Inhibition of Smooth Muscle Cell Proliferation and Migration by a Talin Modulator Attenuates Neointimal Formation after Femoral Arterial Injury
12
0
0
전체 글
(2) Effects of a Talin Modulator on SMCs. Jong-Ho Kim https://orcid.org/0000-0002-1309-0821 Soon Jun Hong https://orcid.org/0000-0003-4832-6678. formation.1) Therapeutic strategies to ameliorate the progression of atherosclerosis have investigated the inhibition of SMC proliferation and migration using animal models of arterial injury.2)3). Funding This research was supported by a research grant from the Korean Cardiac Research Foundation (201701-01), Korea University in 2016 (K1609881), and by the Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Ministry of Science & ICT (NRF2018M3A9A8017949).. Compelling evidence has revealed that focal adhesions play critical roles in cell proliferation and migration.4)5) Talin is one of the proteins that is found in high concentrations in focal adhesions. Talin binds to the cytoplasmic domain of integrin β subunit, and activates and links integrins to the actin cytoskeleton.6)7) Upon integrin activation, talin recruits other focal adhesion and signaling proteins to the focal adhesion sites.8) A previous study has revealed that depletion of talin using small hairpin RNA resulted in the absence of vinculin, paxillin, and focal adhesion kinase (FAK) at the focal adhesion sites.9) FAK is another key component that also interacts with integrins, and is involved in integrin-mediated signal transduction.10) FAK is a non-receptor tyrosine kinase that contains a focal adhesion-targeting (FAT) domain, which binds to talin and paxillin.11) Autophosphorylation of FAK at tyrosine 397 (Y397) subsequently induces additional phosphorylation at tyrosine residues such as Y925, resulting in the complete activation of FAK.12)13) FAK-Src signaling stimulates the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) pathway involved in cell survival and apoptosis. In addition, the FAT domain also interacts with the growth-factor-receptor-binding protein 2 (GRB2) mediated by Y925 phosphorylation, thereby inducing the activation of the mitogenactivating protein kinase (MAPK) cascade.14)15). Conflict of Interest The authors have no financial conflicts of interest. Author Contributions Conceptualization: Kim JH, Hong SJ; Data curation: Lim IR, Kim C, Jung JW, Kim JH, Hong SJ; Formal analysis: Lim IR, Kim C, Jung JW, Kim JH; Funding acquisition: Hong SJ; Investigation: Lim IR, Kim JH; Methodology: Lim IR, Kim C, Jung JW; Project administration: Kim JH, Hong SJ; Resources: Kim C, Jung JW; Software: Jung JW; Supervision: Kim JH, Hong SJ; Validation: Lim IR, Kim JH, Hong SJ; Visualization: Lim IR, Jung JW; Writing - original draft: Lim IR; Writing - review & editing: Kim JH.. In the present study, we investigated the effects of a newly synthesized talin modulator on SMC proliferation and migration and the underlying mechanism. We further identified that oral administration of the talin modulator inhibited neointimal formation after femoral arterial injury in apolipoprotein E knockout (ApoE KO) mice.. METHODS Synthesis of a talin modulator A mixture of 2-chloroacetamide, N,N-dimethylformamide, K2CO3, and 2-methoxyphenol was stirred overnight. The residue was diluted with ethyl acetate and dried with anhydrous MgSO4. After separation using SiO2 column chromatography with ethyl acetate/n-hexane (1:1), the compound (0.2207 mM) and oxalyl chloride (0.509 mM) in dichloroethane were stirred at 85°C for 5 hours and concentrated under reduced pressure. Then, 4-bromo-3-methoxyaniline (0.221 mM) and methanol were added and stirred for 30 minutes at 0°C, and then overnight at room temperature (RT). The mixture was dried over MgSO4 and concentrated under reduced pressure. The final compound (N-((4-bromo-3-methoxyphenyl)carbamoyl)-2-(2methoxyphenoxy)acetamide; talin modulator) was subjected to SiO2 column chromatography with ethyl acetate/n-hexane (1:3); 1H-NMR (dimethyl sulfoxide [DMSO], 500 MHz) δ 10.72 (s, 1H), 10.31 (s, 1H), 7.49 (d, 1H, J = 7.5Hz), 7.31 (s, 1H), 7.16 (d, 1H, J = 9.0Hz), 7.01 (d, 1H, J = 7.5Hz), 6.96–6.85 (m, 3H), 4.81 (s, 2H), 3.82 (s, 3H), 3.78 (s, 3H); LC-MS ([M+Na]+431), and was finally dissolved in DMSO (D8418, Sigma-Aldrich, St. Louis, MO, USA).. Surface plasmon resonance Surface plasmon resonance (SPR) was performed to confirm the interaction between the talin modulator and talin protein as previously reported.16) Briefly, the response curves were obtained using a ProteOn XPR36 system equipped with a sensor chip and ProteOn XPR36 control software ProteOn Manager v.3.1.0.6 (both from Bio-Rad, Hercules, CA, USA). Serially https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 614.
