VOL.24 NO.1 : 001-014 (2011)
苦木의 염증 반응 억제 효과
원광대학교 한의과대학 부인과 교실 류익한, 조한백, 김송백, 서윤정, 최창민
ABSTRACT
The Inhibitory Effect of P icrasmae Lignum on Inflammatory Responses Ik-Han Ryu, Han-Baek Cho, Song-Baeg Kim, Yun-Jung Seo, Chang-Min Choi Dept. of Oriental Gynecology, College of Oriental Medicine, Won-Kwang University Objectives: The purpose of this study was to investigate the anti-inflammatory effects of aqueous extract from Picrasmae Lignum ( PL ).
Methods: To evaluate the anti-inflammatory effects of PL extract, the productions of NO, PGE2 and expression of pro-inflammatory cytokine(IL-1b, IL-6 and TNF-a) were measured in LPS-induced RAW 264.7 cells. Furthermore Western blot analysis has been done to look into the inhibitory mechanisms such as MAPKs and NF-kB.
Results: PL extract down-regulated LPS-induced NO, PGE2, IL-1b, IL-6 and TNF-a productions mainly through JNK and p38 MAPK pathway and NF-kB pathway.
Conclusions: These results suggest that PL extract may be effective for the treatment of inflammatory diseases.
Key Words: Picrasmae Lignum ( PL ), LPS, RAW 264.7 cells, Inflammation
1 )
“이 논문은 2010년도 원광대학교의 교비 지원에 의해서 수행됨.”
“This research was supported by Won-Kwang University in 2010."
교신저자(최창민) : 광주시 남구 주월동 543-8번지 원광대학교 부속 광주한방병원 부인과 전화 : 062-670-6437 이메일 : [email protected]
Ⅰ. 緖 論
염증은 활성화된 면역세포에 의해 일 어나는 선천 면역 계통의 복합적인 반응 으로 1) 감염, 외상, 화학적․기계적․방 사선 및 열에 의한 손상, 과민반응 등에 의해 발생 한다 2) . 국소 순환계의 순환장 애와 혈액 성분의 혈관 외로 삼출 및 조 직 증식이 동반되는 일련의 과정으로 나 타나게 되며, 발적, 종창, 발열, 동통 및 국소 기능장애 등이 나타난다 3) . 이러한 염증 반응은 조직에 가해진 손상을 회복 하기 위한 일련의 과정으로 생명체가 외 부자극에 대한 자기보호를 목적으로 혈 관, 신경, 체액 및 세포를 이용하여 손상 을 국소화 시키고 제거하는 것을 의미 한다 4) . 하지만 염증반응이 너무 강한 경 우 과민성 면역반응, 조직이나 장기의 염증에 의한 지속적인 파괴 및 반흔 형 성, 기능 소실 등 오히려 생체에 불리한 결과를 가져올 수 있다 3) .
여성 생식기에 나타나는 염증 질환은 한의학에서 邪客子門, 熱入血室, 帶下 등 의 기술에서 찾아 볼 수 있는데 이들 질 환을 총칭하는 것이 帶下이다. 帶下라 함 은 생식내강에서 병리적인 상황이 발생 하였을 때 분비되는 출혈 이외의 질 삼 출물을 의미하며, 자궁경부, 질, 난관 등 의 감염에 의한 경우가 많다 5) .
이러한 염증성 질환에서 현재 가장 널리 사용되고 있는 약물인 비스테로이드성 소 염진통제(non-steroidal anti-inflammatory drugs, NSAIDs)는 장기 투여시 출혈성 위장관 궤양을 비롯하여, 신기능 저하, 혈 압 상승 등의 문제점을 안고 있다 6) . 따 라서 기존 약물에 비하여 부작용이 적으
면서 효과적인 항염제 개발이 요구되고 있으며, 천연물들의 약리효과에 대해 관 심이 증대되고 있다.
