- 5 -
The Role of Cytokines in Allergy
Chul Hee Lee, M.D.
ABSTRACT
Several pathophysiologic features of allergic inflammation, such as TH differentiation, the regulation of IgE, eosinophilia, mast cell proliferation, and cellular recruitment, are regulated by various cytokines. It has been of particular interest to investigate the underlying mechanism in the preferential activation of TH2 cells by allergens. Although interleukin (IL)-4 is the major dete- rminant of TH2 differentiation, the original cellular source of IL-4 and the nature of the interaction between IL-4 and TH2 differ- entiation remain unclear. Recent studies have demonstrated that the regulation of IgE depends primarily on the functional activities of IL-4, IL-13, and IFN-γ. Eosinophilia and an increased number of mast cells characterize allergic inflammation, which is a T cell-dependent process. IL-5 is the major chemotactic and activating factor of eosinophils. Mast cell proliferation results from several cytokines, including IL-3, IL-9, and IL-10. It has been suggested that, in the late phase reaction of allergic inflammation, proinflammatory cytokines released from mast cells, eosinophils, and TH2 cells enhance the expression of adhesion molecules and chemokines that further promote the allergic cellular milieu.
KEY WORDS:TH differentiation·IgE·Eosinophil·Mast cell·Cytokine.
INTRODUCTION
Over the last decade, there has been enormous growth in the body of knowledge regarding the biology of cytokines in IgE- mediated allergic inflammation. This body of information now provides a framework for our understanding of the pathogen- esis of chronic allergies and may in the future lead to new th- erapeutic strategies in the management of allergic reactions.
This article will review several important concepts regar- ding the role of cytokines in allergic inflammation and will focus on TH2 differentiation, IgE production, major effector cells including eosinophils and mast cells, and cellular recru- itment.
TH1 / TH2 differentiation and the regulation of IgE
One of the most important elements to clarify in understa- nding the causes of allergic disorders is the TH1/TH2 different- iation process that occurs in response to the allergens. Although genetic predisposition, the specific physiochemical nature of the offending allergen, different antigen-processing and pres-
entation pathways, the type of antigen-presenting cells, and other factors play important roles in TH1/TH2 differentiation,1) one of the most critical determinants of TH differentiation is the nature of the cytokine milieu in which the T lymphocytes are activated (Fig. 1).
It has been demonstrated in mice that there are at least two types of TH cells, each with different patterns of cytokine sy- nthesis.2) TH1 cells produce interleukin (IL)-2, tumor necrosis factor (TNF)-β, and interferon (IFN)-γ, which are key ef- fector cells in cell-mediated immunity and delayed-type hy- persensitivity reactions. TH2 cells, on the other hand, secrete IL-4, which stimulate IgE and IgG1 production, IL-5 (eosin- ophil-activating factor), IL-10, and IL-13, which together with IL-4 inhibit certain macrophage functions.3)4) It has been shown recently that TH1 and TH2 cells also function in vivo. Although the source of several cytokines, such as IL-2, IL-6, IL-10 and IL-13, appear to be less limited in humans, there is a general agreement of the existence of human TH cells that are compa- rable to murine TH1 and TH2 cells.
The major determinant of TH2 differentiation is IL-4.5)6) It has been demonstrated that concomitant exposure of T cell receptor transgenic mice to IL-4 and its relevant antigen induces an increase in the production of IL-4 but inhibits development of IFN-γ-producing TH1 lymphocytes.7) However, the orig- inal source of the IL-4 responsible for TH2 differentiation is yet to be clearly defined.
