 인 무늬바리 Plectropomus leopardus(Meng et al., 2012)와 자바 리(Hur, 2011)는 부화 후 5일째 첫 섭식이 일어날 때 초기 소 화기관이 성립된다. 능성어는 부화 후 5일째에 식도, 위, 장, 직 장 및 부레가 형성되었고, 초기 소화기관이 성립되어 다른 해 산어류와 같은 양상을 보였다.
변태는 동물이 부화 후의 발생과 성장 과정에서 일어나는 단순한 대형화가 아니고 현저한 형태의 변화를 지칭하며, 대 부분의 경골어류 자어는 성어와 형태가 다른데 이는 발육과 정 중에서 변태를 통해 형태 변화가 일어난다. 형태적 변화를 갖는 해산어류는 넙치 Paralichthys olivaceus(Han and Kim, 1997), 참가자미 Pleuronectes herzensteini(Han and Kim, 1999) 및 강도다리 Platichthys stellatus(Byun et al., 2007) 등 가자미 목 어류에서 주로 볼 수 있다.
바리과 어류는 후기 자어 시기에 형태 변화가 일어나는데 이 시기는 등지느러미 두 번째 극조와 배지느러미의 첫 번째 극조가 길게 나오기 시작하는 시기를 기준으로 하였고, 형태 변화 시작 시기를 비교해 보았을 때, 능성어는 부화 후 8일째 전장 2.48~2.59mm에 진행되었고, Lee and Go(2003)는 부화 후 9일 평균 전장 2.94mm에 되었으며, 크게 차이가 없었으나 Kitajima et al.(1991)은 부화 후 13일에 평균 전장 3.45mm로
가장 늦은 시기에 형태가 변화하였으며 크기도 큰 편이었는데 이는 관찰자의 시점과 기준이 다른 결과인 것으로 생각된다.
해산어류 흰동가리 Amphiprion percula(Noh, 2011)는 부화 후 9~10일 사이 체색 발달과 함께 형태 변화 과정에 필요한 충분한 에너지원이 공급되어야 생존율이 높아진다고 하였으 며, 볼락 Sebastes inermis의 경우 알테미아 유생만 공급하였을 때 치어기로 이행하는 형태적 변화 과정에서 대량 폐사가 일 어났으나 냉동 코페포다를 공급한 다음 점차 배합사료로 전환 시켰을 때 정상적인 사육이 가능하였다(Kim, 2002). 이에 따 라 어류의 형태 변화가 시작하기 전에는 어떠한 에너지원을 필요로 하며, 에너지원이 부족할 경우 초기 폐사의 원인이 될 수 있다는 것을 알 수 있다. 능성어 자어는 첫 섭식이 일어나 고 형태 변화가 시작되기 전 부화 후 5~7일 전장 2.40~2.48 mm일 때 외부로부터 영양원을 흡수할 수 있는 초기 소화기관 이 완성되므로 이 시기에는 영양학적으로 충분한 활동을 할 수 있는 먹이를 제공해야 형태 변화 시 발생되는 폐사율을 감 소시킬 수 있을 것으로 생각된다.
위선은 고형물질과 분자량이 높은 단백질성 영양분을 소화 시키고, 위액을 분비시키는 역할을 하며, 유문수와 유문괄약근 은 소화 면적을 넓혀주고 소화 속도를 조절해 준다(Lee and Chin, 1999; Hur, 2011). 위선의 발달은 해산어류마다 차이가 있는데 넙치는 눈의 이동이 시작되는 형태 변화 시기에 소화 관의 상피가 결합조직 중에 함몰되기 시작하고, 함몰된 상피세 포가 위선으로 분화하며, 변태 종료 시에는 위액을 생산하게 된다. 그 밖에 다른 해산어류의 위선분화 시기를 비교해 보면 P. maculatofasciatus(Pena et al., 2003)는 부화 후 16일째, P.