(3) Effects of a Talin Modulator on SMCs. diluted samples were injected through the channels of the HTE chip for 1 minute at 100 μL/ min, and stopped followed by injection with phosphate buffered saline (PBS) containing 0.005% Tween-20 to obtain the dissociation curves. The response unit of the empty channel was used for normalization.. Cell culture Human aortic SMCs (HAoSMCs; CC-2571) were cultured in SMC growth medium-2 (CC-3182, both from Lonza, Basel, Switzerland) supplemented with 100 U/mL penicillin/streptomycin (#15140, Gibco) at 37°C in a 5% CO2 humidified incubator. After 24 hours of attachement, HAoSMCs were treated with the talin modulator or with an equivalent amount of DMSO (vehicle control). All phase-contrast images were obtained using an upright microscope (DMI 300B, Leica Microsystems, Wetzlar, Germany).. Cell proliferation assay Cell proliferation was assessed by water-soluble tetrazolium salt assay (WST-1 assay; 05015944001, Roche, Basel, Switzerland). HAoSMCs were plated and treated with varying concentrations of the talin modulator or equivalent volume of DMSO. After 24 hours, the cells were incubated with the WST-1 reagent for 2 hours, and absorbance at 450 nm and 655 nm (reference wavelength) was detected using a microplate reader (iMark, Bio-Rad).. Wound healing migration assay HAoSMCs were plated in 6-well plates and allowed to grow to be confluent for 24 hours. A scratch wound was made across the midline of each well using a sterile pipette tip and washed with PBS. The cells were then treated with the talin modulator or DMSO, and phasecontrast images of the wounded area were acquired at 0, 8, 12, and 24 hours after scratching and treatment. The area covered by migrated cells across the wounded line was quantified using Image J software (v1.32, National Institutes of Health, Bethesda, MD, USA).. Western blotting Total protein from HAoSMCs was extracted using cell lysis buffer (#9803, Cell Signaling Technology, Danvers, MA, USA). Equal amounts of the lysate were resolved on 8% or 10% gels by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto polyvinylidene difluoride membranes (10600023, GE Healthcare, Chicago, IL, USA). The membranes were blocked with 5% skim milk in 0.1% tris-buffered saline and 0.1% Tween-20 for 1 hour at RT, and incubated with the following primary antibodies overnight at 4°C: talin (T3287), vinculin (V9131, both from Sigma-Aldrich), paxillin (#610051, BD Biosciences, San Jose, CA, USA), AKT (sc-1618, Santa Cruz, Dallas, TX, USA), p-vinculin (#44-1078G, Thermo Fisher Scientific, Waltham, MA, USA), p-paxillin (#2541), p-FAKY397 (#8556), p-FAKY925 (#3284), FAK (#13009), p-AKT (#9271), p-mitogen-activated protein kinase kinase 1/2 (MEK1/2; #9121), MEK1/2 (#9122), p-extracellular signal-regulated kinase 1/2 (ERK1/2; #9106), and ERK1/2 (#9102, all from Cell Signaling Technology). The membranes were washed and incubated with anti-mouse (#7076), anti-rabbit (#7074, both from Cell Signaling Technology), or anti-goat (HAF109, R&D systems, Minneapolis, MN, USA) HRP-conjugated secondary antibodies for 1 hour at RT. The protein bands were visualized by Clarity Western ECL Substrate (#1705061) and ChemiDoc Touch Imaging System (both from Bio-Rad). All band intensities were normalized to glyceraldehyde 3-phosphate dehydrogenase (G8795, Sigma-Aldrich) and analyzed using Quantity One software (Bio-Rad).. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 615.