It has been shown that, in contrast to IL-4, IL-12 induces an increase in the concentration of IFN-γ and a decrease in the concentration of IL-4.8)9) In addition to IL-12 and IFN-γ, Department of Otorhinolaryngology-Head Neck Surgery, Seoul
National University, College of Medicine, Seoul, Korea Address correspondence and reprint requests to Chul Hee Lee, M.D., Department of Otorhinolaryngology-Head Neck Surgery, Seoul National University, College of Medicine, 28 Yongon- dong, Chongno-ku, Seoul 110-744, Korea
Tel:82-2-760-3646, Fax:82-2-745-2387 Accepted for publication on March 10, 1998
IFN-α, transforming growth factor (TGF)-β, and IL-1α receptor antagonist (IL-1Ra) have been reported to promote the differentiation of allergen-specific T cells into the TH1 in- stead of TH2 phenotype.4) Cytokines produced by TH1 and TH2
subsets also antagonize each other’s effector functions. TH2
cytokines, IL-4 and IL-10, inhibit a number of TH1 functions, such as the production of IFN-γ.10) However, the TH1-inducing effects of IL-12 can be overcome with the additional presence of IL-4;thus the effects of IL-4 on undifferentiated TH0 cells become dominant.11) The net result is that, in a milieu in which allergic inflammation is present, increasingly extensive alle- rgic responses against bystander antigens can be expected to develop.
Aside from B cell growth and stimulation, IL-4 stimulates the expression of major histocompatibility complex (MHC) class II antigen, B7.1 (CD80), B7.2 (CD86), CD40, surface IgM, and low affinity IgE receptor by B cells, enhancing the antigen-presenting capacity of B cells.1)12) IL-4 also induces a switch in the immunoglobulin isotype from IgM to IgE.13)14) IgE production occurs after the IL-4-induced germ line tran- scription of ε-heavy chain constant region genes. Additionally, B cell activating cytokines, including TNF-α,15) IL-2,15) IL- 5,16) and IL-6,17) react synergistically with IL-4 in the stimu- lation of IgE secretion.
Evidence suggests that IL-4 regulates IgE synthesis through B lymphocytes, and the extent to which T-cell clones support IgE production is directly proportional to IL-4 production.18) On the other hand, IFN-γ, IFN-α, TGF-β, IL-8, IL-12 and other cytokines have been shown to inhibit IL-4-induced IgE synthesis.19) In addition to its effects on B cells, IL-4 also in- duces T-cell proliferation, though to a lesser extent than IL-2.
IL-4 directs the differentiation of uncommitted T-cell precur- sors toward TH2 differentiation. As well as by TH, IL-4 may be secreted by natural killer cells, mast cells, and basophils.
IL-13 has IL-4-like biological activities and is considered to have 30% homology to IL-4. IL-13 induces an IgE isotype switch and stimulates human B cell growth in allergic infla- mmation.1)6) Moreover, it also induces adhesion molecules, such as vascular cell adhesion molecule (VCAM)-1, at the
sites of the allergic inflammation, thus contributing to the se- lective accumulation of eosinophils and lymphocytes. IL-13 is distinguished from IL-4 by different mechanisms of tran- scriptional regulation:IL-4 tends to be an earlier and more transient signal in comparison to IL-13. IL-13 is distinguished from IL-4 also by the absence of effects on T lymphocytes.