erythrinus(Micale et al., 2006)는 부화 후 28일째, M. miiuy (Lian et al., 2007)는 부화 후 25일째, 흰동가리(Umur and Chris, 2008)는 부화 후 11일째, O. bimaculatus(Pradhan et al., 2012)는 부화 후 8일째에 위선이 분화되어 다른 어류에 비해 빠른 시기에 분화하였다. 자바리(Hur, 2011)는 유문괄약근과 유문수가 분화하고 부화 후 30일째에 발달하였고, 무늬바리 (Meng et al., 2012)는 유문괄약근과 유문수가 분화하기 전인 부화 후 16일째에 위선이 발달하였다. 능성어는 부화 후 28일 째에 위선이 발달하였는데 자바리보다는 먼저 발달하였고, 무 늬바리보다는 늦게 발달하여 위선 분화 시기에 차이를 보였 다. 또한 능성어의 위선분화는 유문괄약근과 유문수가 생긴 뒤 분화된 것으로 보아 자바리와 비슷한 발달 양상을 보였다.
배상세포는 소화관 점막표면의 윤활제로서 물리적인 작용 외에 각종 화학적 손상과 소화 효소의 침해로부터 소화관 점 막을 보호하는 역할을 한다(Allen et al., 1986). 바리과 어류인 흉기흑점바리 E. malabaricus(Li et al., 2012)의 경우 배상세포 는 부화 후 3일째 식도에서 나타났고, 자바리(Hur, 2011)는 부 화 후 4일째 식도에서 처음 발견되어 부화 후 20일째 장 전체
에서 나타났다.
능성어는 부화 후 11일째 식도 부근에서 배상세포가 나타났 고, 부화 후 19일째 전장 부근에 나타나면서 장 전체에 배상세 포가 발견되었다.
능성어 후기 자어는 부화 후 19~28일 사이에 장 전체에서 배상세포 형성과 위선, 유문수 및 유문괄약근의 분화가 형태 변화에 소모한 대사물질과 에너지원 등을 보다 능동적이고 효 율적으로 소화 흡수할 수 있도록 발달되는 것으로 보여진다.
육식성 어류의 경우 성장함에 따라 동족끼리 큰 개체가 작 은 개체를 잡아먹는 공식 현상이 일어나는데 주로 사육밀도가 높고, 공급되는 먹이량이 부족하거나 크기 차이가 많이 날 때 일어난다. 자치어 시기에 일어나는 공식 현상은 양식 어류의 초기 생존율과 대량 폐사를 야기시키나 이를 중점적으로 연구 한 사례가 적어 향후 종묘생산 연구과정에서는 반드시 필요한 연구라고 생각된다. 공식 현상이 주로 발생하는 어류는 고등 어 Scomber japonicus(Kim, 2009)와 자주복 Takifugu rubripes (Pyen and Rho, 1970) 및 넙치(NFRDI, 2009) 등이 있다. 또, 넙치는 강도다리에 비해 성장은 빠르지만 자치어 시기에 공식 현상이 일어나 치어기 이후의 생존율은 강도다리보다 낮다. 고등어는 자치어 시기에 성장이 빠르지만 공식이 심하여 초 기 생존율이 낮으며, 자주복은 사육 도중에 상호 체장의 크고 작음에 관계없이 동족 간 꼬리 부분을 공격하는 습성 때문에 대량 폐사가 발생한다. 능성어는 부화 후 19일째 전장 4.79~
5.99mm일 때 공식 현상이 시작되었고, 형태 변화가 일어나는 시기에 1차 대량 폐사가 발생하였으며, 이후 공식 현상이 시작 되는 시기에 2차 대량 폐사가 관찰되었다. 부화 후 38일째에 나타나는 위체부 팽창은 공식으로 인하여 발생한 숙성어와 약 한 개체의 폐사, 공식 현상과 먹이 계열의 변화로 점차 큰 먹 이를 섭취함으로써 위 내부가 팽창된 것으로 생각된다. 또한 이 시기의 소화관 발달과정 중 유문수의 분화, 장 전체 점액세 포의 분포, 유문괄약근 분화 및 위선분화로 인하여 많은 양의 먹이를 요구하게 되고, 이에 공식 현상이 발생되는 것이라 생 각된다.