(4) Effects of a Talin Modulator on SMCs. Immunofluorescence staining HAoSMCs were fixed with 2% paraformaldehyde (PFA; P6148) and permeablized using 0.1% Triton X-100 (×100, both from Sigma-Aldrich). The cells were blocked with 5% normal goat serum (NGS; #16210, Gibco), and incubated with phosphohistone H3 (PHH3) antibody (06570, Merck Millipore, Burlington, MA, USA) and Alexa Fluor 594-conjugated goat anti-rabbit immunoglobulin G (IgG) (A11012, Thermo Fisher Scientific). Cell nuclei were stained with 4′,6-diamidino-2-phenylindole (D9542, Sigma-Aldrich). Immunofluorescent images were acquired using a BX61 motorized system microscope (Olympus, Tokyo, Japan).. Induction of femoral arterial injury All animal experiments were approved by the Institutional Animal Care and Use Committee of Korea University College of Medicine (KOREA-2017-0126). Male, 10-week-old ApoE KO mice (#002052, The Jackson Laboratory, Bar Harbor, ME, USA) were anesthetized by intraperitoneal (IP) injection of ketamine (80 mg/kg; Yuhan, Seoul, Korea) and xylazine (8 mg/kg; Bayer, Leverkusen, Germany). After a groin incision, the left femoral arteries were injured by passing a sterile wire followed by ligation. ApoE KO mice were randomly assigned to the following 3 groups; sham (subjected to the procedure alone; n=5), vehicle control (DMSO; n=5), and talin modulator (5 mg/kg; n=6). The talin modulator and an equivalent volume of DMSO were prepared in 0.5% carboxymethylcellulose sodium salt (C5678, SigmaAldrich) for administration via oral gavage daily for 28 days. The mice were anesthetized at 7, 14, and 28 days post-procedure and the blood flow in the ventral side was analyzed using a laser Doppler imager (Moor Instruments, Devon, UK).. Histological analyses At day 28, all mice were euthanized by IP injection of ketamine and xylazine, and femoral arteries were harvested. PFA-fixed paraffin-embedded arteries were sectioned in a 5 μm of thickness and stained with hematoxylin and eosin or elastic staining solutions. After de-paraffinization, the tissue sections were incubated with proteinase K solution (#21627, Merck Millipore) for antigen retrieval and blocked with 5% NGS in 0.1% PBST. The sections were then incubated with α-smooth muscle actin (α-SMA; A2547, Sigma Aldrich) antibodies for 1 hour at RT and then with biotinylated anti-mouse IgG antibodies (BA-2000, Vector, Burlingame, CA, USA) for 30 minutes at RT. Subsequently, the sections were incubated with the ABC reagent (PK6100) and peroxidase substrate (SK-4100), and mounted using VectaMount (H-5000, all from Vector). A BX61 motorized system microscope was used to obtain bright-field images.. Enzyme-linked immunosorbent assay Blood serum collected from the abdominal vein of mice was separated using BD Vacutainer SST tubes (367955, BD Diagnostics, Franklin Lakes, NJ, USA). Mouse interleukin-6 (IL-6; M6000B) and tumor necrosis factor alpha (TNF-α; MTA00B, both from R&D Systems) enzyme-linked immunosorbent assay (ELISA) kits were used to evaluate serum levels after 28 days of femoral arterial injury and oral administration of the talin modulator. The ELISA reaction was measured using a microplate reader following the manufacturer's protocol.. Statistical analyses Data are represented as the mean value±standard deviation. Significance of differences was analyzed by 1-way analysis of variance and Student-Newman Keuls test, and results were considered statistically significant when p<0.05. All experiments were repeated at least 3 times independently. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 616.