IFN-γ is an another critically important cytokine in the re- gulation of IgE synthesis.1) It functions as an inhibitor of al- lergic responses by inhibiting the IL-4-mediated expression of low-affinity IgE receptors and IgE isotype switch. The ability of T-lymphocyte clones to support IgE production is inversely proportional to their IFN-γ production, and clones producing both IL-4 and IFN-γ will support IgE production only in the presence of anti-IFN-γ antibodies.14)15) A recent study has shown that allergen challenge in seasonal allergic rhinitis res- ults in no increase in IFN-γ in nasal lavage fluids.20) Additional cytokines that have been reported to inhibit IL-4-induced IgE production are IL-12, TGF-β, and IL-8.21)22)
Cytokine regulation of eosinophils
Eosinophils are major effector cells in allergic diseases. Eo- sinophilia is a T-lymphocyte-mediated response, as demonst- rated by its absence in athymic or T-cell-depleted animals.1)6) In addition to TH lymphocytes, potentially more important sources of IL-4 and IL-5 in allergic inflammation may be eo- sinophils and mast cells. It has recently been reported that the discrepancy between the mRNA and protein expressions of IL-4 and IL-5 may be attributed to their granule storage in gr- anulocytes, while they are secreted by T cells.23)
IL-5 is of particular interest because it is the most important eosinophilopoietin in allergic inflammation. IL-5 induces ma- turation of a homogeneous population of eosinophils when exposed to bone marrow precursors.24) In addition to stimula- ting eosinophil production, IL-5 acts as a chemoattractant to eosinophils, activates mature eosinophils, induces eosinophil secretion, and prolongs eosinophil survival by reducing apo- ptosis.25) Demonstrating the importance of IL-5, it has been shown that the expression of IL-5 mRNA in the nasal mucosa
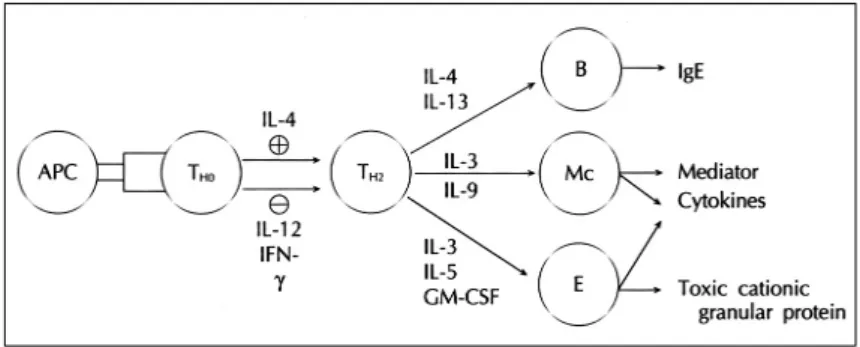
Fig. 1. Major cytokines in allergic inflammation. TH0
cells differentiate into TH2 cells after allergen stimul- ation. IL-4 is a major cytokine of TH2 differentiation, while IL-2 and IFN-γ are the major inhibitors of TH2 di- fferentiation. These two patterns of cytokines antag- onize each other, to contribute to the production of TH2-derived cytokines in allergic inflammation. IL-4 and IL-13 are the major cytokines in IgE synthesis. IL-3 and IL-9 play key roles in mast cell growth and differenti- ation. IL-5 is particularly important for eosinophil dif- ferentiation, survival, and chemotaxis.
of patients with perennial allergic rhinitis increases during na- tural allergen exposure.26)
In addition to IL-5, two colony-stimulating factors, IL-3 and granulocyte-macrophage colony-stimulating factor (GM-CSF), contribute to eosinophil activity in allergic inflammation by prolonging eosinophil survival and generating activated, hyp- odense eosinophils. Recent study further supports the view that GM-CSF is a major cytokine in allergic airway disease.20) In this study, nasal lavage fluid levels of GM-CSF were shown to increase in subjects with allergic rhinitis but not in the su- bjects with the common cold. Additional studies using in situ hybridization have demonstrated that GM-CSF mRNA incre- ases in the nasal mucosa of allergic rhinitis patients after all- ergen provocation in association with tissue eosinophilia.27) RAN-TES (regulated upon activation, normal T expressed and presumably secreted),28) macrophage inflammatory protein (MIP)-1α, monocyte chemotactic peptide (MCP)-3, eotax- in-A and other members of the chemokine family are also in- volved in tissue eosinophilia. Compared to resting eosinophils, cytokine-stimulated eosinophils bind to vascular endothelium more avidly, migrate more rapidly and produce higher levels of newly-synthesized mediators. GM-CSF stimulates stem cell proliferation and activates eosinophils to prolong their survi- val.29) The production of some chemokines, such as RANTES and eotaxin, by respiratory epithelial cells likely attracts criti- cal effector cells such as eosinophils.30)
Mast cell proliferation
As with elevated IgE concentrations and eosinophilia, an increase in the number of mast cells, which characterizes all- ergic diseases, is a T cell-dependent process. Mucosal mast cells (MCT) proliferate in allergic rhinitis under the influence of TH2 cytokines.31) Mast cell proliferation results from the activities of several cytokines, including IL-3, IL-9, and IL-10.