따라서 능성어의 종묘 생산과정 중 초기 대량 폐사의 원인 을 규명하고 형태 변화가 일어나기 전 먹이생물의 영양강화, 새로운 먹이생물 개발 등 공식을 억제할 수 있는 연구 등 효율 적인 종묘생산을 위한 지속적인 연구가 이루어져야 할 것으로 생각된다.
요 약
이 연구는 능성어(Epinephelus septemfasciatus)의 소화관 발달과정을 형태학적과 조직학적인 방법으로 부화 후부터 60 일 동안 관찰하였다. 먹이 급이는 부화 후 2일부터 20일 동안
Rotifer(Brachionus rotundiformis)와 클로렐라(Chlorella ellip
soidea)를 급이 하였고, 20일부터 Rotifer와 brine shrimp(Arte
mia salina)를 급이 하였으며, 23일부터 Rotifer와 Artemia, 배 합사료를 급이 하였다. 자치어 10마리를 임의로 선택하여 10%
중성포르말린에 고정한 후 형태학적 관찰과 파라핀포매법에 의한 조직학적인 관찰을 하였다. 능성어의 RLG는 평균 0.87 으로 육식성어류의 특성을 보여주었다. 부화 직후의 자어는 입과 항문은 열려있지 않았고, 소화관은 난황을 따라 일직선 상으로 관찰되었다. 부화 후 5일째 후기 자어는 입과 항문이 열리면서 첫 섭식 활동이 관찰되었고, 부화 후 8일째 변태가 시작되었다. 부화 후 11일째 후기 자어 식도는 네 개의 층이 구분 되었으며, 식도와 중장, 직장에서는 배상세포가 관찰되 었다. 부화 후 19일째 후기 자어는 공식이 시작되었으며, 그로 인해 개체의 성장차이가 관찰되었다. 위는 내부가 팽창되면서 분문위, 위체부 및 유문위의 구분이 가능해졌고, 유문수가 분 화되었으며, 장 전체에서는 배상세포를 관찰할 수 있었다. 부 화 후 28일째 위내부에 위선이 분화되었다. 부화 후 38일째 후 기 자어는 위에서 위액이 분비되는 것을 확인할 수 있었고, 유 문수에 점막주름이 확인되었다. 부화 후 38일째에서 치어기로 이행하는 시기 동안 위 내강은 넓어지고, 위선의 숫자와 장내 점액세포, 점막주름은 지속적으로 증가하거나 길어졌으나 조 직화학적인 변화는 관찰되지 않았다. 따라서 본 연구는 형태 학적 및 조직학적 방법을 이용하여 능성어의 소화관 분화 및 발달에 관한 정보를 얻고자 하였으며, 초기 종묘생산과정에서 적절한 먹이투여시기를 확립함으로써 성장 및 생존율 향상에 기초자료를 제공하고자 하였다.
사 사
본 연구는 국립수산과학원 “해수 순환여과양식 시스템 기술 개발” 과제(R2016021)의 연구비 지원에 의해 수행되었습니다.
REFEREncES
Allen, A., D.A. Hutton, A.J. Leonard, J.P. Pearson and L.A. Sellers.
1986. The role of mucus in the protection of the gastroduo
denal mucosa. Scand J. Gastroenteral., 21: 7177.
Byun, S.G., B.I. Lee, J.H. Lee, H.D. Kim, S.U. Park, S.M. Yun, S.Y.