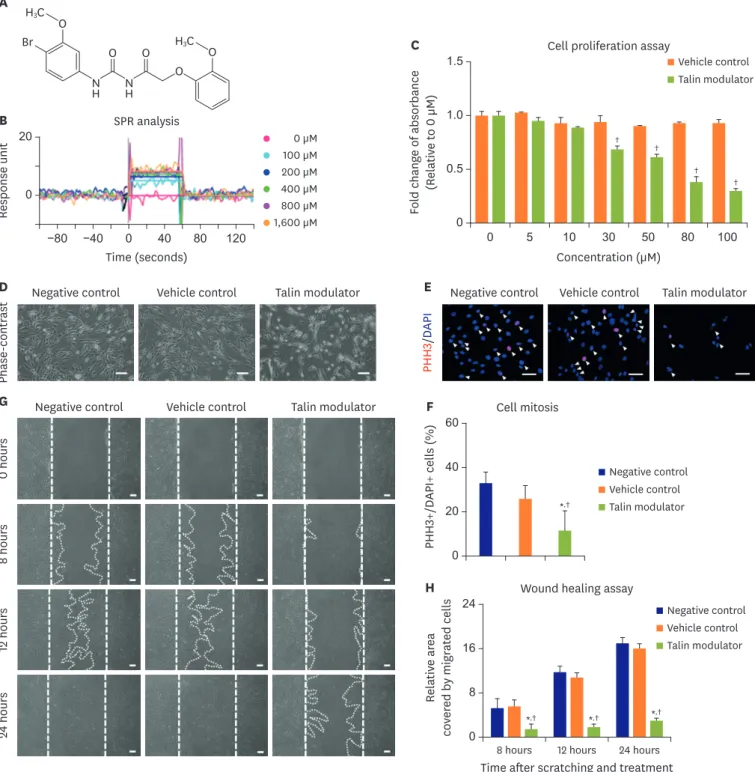
(5) Effects of a Talin Modulator on SMCs. RESULTS A talin modulator reversibly interacted with talin protein In this study, a new talin modulator (N-((4-bromo-3-methoxyphenyl)carbamoyl)-2-(2methoxyphenoxy)acetamide) designed to bind to the talin protein was synthesized (Figure 1A). Interaction between the talin protein and the talin modulator was confirmed using SPR analysis as shown in Figure 1B. The talin modulator bound to the talin protein at all tested concentrations and the binding was reversible.. A talin modulator inhibited cell proliferation and migration of human aortic smooth muscle cells The effect of various concentrations (5–100 μM) of the talin modulator on the proliferation of HAoSMCs was examined by WST-1 assay (Figure 1C). The talin modulator induced a concentration-dependent inhibition of cell proliferation, and the half maximal inhibitory concentration of the talin modulator on cell proliferation was 50 μM (0.612±0.033) relative to the untreated condition (0 μM; 1.0±0.041). After 24 hours of treatment at the concentration of 50 μM, the talin modulator decreased the attached cell density compared to the negative and vehicle controls (Figure 1D). To confirm these results, we also performed immunofluorescence staining for PHH3, a marker for mitosis (Figure 1E). Consistently, the talin modulator significantly decreased the percentage of PHH3-positive cells (11.69±8.83%) compared to the negative and vehicle controls (32.96±5.03% and 25.85±5.93%; Figure 1F). Moreover, the anti-migratory effect of the talin modulator was evaluated by wound healing migration assay (Figure 1G). In the negative and vehicle controls, continuous migration of HAoSMCs was observed in a time-dependent manner, however, the talin modulator significantly suppressed cell migration at 8, 12, and 24 hours (Figure 1H). Therefore, the talin modulator exhibited significant inhibitory effects on cell proliferation and migration of HAoSMCs.. A talin modulator downregulated talin-related downstream molecules We examined the effects of the talin modulator on focal adhesion complexes that mainly consist of integrin, talin, vinculin, paxillin, and FAK. Western blot analyses in Figure 2A and B revealed that the talin modulator did not alter the expression of the talin protein in spite of binding to it (Figure 1B). Intriguingly, the talin modulator dramatically reduced the phosphorylation of both vinculin and paxillin after 24 hours of treatment compared to the negative and vehicle controls (Figure 2C and D). Moreover, paxillin expression was also significantly decreased upon treatment of HAoSMCs with the talin modulator. As FAK is well known as a key downstream molecule in integrin-talin interaction, we next performed experiments to elucidate the underlying mechanisms of inhibition of the talin modulator in HAoSMCs (Figure 2E). The integrin-induced auto-phosphorylation of FAK at Y397 was significantly decreased at 4 hours of treatment with the talin modulator compared to that in the negative and vehicle controls (Figure 2F). The talin modulator also markedly reduced FAK phosphorylation at Y925 in HAoSMCs. Subsequently, the phosphorylation of MEK1/2 and ERK1/2 in the MAPK pathway was significantly decreased in talin modulator-treated cells after 4 hours compared to that in the negative and vehicle controls (Figure 2G), due to decreased FAK phosphorylation at Y925. In addition, a significant reduction in AKT phosphorylation was also observed when HAoSMCs were treated with the talin modulator (Figure 2H). Taken together, these findings suggest that the talin modulator inhibits cell proliferation and migration of HAoSMCs possibly via suppression of focal adhesion molecules and downstream signaling pathways of talin.. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 617.