Prolonged exposure of human bone marrow cells to IL-3 re- sults in a predominance of mast cells. This IL-3-stimulated mast cell proliferation is inhibited by GM-CSF.32) IL-9 is a TH2 lymphocyte-derived T cell growth factor that has been found to function as a mast cell growth factor in mice.33) IL-10 is also a cofactor in mast cell proliferation.
In addition to those factors involved in mast cell prolifer- ation, several cytokines have been shown to induce histamine release from basophils and possibly mast cells. These factors include IL-3 and several chemokines, including RANTES, MIP- 1α, MCP-1, MCP-3, and connective tissue activating peptide- III. Of these, MCP-1 and RANTES are the most effective hi- stamine releasing factors.34)35) IL-8 inhibits cytokine-mediated histamine release by basophils.36) Mast cell degranulation is the critical initiating event of acute allergic symptoms. A variety
of cytokines, including TNF-α, IL-4, IL-5, IL-6, TGF-β and IL-13, may be released from mast cells.37)
Cytokines in chronic allergic inflammation
The mechanism that regulates the persistence of local tissue inflammation in chronic allergic diseases remains unclear. Ho- wever, several cytokine-related factors may play roles.6) First, once initiated, a TH2 cell response inhibits the function and activation of TH1 cells. Cytokines such as IL-4 and IL-13 inhibit the development of TH1-delayed-type hypersensitivity responses and enhance the development of TH2 cells, thus po- larizing the T cell response.37) Second, the prolonged survival of inflammatory cells arising from reduced apoptosis at the inflamed sites may result in the establishment of chronic infl- ammation. Increased productions of GM-CSF and IL-5 likely contribute to decreased apoptosis of monocytes and eosinophils, respectively.38)39) Eosinophil apoptosis has been shown to play an important role in the resolution of airway inflammation in asthma.40) Finally, recent studies on mononuclear cells from atopic asthma cases indicate that allergen-induced immunity activation, which gives rise to high concentrations of IL-2 and IL-4, alters the T cell response to glucocorticoid receptor bi- nding affinity and steroid responsiveness.41)
Cytokines and late phase response (LPR)
The regulation of the LPR is becoming better understood.
One of the most important processes in regulating LPR is the production of cytokines. These cytokines include IL-4 (prod- uced by mast cells), IL-3, IL-4, IL-5 and GM-CSF (produced by TH2 cells), and IL-6 (produced by epithelial cells). Indeed, in situ hybridization studies have demonstrated that mRNA expression is preferential to IL-4 and IL-5 during the late phase at 24 hours.27)42) Elevations in IL-3 and GM-CSF expr- ession have been also observed, with no increases in IL-2 or IFN-γ. Moreover, it has been found that a correlation exists between the expression of TH2 cytokine mRNA, particularly IL-5, and the number of EG2+ eosinophils and contribute to the development of LPR and tissue eosinophilia.