Hwang, Y.C. Kim and H.G. Han. 2007. Egg development and morphological change of larvae and juveniles of the starry flounder, Platichthys stellatus. Korean J. Ichthyol., 19: 350359.(in Korean)
Ferraris, R.P., J.D. Tan and M.C. De La Cruz. 1987. Development of the digestive tract of milk fish Chanos chanos(Forsskaal):
histology and histochemistry. Aquaculture, 61: 241257.
Han, K.H. and Y.U. Kim. 1997. The early life history of the floun
der, Paralichthys olivaceus Ⅰ. Development of egg, larvae and juveniles. Bull. Yosu Nat. Fish. Univ., 11: 10117.(in Korean)
Han, K.H. and Y.U. Kim. 1999. Eggs development and morphology of larvae of the flounder, Limanda herzensteini. Korean J.
Ichthyol., 11: 8693.(in Korean)
Han, K.H., K.S. Kim and S.H. Lee. 2007. Morphological studies on the digestive tracts of the larvae and juveniles of the floun
der, Paralichthys olivaceus. Dev. Reprod., 11: 121125.(in Korean)
Hur, S.W. 2011. Characterization of cholecystokinin and mucus se
creting goblet cell in longtooth grouper, Epinephelus brune- us. Jeju Univ., 85pp.(in Korean)
Kendall, A.W. Jr., E.H. Ahlstrom and H.G. Moser. 1984. Early life history stages of fishes and their characters. In: Moser, H.G. et al.(eds.), Ontogeny and Systematics of Fishes. Am.
Soc. Ichthyol. Herpetol., Spec. Publ., 1: 1122, Allen Press, Lawrence, KS.
Kim, B.W. 2000. Histological study of digestive tract in larvae and juveniles of tiger puffer, Takifugu rubripes. Jeju Univ., 38pp.
(in Korean)
Kim, B.W., O.S. Na, C.B. Park, H.B. Go, B.S. Kang, Y.C. Choi and Y.D. Lee. 2003. Development of the digestive tract in tiger puffer(Takifugu rubripes). Dev. Reprod., 7: 2934.(in Ko
rean)
Kim, D.H. 2009. Technological development of seed production of pacific mackerel, Scomber japonicus. Rep. of Nat. Fish.
Res. Dev. Ins., pp. 3539.(in Korean)
Kim, I.S., Y. Choi, C.L. Lee, Y.J. Lee, B.J. Kim and J.H. Kim. 2005.
Illustrated book of Korean fishes. Kyo-Hak Publishing Co., Ltd., Seoul, pp. 276286.(in Korean)
Kim, J.M. 2002. Studies on the development of marine ranching program in tongyeong. Korea Ocean Res. Dev. Ins., pp.
405502.(in Korean)
Kitajima, C., M. Takaya, Y. Tsukashima and T. Arakawa. 1991. De
velopment of eggs, larvae and juvenile of the grouper, Epi- nephelus septemfasciatus reared in the laboratory. Japan. J.
Ichthyol., 38: 4755.
Lee, J.S. and P. Chin. 1999. Morphology and histology of the diges
tive tract of the black sea bream, Acanthopagrus schlegeli. J.
Korean Fish. Soc., 32: 642648.(in Korean)
Lee, Y.D., H.B. Kim, C.B. Song, S. Rho and J.J. Lee. 1996. Hormo
nal induction of sex reversal in serranid fish, Epinephelus septemfasciatus. J. Aquacult., 9: 1923.(in Korean) Lee, Y.D. and K.M. Go. 2003. Aquatic industrialization by devel
opment of reproductive technology of sevenband grouper, Epinephelus septemfasciatus. Jeju Univ., pp. 4554.(in Kor
ean)
Li, X., L.I. Jian, C. Weiping and G. Hen. 2012. Epinephelus mala- baricus(Bloch & Schneider) larval and juvenile digestive tract mucous cell. Progress in Fish. Sci., 33: 1523.
Lian, Q., J. Zhong and B. Lou. 2007. Histological studies on the de
velopment of digestive system in larval and juvenile Miich- thys miiuy. J. Shanghai Fish. Univ., 16: 212218.