苦木( Picrasmae Lignum, PL )은 소태 나무 (Picrasma Quassioides (D. Don) Benn. )의 나무껍질을 벗겨낸 木部를 말 한다 7) . quassin, picrasmin 등이 함유되 어 있으며 8) , 항암활성과 면역조절작용을 지니고 있으며, 항고혈압, 항산화, 소염, 해독 작용 등을 갖고 있다고 알려져 있
으며 9,10) , 기존의 연구를 통해 항산화 활
성 11) 및 항암작용 12-14) 이 보고되었다. 또한 苦木은 내복 및 외용으로 습진 및 골반 내조직의 염증 등에도 상당한 효과가 있 었으며, 소염(특히 급성 염증에 대한), 지통 작용이 있다 8) 고 하여 부인과 질환 에의 응용이 가능할 것으로 기대된다.
이에 본 논문에서는 苦木의 항염증 작 용을 실험적으로 규명하기 위하여 lipopolysaccharide(LPS)로 유도된 RAW 264.7 세포에서 nitric oxide(NO), prostaglandin E 2 (PGE 2 ), tumor necrosis factor alpha(TNF-a), interleukin(IL)-1b, IL-6의 생성을 측정하였고, 염증억제 작용 기전을 구명하기 위해 mitogen-activated protein kinase(MAPK) family인 extracelluar signal-regulated kinase(ERK), c-Jun NH 2
-terminal kinase(JNK), p38 MAPK 및 inhibitory kappa B alpha(Ik-Ba)의 발 현을 관찰하여 有意한 결과를 얻었기에 보고하는 바이다.
Ⅱ. 實驗方法
1. 재 료
1) 약 재
실험에 사용한 苦木은 옴니허브(경북 영천)에서 구입 후 원광대학교 한의과대 학 본초학 교실에서 정선하여 사용하였다.
2) 시 약
Fetal bovine serum(FBS), RPMI-1640, penicillin-streptomycin 등의 세포 배양용 시약들은 Gibco BRL(Grand Island, USA) 사에서 구입하였으며, 배양조는 Corning (Rochester, USA)사에서 구입하였다. 실 험에 사용된 시약 중 chloroform, LPS, Tris-HCl, sodium dodesyl sulfate(SDS), acrylamide, bisacrylamide 등은 SIGMA (St. Louis, USA)사에서 구입하였으며, anti-phospho-ERK1/2, anti-phospho-p38, anti-phospho-JNK, anti-Ik-Ba는 Cell Signaling Technology(MA, USA)사에서 구입하였다. Trizol은 invitrogen(Carlsbad, CA, USA)사에서, PCR master mix는 Roche (Basel, Switzerland)사에서, ECL detection 용액은 Amersham(Buckinghamshire, UK) 사에서 구입하였다. 실험에 사용된 모든 시약은 분석용 등급이상으로 사용하였다.
2. 방 법 1) 시료 제조
3차 증류수 1 L에 苦木 100 g을 넣고 2 시간 30 분 동안 전탕한 액을 -80℃에 서 동결 건조시켜 얻어진 분말을 3차 증 류수에 녹여서 filter로 여과하여 사용하 였다.
2) 세포배양
대식세포 계열인 RAW 264.7 세포를 한국세포주은행(KCLB; Seoul, Korea) 으로부터 분양받아 사용하였다. 5% CO 2 , 37℃ incubator 조건하에서 10% FBS와 penicillin-streptomycin이 함유된 RPMI- 1640 배지에 배양하였으며, 2일 마다 10
㎝ dish에 분주하였다.
3) MTT 분석 - 세포독성 평가 RAW 264.7 세포의 생존율은 밀집세 포의 미토콘드리아 탈수소 효소에 의해 자줏빛 formazan 생성물로 변하는 MTT 환원을 바탕으로 MTT 분석법으로 측정 했다. 간단히 설명하면 지수성장을 하는 세포들을 RPMI-1640 배지에서 2 × 10 5 cells/well의 밀도로 현탁하여 여러 농도 (0.05 ㎎/㎖, 0.1 ㎎/㎖, 0.5 ㎎/㎖)로 苦 木 추출물을 처리하였다. 24 시간 동안 배양한 뒤 5 ㎎/㎖의 농도로 배양하기 위해서 MTT 용액을 첨가하고 다시 30 분 동안 배양하였다. MTT-formazan 생성물 은 DMSO 200 ㎕를 첨가함으로써 용해했 다. formazan의 양은 해액(解液)을 96-well plate에 loading하여, spectrophotometer (MD, USA)를 이용하여 540 nm에 흡수 되는 양을 측정함으로서 결정했다.