Upon stimulation with proinflammatory cytokines, endot- helial cells express different families of adhesion molecules, starting an adhesion cascade that leads to the transendothelial migration of inflammatory cells.43) However, this process is not allergy-specific and has also been shown in viral rhinitis.44) Allergen exposure induces the expression of E-selectin in en- dothelial cells,and TNF strongly upregulates its expression.44)45) Intercellular adhesion molecule (ICAM)-1 is upregulated by IL-1β, TNF-α, and IFN-γ.45) When stimulated by cytokines such as IL-1, IL-4, IFN-γ, TNF-α, IL-5, and IL-13,46)47) and
chemokines such as RANTES48) and IL-8,35) endothelial cells become adhesive for eosinophils, basophils or neutrophils. The expression of adhesion molecules in epithelial cells rapidly increases following exposure to cytokines (IFN-γ or TNF- α).49) The release of TNF-α from mast cells promotes upr- egulation of leucocyte endothelial adhesion molecules E-selectin, ICAM-1, and VCAM-1.29) The release of mast cell IL-4 enh- ances the upregulation of VCAM-1, an adhesion molecule whose expression is evident in both perennial and seasonal allergic rhinitis.50) VCAM-1 upregulation enhances the adhe- siveness of endothelium for T cells, eosinophils, basophils, and monocytes but not for neutrophils, which is characteristic of allergic reactions. The corelease of IL-5 promotes eosinophil priming and chemotaxis.51)52) It has been suggested that, in the LPR of allergic inflammation, proinflammatory cytokines re-
leased from mast cells, eosinophils, and TH2 cells may enhance the expression of both adhesion molecules and chemoattract- ants to promote eosinophilic and basophilic infiltration.
CONCLUSION
Various cytokines regulate several pathophysiologic features of allergic inflammation, such as TH differentiation, the regu- lation of IgE, eosinophilia, mast cell proliferation, and cellular recruitment. Table 1 summarizes the specific role of allergy- related major cytokines. Of the pathophysiologic features of allergic inflammation, it is most important to understand the mechanism underlying the preferential activation of TH2 cells regulated by cytokines. Although IL-4 is the major determinant of TH2 differentiation, the original cellular source and genetic
Table 1. Cytokines and allergy
Cytokine Activity
TH1/TH2 differentiation IL Stimulates TH2 differentiation
IL Inhibits IL-4
IgE regulation IL-4, IL-13 ε isotype switching
IL-4 Generation of IL-4-producing T lymphocytes Synergize with IL-4, IL-13
IL-2, IL-5, IL-6 Inhibit IL-4, IL-13
IFN-γ, TGF-β Stimulates IFN-γ production by NK cells and T cells
IL-12 Inhibits differentiation of IL-4-producing T lymphocytes
Eosinophilia IL-5, IL-3, GM-CSF Eosinophilopoietins
RANTES, MIP-α, eotaxin, MCP-3 Eosinophil chemotaxis and activation IL-1, TNF Eosinophil activation
Mast cell development, IL-3, IL-9, IL-10 Mast cell growth factors
activations and GM-CSF Inhibits mast cell
proliferation MIP-1α, MCP-1 MCP-3, RANTES
Basophil chemotaxis and histamine release
IL-8 Inhibits histamine release
T-cell growth and activation IL-1, IL-2, IL-4, IL-5 IL-6, IL-7, IL-9, IL-11 IL-12, IL-15
T-cell growth factors
Inflammation IFN-γ, GM-CSF, TNFs IL-1, IL-4, IL-6, IL-8
Neutrophil activating factors GM-CSF, TNFs, IL-1,
IL-3, IL-5, RANTES
Eosinophil activating factors IFN-γ, GM-CSF, TNFs,
IL-1, IL-2, IL-3, IL-4
Macrophage activating factors
Anti-inflammatory IL-10, TGF-β Inhibit cytokine production, monocytes / T-cell functions
Adhesion molecule expression IL-1, TNF Express selectins, VCAM-1, and ICAM-1
IL-4 Expresses ICAM-1 and VCAM-1
IL-3, IL-5, IL-8 GM-CSF
Express integrins
control of IL-4 remain unclear.
REFERENCES
1) Borish L, Rosenwasser LJ. Update on cytokines. J Allergy Clin Immunol 1996;97:719-34.
2) Mosmann TR, Cherwinski H, Bond MW, Giedlin MA, Coffman RL.
Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. Immunol 1986;136:2348-57.
3) Mosmann TR, Coffman RL. TH1 and TH2 cells: Different patterns of lymphokine secretion lead to different functional properties.
Annu Rev Immunol 1989;7:145-73.
4) Romagnani S. Biology of human TH1 and TH2 cells. J Clin Imm- unol 1995;15:121-9.
5) Abehsira-Amar O, Gibert M, Joliy M, Theze J, Jankovic DL. IL-4 plays a dominant role in the differential development of Th0 into Th1 and Th2 cells. J Immunol 1992;148:3820-9.
6) Jirapongsananuruk O, Leung DYM. Clinical application of cytok- ines: New directions in the therapy of atopic diseases. Ann Allergy Asthma Immunol 1997;79:5-20.
7) Seder RA, Paul WE, Davis MM, Fazekas de St Groth BF. The pr- esence of interleukin 4 during in vitro priming determines the ly- mphokine-producing potential of CD4+ T cells from T cell receptor transgenic mice. J Exp Med 1992;176:1091-8.
8) Boguniewicz M, Martin RJ, Martin D, Gibson U, Celniker A, Williams M, Leung DY. The effects of nebulized recombinant int- erferon-gamma in asthmatic airways. J Allergy Clin Immunol 1995;
95:133-5.
9) Jaffe HA, Buhl R, Mastrangeli A, Holroyd KJ, Saltini C, Czerski D, et al. Organ specific cytokine therapy. Local activation of mo- nonuclear phagocytes by delivery of an aerosol of recombinant in- terferon-gamma to the human lung. J Clin Invest 1991;88:297-302.
10) de Waal Malefyt R, Abrams J, Bennett B, Figdor CG, de Vries JE.
Interleukin 10 (IL-10) inhibits cytokine synthesis by human mo- nocytes: An autoregulatory role of IL-10 produced by monocytes.
J Exp Med 1991;174:1209-20.
11) Martin RJ, Boguniewicz M, Henson JE, Celniker AC, Williams M, Giorno RC, et al. The effects of inhaled interferon gamma in normal human airways. Am Rev Respir Dis 1993;148:1677-82.
12) Paul WE. Interleukin-4: A prototypic immunoregulatory lympho- kine. Blood 1991;77:1859-70.
13) Del Prete G, Maggi E, Parronchi P, Chretien I, Tiri A, Macchia D, et al. IL-4 is an essential factor for the IgE synthesis induced in vitro by human T cell clones and their supernatants. J Immunol 1988;140:4193-8.
14) Romagnani S. Regulation and deregulation of human IgE synthesis.
Immunol Today 1990;11:316-21.
15) Coffman RL, Carty JS. T cell activity that enhances polyclonal IgE production and its inhibition by interferon-γ. J Immunol 1986;
136:949-54.
16) Moser R, Groscurth P, Carballido JM, Bruijnzeel PL, Blaser K, Heusser CH, et al. Interleukin-4 induces tissue eosinophilia in mice:
Correlation with its in vitro capacity to stimulate the endothelial cell- dependent selective transmigration of human eosinophils. J Lab Clin Med 1993;122:567-75.
17) Vercelli D, Jabara HH, Arai K, Yokota T, Geha RS. Endogenous IL-6 plays an obligatory role in IL-4 dependent human IgE syn- thesis. Eur J Immunol 1989;19:1419-25.
18) Jabara H, Fu S, Vercelli D. CD40 and IgE synergism between anti-
CD40 monoclonal antibody and interleukin-4 in the induction of IgE synthesis by highly purified human B cells. J Exp Med 1990;
172:1861-4.
19) Leung DYM. Mechanisms of the human allergic response: Clinical implications. Pediatr Clin N Am 1994;41:727-43.
20) Linden M, Greiff L, Andersson M, Svensson C, Akerlund A, Bende M, et al. Nasal cytokines in common cold and allergic rhinitis. Clin Exp Allergy 1995;25:166-72.