(6) Effects of a Talin Modulator on SMCs. H3C. O. Br. O N H. Response unit. B. H3C. O. C. O. O. N H. SPR analysis. 20. 0 µM 100 µM 200 µM 400 µM 800 µM 1,600 µM. 0 −80. −40. 0. 40. 80. Cell proliferation assay. 1.5. Fold change of absorbance (Relative to 0 µM). A. Vehicle control Talin modulator. 1.0 †. 0.5. † †. 0. 120. 0. 5. Time (seconds) Negative control. Vehicle control. E. Talin modulator. 30. 50. 80. 100. Negative control. Vehicle control. Talin modulator. PHH3/DAPI Vehicle control. Talin modulator. F PHH3+/DAPI+ cells (%). Negative control. 8 hours. 0 hours. G. 10. Concentration (µM). Phase-contrast. D. †. 40. 0. Relative area covered by migrated cells. 12 hours. Negative control Vehicle control Talin modulator. *,†. 20. H. 24 hours. Cell mitosis. 60. 24. Wound healing assay Negative control Vehicle control Talin modulator. 16 8 * ,†. 0. 8 hours. * ,†. 12 hours. *,†. 24 hours. Time after scratching and treatment Figure 1. Inhibition of proliferation and migration of HAoSMCs by a talin modulator. (A) Chemical structure of the newly synthesized talin modulator. (B) SPR analysis of the talin modulator showing the interaction with a talin protein. (C) Concentration-dependent decrease in cell proliferation by treating HAoSMCs with the talin modulator. The experimental concentration of the talin modulator was set as 50 μM. (D) Phase-contrast images after treatment of HAoSMCs with the talin modulator for 24 hours. (E, F) Immunofluorescence images and quantification of PHH3-positive cells (white arrowheads) in HAoSMCs. (G, H) Wound healing migration assay to evaluate the effects of the talin modulator on cell migration. Cell migration was assessed by quantifying the area covered by migrated cells across the wounded line. All scale bars represent 100 μm. DAPI = 4′,6-diamidino-2-phenylindole; HAoSMC = human aortic smooth muscle cell; PHH3 = phosphohistone H3; SPR = surface plasmon resonance. *p<0.05 vs. negative control at each time point; †p<0.05 vs. vehicle control at each concentration or each time point. All experiments were repeated at least 3 times independently.. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 618.
(7) p-FAK. Y397. https://doi.org/10.4070/kcj.2019.0421. F. NS. *. ,†. l l r ro ro to nt ont ula o c c od ive cle m at ehi lin g V Ta Ne. 0. 0.5. 1.0. 1.5. p-FAKY397. l l r tro tro ato on con dul c o ive cle m at ehi lin g a V T Ne. 0. 0.5. 1.0. 1.5. Talin. C NS. Vinculin. l tro. * * ,†. l r ro to n ont ula o c c od ive cle m at ehi lin g V Ta Ne. 0. 0.5. 1.0. 1.5. p-FAKY925. l l r tro tro ato on con dul c o ive cle m at ehi lin g a V T Ne. 0. 0.5. 1.0. 1.5. 2.0. G. *. ,†. p-Vinculin. l tro. *,†. p-MEK1/2. l r ro to n ont ula o c c od ive cle m at ehi lin g V Ta Ne. 0. 0.5. 1.0. 1.5. l l r tro tro ato on con dul c o ive cle m at ehi lin g a V T Ne. 0. 0.5. 1.0. 1.5. D Paxillin. *. ,†. *. ,†. p-ERK1/2. l l r ro ro to n t on t u la o c c od ive cle m at ehi lin g V Ta Ne. 0. 0.5. 1.0. 1.5. l l r tro tro ato on con dul c o ive cle m at ehi lin g a V T Ne. 0. 0.5. 1.0. 1.5. H. *,†. p-Paxillin. p-AKT. *,†. l l r ro ro to n t on t u l a o c c od ive cle m at ehi lin g V Ta Ne. 0. 0.5. 1.0. 1.5. 0. 0.5. 1.0. 