4) NO 농도의 측정
NO의 기질인 L-arginine은 L-citrulin 과 NO로 변하는데, 이는 빠르게 안정된 이산화질소, 아질산염 및 질산염으로 변 한다. 그리스 시약(Griess reagent: 0.5%
의 sulphanilamide, 2.5%의 phosphoric acid 및 0.5%의 naphtylethylendiamide) 은 아질산염과 화학 반응하여 보라색의 아조염을 형성하는데 이것은 NO의 농도 와 일치한다. 따라서 아조염의 농도로부 터 아질산염의 농도를 측정하여 NO의 농도를 계산한다.
RAW 264.7 세포를 RPMI-1640 배지
에서 2 × 10 5 cells/well의 밀도로 현탁하
여 苦木 추출물을 여러 농도(0.05 ㎎/㎖,
0.1 ㎎/㎖, 0.5 ㎎/㎖)로 1 시간 동안 전
처리 하였다. LPS(500 ng/㎖)로 자극한
후 24 시간 동안 배양한 뒤, 세포 상층액
을 취해 96-well plate에 loading하였다.
100 ㎕의 그리스 시약을 첨가하고, 그 혼 합물의 흡광도를 spectrophotometer로 540 nm에서 측정하였다. NO의 농도는 아질 산염의 표준커브로부터 계산하였다.
5) PGE 2 및 TNF-a, IL-1b, IL-6의 생성량 측정
RAW 264.7 세포를 RPMI-1640 배지 에서 2 × 10 5 cells/well의 밀도로 현탁하 여 苦木 추출물을 여러 농도(0.05 ㎎/㎖, 0.1 ㎎/㎖, 0.5 ㎎/㎖)로 1 시간 동안 전 처리 하였다. LPS(500 ng/㎖)로 자극한 후 24 시간 동안 배양한 뒤, 세포 상층액 에서 PGE 2 및 TNF-a, IL-1b, IL-6을 ELISA 방법으로 정량하였다. ELISA는 Mouse ELISA kit(PGE 2 , TNF-a, IL-1b, IL-6)를 BD pharmingen(CA, USA)사에 서 구입하여 사용하였다.
6) mRNA 발현 측정 (1) RNA 추출
Total RNA는 Trizol 시약을 이용하여 추출하였다. RAW 264.7 세포를 RPMI- 1640배지에서 2 × 10 5 cells/well의 밀도 로 현탁하여 苦木 추출물을 여러 농도 (0.05 ㎎/㎖, 0.1 ㎎/㎖, 0.5 ㎎/㎖)로 1 시 간 동안 전처리 하였다. LPS(500 ng/㎖) 로 자극한 후 24 시간 동안 배양한 세포 를 PBS로 2 회 씻은 다음 PBS를 1 ㎖씩 가해 세포를 포집하고, 원심분리(5000 rpm, 5 분)하여 위의 PBS는 버리고 바 닥에 남은 세포를 TRIzol 용액 1 ㎖를 넣어서 용해시킨다. 100 ㎕의 chloroform 용액을 가하고 두세 번 잘 섞어준 뒤 원 심분리(15,000 rpm, 15 분)하여 맨 위의 상층액을 취하였다. 그 후 2-propanol과 1 : 1로 섞은 뒤 원심분리(15,000 rpm, 10 분)하여 위의 상층액은 버리고 남은
침전물을 80% ethanol로 2 회 씻고 침전 물을 건조시켰다. 침전물에 DEPC 처리 한 증류수를 15 ㎕씩 넣어 RNA를 용해 시키고 정량하였다.