21) Gauchat JF, Gascan H, de Wall Malefyt R, de Vries JE. Regulation of germ-line ε transcription and induction of ε switching in cl- oned EBV-transformed and malignant human B cell lines by cyt- okines and CD4+ T cells. J Immunol 1992;148:2291-9.
22) Kiniwa M, Gately M, Gubler U, Chizzonite R, Fargeas C, Delespesse G. Recombinant interleukin-12 supresses the synthesis of immu- noglobulin E by interleukin-4 stimulated human lymphocytes. J Clin Invest 1992;90:262-6.
23) Kay AB, Barata L, Meng Q, Durham SR, Ying S. Eosniphils and eosinophil-associated cytokines in allergic inflammation. Int Arch Allergy Immunol 1997;113:196-9.
24) Clutterbuck EJ, Hirst EMA, Sanderson CJ. Human interleukin 5 (IL-5) regulates the production of eosinophils in human bone ma- rrow cultutures: Comparison and interaction with IL-1, IL-3, IL-6, and GM-CSF. Blood 1989;73:1504-12.
25) Rothenberg ME, Petersen J, Stevens RL, Silberstein DS, McKenzie DT, Austen KF, et al. IL-5-dependent conversion of normodense human eosinophils to the hypodense phenotype uses 3T3 fibroblasts for enhanced viability, accelerated hypodensity, and sustained ant- ibody-dependent cytotoxicity. J Immunol 1989;143:2311-6.
26) Lee CH, Rhee CS, Oh SH, Min YG, Lee MS. Increase in expression of IL-4 and IL-5 mRNA in the nasal mucosa of patients with per- ennial allergic rhinitis during natural allergen exposure. Ann Otol Rhinol Laryngol 1997;106:215-9.
27) Durham SR, Ying S, Varney VA, Jacobson MR, Sudderick RM, Mackay IS, et al. cytokine messenger RNA expression for IL-3, IL-4, IL-5, and granulocyte/macrophage colony-stimulating factor in the nasal mucosa after local allergen provocation: Relationship to tissue eosinophilia. J Immunol 1992;148:2390-4.
28) Lee CH, Lee KS, Rhee CS, Lee SO, Min YG. Distribution of RA- NTES and IL-5 in the allergic nasal mucosa and nasal polyp. Ann Otol Rhinol Laryngol (accepted).
29) Bousquet J, Vignola AM, Cambell AM, Michel FB. Pathophysiology of allergic rhinitis. Int Arch Allergy Immunol 1996;110:207-18.
30) Martin LB, Kita H, Leiferman KM, Gleich GJ. Eosinophils in all- ergy: Role in disease, degranulation, and cytokines. Int Arch All- ergy Immunol 1996;109:207-15.
31) Kawabori Y, Kanai N, Tosho T. Proliferative activity of mast cells in allergic nasal mucosa. Clin Exp Allergy 1995;50:15-22.
32) Bressler RB, Thomson HL, Keffer JM, Metcalfe DD. Inhibition of the growth of IL-3-dependent mast cells from murine bone marrow by recombinant granulocyte-macrophage colony-stimulating factor.
J Immunol 1989;143:135-9.
33) Hultner L, Druez C, Moeller J, Uytterhove C, Schmitt E, Rude E, et al. Mast cell growth-enhancing activity (MEA) is structurally related and functionally identical to the novel mouse T cell growth factor P40/TCGFIII (interleukin 9). Eur J Immunol 1990;20:
1413-6.
34) Kuna P, Reddigari SR, Schall TJ, Rucinski D, Sadick M, Kaplan AP. Characterization of the human basophil response to cytokines, growth factors, and histamine releasing factors of the intercrine / chemokine family. J Immunol 1993;150:1932-43.
