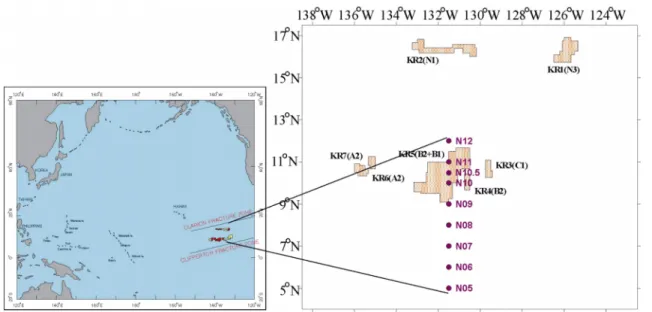
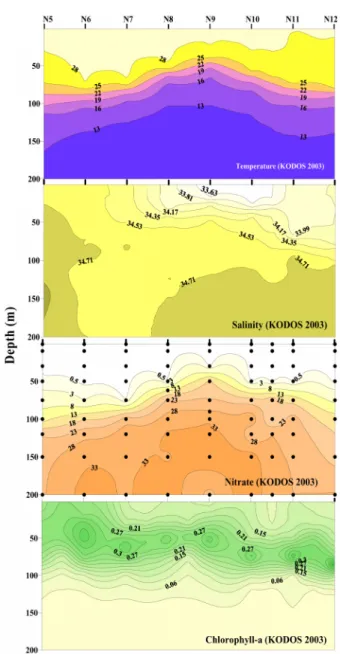
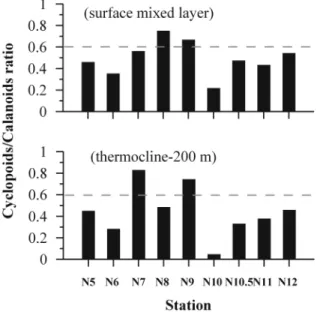
Latitudinal Differences in the Distribution of Mesozooplankton in the Northeastern Equatorial Pacific
10
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
수치

+2

관련 문서
작곡가의 원곡이 지휘자와 연주가에 의해 새롭게 태어나듯이, 우리가 배우고 체득한 모든 이론들과 지식, 테크닉들은 우리 자신에게서 새롭게
2재화 2요소 헥셔-올린 모형에서는 어느 한 경제에서 어느 한 요소의 양이 증가하면, 그 요소를 집약적으로 사용하는 산업의 생산량은 증가하고 다른
Ross: As my lawfully wedded wife, in sickness and in health, until
glen plaids 글렌 플레이드와 캐시미어 카디건, 캐리지 코트, 그리고 케이프 -> 격자무늬의 캐시미어로 된 승마용 바지, 마부용 코트, 말 그림이 수
다양한 번역 작품과 번역에 관한 책을 읽는 것은 단순히 다른 시대와 언어, 문화의 교류를 넘어 지구촌이 서로 이해하고 하나가
The index is calculated with the latest 5-year auction data of 400 selected Classic, Modern, and Contemporary Chinese painting artists from major auction houses..
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
Second, the analysis of the differences in sports participation time showed statistically significant differences in the operation of the competition and