(2) 중합효소 반응
Trizol로 추출한 RNA는 MML-V reverse transcriptase의 protocol을 사용하여 cDNA 로 합성하였다. 역전사 반응을 위하여 total RNA 1 ㎍에 0.5 ㎎의 oligo-(dT)을 넣 고 70℃에서 10 분간 변성시켰다. 그 후 1× single strand buffer, 0.5 mM DTT, 500 mM dNTPs, 200 Unit MMLV reverse transcriptase을 첨가하고 42℃에 서 1 시간 동안 반응시켰다. 그 후에 PCR 은 각각의 tube에 1 ㎕ cDNA, 1× PCR buffer, 1 mM MgCl 2 , 200 mM dNTPs, 0.2 mM의 primer를 넣고 PCR 조건인 92℃에서 30 초, 60℃에서 45 초, 72℃에 서 30 초를 30 cycle 반복하였다. 사용한 primer는 다음과 같다.
iNOS: 5'-AGCCCAACAATACAAAT GACCCTA-3'(forward)
5'-TTC CTG TTG TTT CTA TTT CCT TTG T-3'(reverse)
COX-2: 5'-CACTCAGTTTGTTGAG TCATTC-3'(forward)
5'-GAT TAG TAC TGT AGG GTT AAT G-3'(reverse)
β -actin : 5'-TGT GAT GGT GGG AAT GGG TCA G-3'(forward)
5'-TTT GAT GTC ACG CAC GAT TTC C-3'(reverse)
PCR반응이 끝난 후 4× 샘플링 buffer 를 섞은 뒤 1.5% agarose gel에 10 ㎕씩 을 넣고 전기영동 한 후 자외선을 이용 하여 반응을 확인 하였다.
(3) 정량 중합 효소 반응
mRNA의 발현을 정량적으로 표현하 기 위해 정량 중합 효소 반응을 측정하였 다. 합성된 cDNA 1 ㎕, Real time PCR master mix 4 ㎕, pimer 및 probe를 넣고 PCR 조건으로 반응 시켰다. PCR 조건 은 92℃에서 30 초, 60℃에서 45 초, 7 2℃에서 30 초를 40 cycle로 하였다. 정 량 중합 효소 반응에 쓰인 Forward(f)와 reverse(r) primer 및 TaqMan probe는 Applied Biosystems에서 합성 하였다.
7) Western blot analysis
RAW 264.7 세포를 60 ㎜ culture dish 에 5 × 10 6 cells/dish로 세포를 현탁하고 serum free media(RPMI 1640)로 12 시간 starvation 시킨 후 苦木 추출물를 여러 농도(0.05 ㎎/㎖, 0.1 ㎎/㎖, 0.5 ㎎/㎖)로 1 시간 동안 전처리 하였다. LPS(500 ng/㎖)로 자극한 후 24 시간 동안 배양 한 뒤, cold PBS로 3 회 세척하였다. 농 도별로 cell을 harvest하여 cell을 얻은 뒤 원심분리(5,000 rpm, 5 분)하여 그 상층 액을 버리고 cell pellet을 수거하였다.
RIPA lysis buffer(RIPA buffer 1 ㎖ + phosphotase inhibitor 10 ㎕ + protase inhibitor 10 ㎕)를 넣어 단백질을 lysis시 키고 원심분리(15,000 rpm, 20 분)하여 찌꺼기를 가라앉힌 뒤 단백질을 정량하 였다. 동일한 양의 단백질을 샘플링 버 퍼(4×)를 같이 넣어 섞은 다음 그 샘플 을 10% SDS-PAGE에 전기영동 한 후 5% skim milk로 2 시간 blocking 하였 다. ERK, p38, JNK의 phosphorylation 과 Ik-Ba를 ECL detection 용액으로 확 인하였다.
3. 통계처리
모든 실험 결과는 3회 이상 실시하여
그 평균값을 기초로 Mean ± S.D. 로 나 타내었다. 실험결과에 대한 통계처리는 Oneway ANOVA에 준하였고, p-value가 0.05 미만일 경우 유의한 것으로 판정하 였다.