35) Baggioloni M, Dahinden CA. CC chemokines in allergic inflam-
mation. Immunol Today 1994;15:127-33.
36) Kuna P, Reddigari SR, Kornfield D, Kaplan AP. IL-8 inhibits hist- amine release from human basophils induced by histaminereleasing factors, connective tissue activating peptide III, and IL-3. J Imm- unol 1991;147:1920-4.
37) Baraniuk JN. Pathogenesis of allergic rhinitis. J Allergy Clin Im- munol 1997;99:S763-72.
38) Bratton DL, Hamid Q, Boguniwicz M, Doherty DE, Kailey JM, Leung DY. Granulocyte macrophage-colony stimulating factor co- ntributes to enhanced monocyte survival in chronic atopic dermat- itis. J Clin Invest 1995;95:211-8.
39) Suzuki S, Okubo M, Kaise S, Ohara M, Kasukawa R. Gold sodium thiomalate selectivity inhibits interleukin-5-mediated eosinophil su- rvival. J Allergy Clin Immunol 1995;96:251-6.
40) Woolley KL, Gibson PG, Carty K, Wilson AJ, Twadeu SH, Woolley MJ. Eosinophil apoptosis and the resolution of airway inflammation in asthma. Am J Respir Crit Care Med 1996;154:237-43.
41) Nimmagadda SE, Szefler SJ, Spahn JD, Surs W, Leung DY. All- ergen exposure decreases glucocorticoid receptor binding affinity and steroid responsiveness in atopic asthmatics. Am J Respir Cell Mol Biol 1997;155:87-93.
42) Terada N, Konno A, Fukuda S, Yamashita T, Shirotori K, Okamoto Y, et al. Interleukin-5 gene expression in nasal mucosa and changes in amount of interleukin-5 in nasal lavage fluid after antigen chal- lenge. Acta Otolaryngol (Stockh) 1994;114:203-8.
43) Baroody FM, Lee BJ, Lim MC, Bochner BS. Implicating adhesion molecules in nasal allergic inflammation. Eur Arch Otorhinolaryngol 1995;252:S50-8.
44) Bachert C, Wagenmann M, Holtappels G. Cytokines and adhesion molecules in allergic rhinitis. Am J Rhinol 1998;12:3-8.
45) Bachert C. Hauser U, Prem B, Rudack C, Ganzer U. Proinflamm- atory cytokines in allergic rhinitis. Eur Arch Otorhinolaryngol 1995;
252(Suppl. 1):S44-9.
46) Bochner B, Schleimer R. The role of adhesion molecules in human eosinophil and basophil recruitment. J Allergy Clin Immunol 1994;
94:427-39.
47) Bochner BS, Klunk DA, Sterbinsky SA, Coffman RL, Schleimer RP.
IL-13 selectively induces vascular cell adhesion molecule-1 expr- ession in human endothelial cells. J Immunol 1995;154:799-803.
48) Alam R, Stafford S, Forsythe P, Harrison R, Faubion D, Lett-Brown MA, et al. RANTES is a chemotactic and activating factor for human eosinophils. J Immunol 1993;150:3442-8.
49) Altman LC, Ayars GH, Baker C, Luchtel DL. Cytokines and eosi- nophil-derived cationic proteins upregulate intercellular adhesion molecule-1 on human nasal epithelial cells. J Allergy Clin Immunol 1993;92:95-104.
50) Bradding P, Feather IH, Wilson S, Bardin P, Holgate ST, Howarth PH. Immunolocalisation of cytokines in the nasal mucosa of normal and perennial allergic rhinitis subjects: The mast cell as a source of IL-4, IL-5 and IL-6 in human allergic inflammation. J Immunol 1993;151:3853-65.
51) Bittleman DB, Casale TB. Allergic models and cytokines. Am J Respir Crit Care Med 1994;150:572-6.
52) Howarth PH. The cellular basis for allergic rhinitis. Allergy 1995;
50:6-10.