Meng, Q., D. Shaoxiong, X. Xiaojing, S. Minghui, Y. Yingzhe and S.
Yongquan. 2012. Ontogenetic development of the digestive system and growth in coral trout(Plectropomus leopardus).
Aquaculture, 334337: 131141.
Micale, V., M. Garaffo, L. Genovese, M.T. Spedicato and U. Mug
lia. 2006. The ontogeny of the alimentary tract during larval development in common pandora Pagellus erythrinus.
Aquacult., 251: 354365.
NFRDI. 2009. Aquacultural technique development of starry floun- der. Rep. of Nat. Fish. Res. and Dev. Ins., pp. 2637.(in Korean)
Noh, K.E. 2011. Studies on developmental physiology for mass production of clownfishes. Pukyong Nat. Univ., p. 118.(in Korean)
Oh, M.J., I. Takami, T. Nishizawa, W.S. Kim, C.S. Kim, S.R. Kim and M.A. Park. 2012. Field tests of Poly(I:C) immunization withn ervous necrosis virus(NNV) in sevenband grouper, Epinephelus septemfasciatus(Thunberg). J. Fish. Dis., 35:
187191.
Park, J.Y., C.G. Hong, J.K. Cho, M.H. Son, K.H. Han and J.M. Park.
2015. Early osteological development of the larvae and ju
veniles in sevenband grouper, Epinephelus septemfasciatus.
Korean J. Ichthyol., 27: 189198.(in Korean)
Park, S.J. 2013. Ontogenetic development of the digestive system in pacific mackerel, Scomber japonicus larval and juvenile.
Gyeongsang Univ., 26pp.(in Korean)
Pena, R., S. Dumas, M. Villalejo and J.L. Ortíz. 2003. Ontogenetic development of the digestive tract in reared spotted sand bass Paralabrax maculatofasciatus larvae. Aquaculture, 219: 633644.
Pradhan, P.K., J. Jena, G. Mitra, N. Sood and E. Gisbert. 2012.
Ontogeny of the digestive enzymes in butter catfish Ompok bimaculatus(Bloch) larvae. Aquacult., 38: 16011617.
Pyen, C.K. and S. Rho. 1970. Breeding of the puffer fugu rubripes.
Bull. Korean Fish. Soc., 3: 5264.(in Korean)
Shein, N.L., H. Chuda, T. Arakawa, K. Mizuno and K. Soyano.
2004. Ovarian development and final oocyte maturation in cultured sevenband grouper Epinephelus septemfasciatus.
Fish. Sci., 70: 360365.
Song, Y.B., H.J. Beak, H.B. Kim, K. Soyano, S.J. Kim and Y.D. Lee.
2008. Induction of maturation and ovuration with HCG treatment in the sevenband grouper Epinephelus septemfas- ciatus. J. Aquacult., 21: 96101.(in Korean)
Takeuchi, T. 1991. Digestion and nutrition. In: Itazawa, Y. and I.
Hanyu(eds.), Fish physiology, Koseisha Kosei kaku Tokyo, pp. 67101.
Tanaka, M. 1969. Studies on the structure and function of the diges
tive system in larvae at the stage of first feeding. Japan. J.
Ichthyol., 16: 164174.
Tanaka, M. 1971. Studies on the structure and function of the diges
tive system in teleost larvae Ⅲ. Development of the diges
tive system during postlarval stage. Dep. Fish. Agric. Kyoto Univ., 18: 164174.
Tanaka, M. 1973. Studies on the structure and function of the di
gestive system of teleost larvae. Ph. D. Thsis Kyoto Univ.,
136pp.
Umur, N. and L. Chris. 2008. Ontogeny of the digestive tract of larval percula clownfish, Amphiprion percula(Lacepede 1802): a histological perspective. Aquaculture, 39: 1077
1086.