Ⅲ. 結 果
1. 苦木 추출물의 RAW 264.7 세포에 대한 세포독성
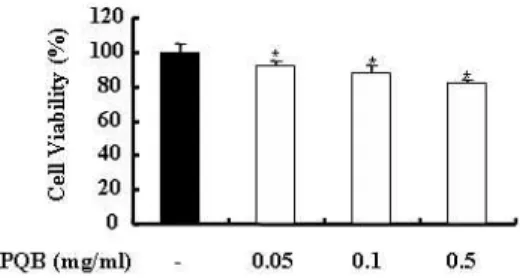
苦木의 세포독성에 관해 알아보기 위 하여 RAW 264.7 세포에 苦木 추출물을 농도 의존적으로 처리하여 24 시간 후에 세포의 생존율을 측정하였다. 그 결과 실험군은 대조군에 비해 세포 생존율을 유의하게 저하시키지 않았다. 즉, 苦木 추출물은 RAW 264.7 세포에 독성을 나 타내지 않았다(Fig. 1).
Fig. 1. Effects of PL extract on cytotoxity in RAW 264.7 cells.
RAW 264.7 cells were incubated with or without
PL
extract as indicated concentration.After 24 hrs, cell viability was measured by MTT assay as described in materials and methods. Data are means of three independent experiment. *P < 0.05 : in comparison with saline. The similar results were obtained from three additional experiments.
2. 苦木 추출물이 NO 생성에 미치는 영향
苦木 추출물의 항염증 작용을 알아보
기 위하여 RAW 264.7 세포에서 LPS 자 극에 의한 NO 생성에 미치는 영향을 조 사하였다.
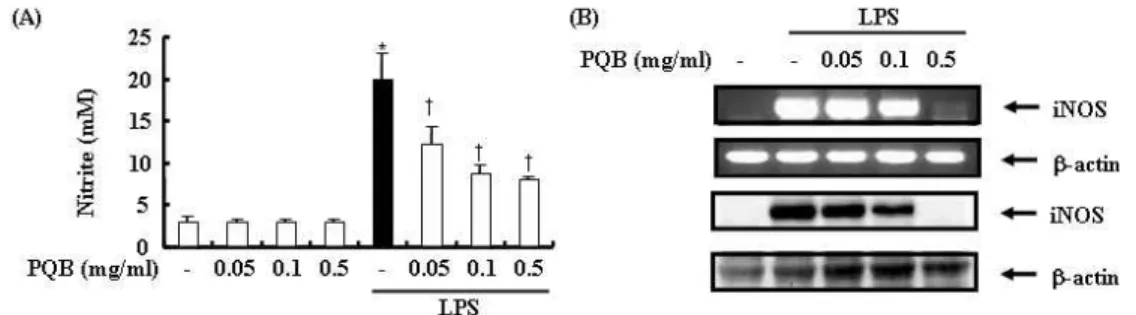
苦木 추출물을 다양한 농도로 전처리 하고 LPS로 자극하였다. 24 시간 후 세 포 상층액에서 NO의 생성을 측정한 결 과 LPS 자극은 대식세포에서 NO의 생 성을 증가 시켰다. 苦木 추출물을 전처
리한 실험군에서는 NO의 생성이 유의하 게 억제되었다(Fig. 2-A).
또한 苦木 추출물이 iNOS mRNA의 발현에 미치는 영향을 조사한 결과 LPS 자극은 iNOS mRNA 발현을 증가시켰 다. 苦木 추출물을 전처리한 실험군에서 는 iNOS mRNA 발현이 억제되었다(Fig.
2-B).
Fig. 2. The inhibitory effect of PL extract on LPS-induced NO production.
The cells were pre-treated with
PL
extract as indicated concentration for 1 hr, and then incubated with or without LPS(500 ng/㎖) for 24 hrs. Detail methods were described in Materials and Methods. (A) NO release was measured by the method of Griess. (B) The level of iNOS mRNA was measured by RT-PCR. *P < 0.05 : in comparison with saline,†
P < 0.05 : in comparison with LPS alone. The similar results were obtained from three additional experiments.3. 苦木 추출물이 PGE 2 생성에 미치는 영향
苦木 추출물의 항염증 작용을 알아보 기 위하여 RAW 264.7 세포에서 LPS 자 극에 의한 PGE 2 생성에 미치는 영향을 조사하였다.
苦木 추출물을 다양한 농도로 전처리 하고 LPS로 자극하였다. 24 시간 후 세 포 상층액에서 PGE 2 의 생성을 측정한 결과 LPS 자극은 대식세포에서 PGE 2
생성을 증가시켰다. 苦木 추출물을 전처 리한 실험군에서는 PGE 2 생성이 유의하 게 억제되었다(Fig. 3-A).
또한 苦木 추출물이 COX-2 mRNA의 발현에 미치는 영향을 조사한 결과 LPS 자극은 COX-2 mRNA 발현을 증가시켰 다. 苦木 추출물을 전처리한 실험군에서는 COX-2 mRNA 발현이 억제되었다(Fig.
3-B).
Fig. 3. The inhibitory effect of PL extract on LPS-induced PGE 2 production.
The cells were pre-treated with
PL
extract as indicated concentration for 1hr, and then incubated with or without LPS(500 ng/㎖) for 24 hrs. Detail methods were described in Materials and Methods. (A) PGE2
release was measured by the method of ELISA. (B) The level of COX-2 mRNA was measured by RT-PCR. *P < 0.05 : in comparison with saline,†
P < 0.05 : in comparison with LPS alone. The similar results were obtained from three additional experiments.4. 苦木 추출물이 IL-1b, IL-6, TNF-a 발현에 미치는 영향
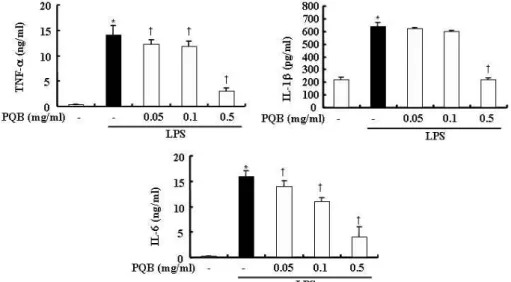
RAW 264.7 세포에서 전염증성 사이 토카인의 발현에 대한 苦木 추출물의 효 과를 조사하기 위하여, 苦木 추출물을 다양한 농도로 전처리하고 LPS로 자극하 였다. 24 시간 후 세포 상층액에서 IL-1b,
IL-6, TNF-a를 측정한 결과 LPS 자극 은 대식세포에서 IL-1b, IL-6, TNF-a 발현을 증가시켰다. 苦木 추출물을 전처 리한 실험군에서는 TNF-a, IL-6 발현이 유의하게 억제되었다. IL-1b의 발현 또 한 억제되었으나 0.5 ㎎/㎖ 농도에서 통 계적인 유의성이 있었다(Fig. 4).
Fig. 4. Effects of PL extract on the productions of IL-1b, IL-6 and TNF-a in RAW 264.7 cells.
The cells were pre-treated with
PL
extract as indicated concentrations for 1 hr, and then incubated with or without LPS(500 ng/㎖) for 24 hrs. Detail methods were described in Materials and Methods. *P < 0.05 : in comparison with saline,†
P < 0.05 : in comparison with LPS alone. The similar results were obtained from three additional experiments.5. 苦木 추출물이 IL-1b, IL-6, TNF-a 의 mRNA 발현에 미치는 영향
苦木 추출물이 RAW 264.7 세포에서 전염증성 인자들을 단백질 수준에서 억 제하였음에 착안하여(Fig. 4), mRNA수 준에서도 전염증성 인자들을 억제하는지 알아보았다.
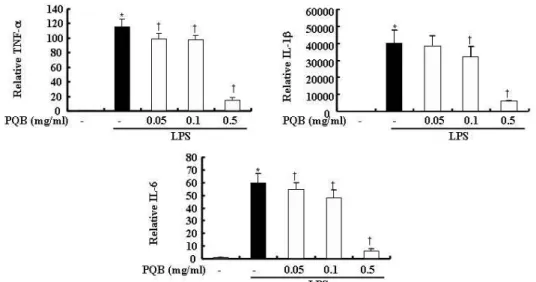
苦木 추출물을 다양한 농도로 전 처리한
후 LPS로 자극하였다. 24 시간 후 IL-1b, IL-6, TNF-a의 mRNA를 측정한 결과 LPS 자극은 mRNA의 발현을 증가시켰 다. 苦木 추출물을 전처리한 실험군에서 는 TNF-a, IL-6의 mRNA 발현이 유의 하게 억제되었다. IL-1b의 mRNA 발현 또한 억제되었으나 0.05 ㎎/㎖ 농도에서 는 통계적인 유의성이 없었다(Fig. 5).
Fig. 5. Effects of PL extract on the mRNA expression of IL-1b, IL-6 and TNF-a.
The cells were pre-treated with
PL
extract as indicated concentrations for 1 hr, and then incubated with or without LPS(500 ng/㎖) for 24 hrs. Detail methods were described in Materilas and Methods. *P < 0.05 : in comparison with saline,†
P < 0.05 : in comparison with LPS alone. The similar results were obtained from three additional experiments.6. 苦木 추출물이 MAPKs 활성에 미 치는 영향
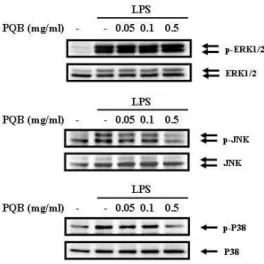
苦木 추출물이 RAW 164.7 세포에서 LPS로 유도된 cytokine의 생성을 억제하 는 기전을 알아보기 위해 MAPK family 인 ERK, JNK, p38의 활성화 정도를 알 아보기로 하였다. MAPKs의 활성화는 각각의 인산화를 통해 이루어진다.
苦木 추출물을 다양한 농도로 전처리 한 후 LPS로 자극하였다. 15 분 후 인산
화 정도를 측정한 결과 LPS로 자극한
RAW 264.7 세포에서는 ERK, JNK, p38
의 활성이 증가하였다. 苦木 추출물을
전처리한 실험군에서는 0.5 ㎎/㎖의 농
도에서 JNK와 p38의 활성이 억제되었
다(Fig. 6).
Fig. 6. Effects of PL extract on the activation of MAP kinase on LPS- stimulated RAW 264.7 cells.
The cells were pre-treated with
PL
extract as indicated concentrations for 1 hr, and then incubated with or without LPS(500 ng/㎖) for 15 mins. Detail methods were described in Materials and Methods.7. 苦木 추출물이 NF-kB의 활성에 미 치는 효과
苦木 추출물이 RAW 164.7 세포에서 LPS로 유도된 cytokine의 생성을 억제하 는 기전을 알아보기 위해 NF-kB의 활 성화 정도를 알아보기로 하였다. NF-kB 의 활성은 Ik-Ba의 분해에 의존하므로 Ik-Ba의 농도를 관찰하여 NF-kB의 활 성을 알아보았다.
苦木 추출물을 다양한 농도로 전처리 한 후 LPS로 자극하였다. 15분 후 Ik-Ba 의 농도를 측정한 결과 LPS로 단독 자 극한 대조군에서는 Ik-Ba의 농도가 감 소하였다. 실험군에서는 苦木 추출물의 농도가 증가할수록 Ik-Ba의 농도가 증 가하였으며, 0.5 ㎎/㎖의 농도에서 대조 군과 비교하여 Ik-Ba의 농도가 높았다 (Fig. 7).
Fig. 7. Effects of PL extract on the activation of NF-kB on LPS-stimulated RAW 264.7 cells.
The cells were pre-treated with