1.5. l l r tro tro ato on con dul c o ive cle m at ehi lin g a V T Ne. Fold change of intensity (Normalization to paxillin). Figure 2. Effects of a talin modulator on focal adhesion molecules and related signaling pathways. (A) Western blot analyses of the expression of focal adhesion-associated molecules including talin (B), vinculin (C), and paxillin (D) in HAoSMCs after 24 hours of treatment with the talin modulator. All band intensities were normalized to GAPDH, and the phosphorylated forms were then normalized to the total form of the respective forms. (E) Western blot analyses of downstream signaling pathways indicating decreased phosphorylation of FAK at Y397 and Y925 (F), MEK1/2, ERK1/2 (G), and AKT (H) by treating HAoSMCs with the talin modulator for 4 hours. All band intensities were normalized to GAPDH, and the phosphorylated forms were then normalized to the total form of the respective forms. AKT = protein kinase B; ERK1/2 = extracellular signal-regulated kinase 1/2; FAK = focal adhesion kinase; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; HAoSMC = human aortic smooth muscle cell; MEK1/2 = mitogen-activated protein kinase kinase 1/2; NS = not significant. *p<0.05 vs. negative control; †p<0.05 vs. vehicle control. All experiments were repeated at least 3 times independently.. l l r tro tro ato on con dul c o ive cle m at ehi lin g V Ta Ne. GAPDH. AKT. p-AKT. ERK1/2. p-ERK1/2. MEK1/2. p-MEK1/2. FAK. p-FAKY925. E. l l r ro ro to nt ont ula o e c le c od tiv hic in m a g Ve al T Ne. GAPDH. Paxillin. p-Paxillin. Vinculin. p-Vinculin. Talin. 2.0. Fold change of intensity (Normalization to GAPDH) Fold change of intensity (Normalization to FAK). B. Fold change of intensity (Normalization to GAPDH). Fold change of intensity (Normalization to FAK). Fold change of intensity (Normalization to vinculin) Fold change of intensity (Normalization to MEK1/2). Fold change of intensity (Normalization to GAPDH) Fold change of intensity (Normalization to ERK1/2). https://e-kcj.org Fold change of intensity (Normalization to AKT). A. Effects of a Talin Modulator on SMCs. 619.
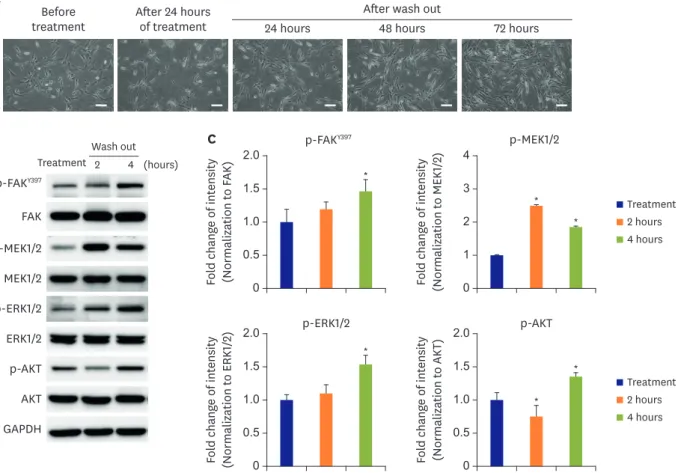
(8) Effects of a Talin Modulator on SMCs. The effect of a talin modulator was reversible in human aortic smooth muscle cells To investigate the reversibility of downstream molecules that were suppressed upon treatment with the talin modulator in HAoSMCs, cells were treated with the talin modulator and then the culture medium was replaced with fresh medium. As shown in Figure 3A, the number of viable and attached cells was decreased after treatment with the talin modulator; however, the proliferative capacity of the cells was restored with no morphological changes after 72 hours of wash out of the talin modulator. Furthermore, the auto-phosphorylation of FAK at Y397 was completely reversed at 4 hours after removal of the talin modulator. The reversible effect of the talin modulator at 4 hours of treatment significantly restored the downregulated phosphorylated levels of MEK1/2, ERK1/2, and AKT in HAoSMCs (Figure 3B and C). These reversible effects are due to the reversible binding of the talin modulator to the talin protein as shown in Figure 1B.. A talin modulator suppressed neointimal formation after femoral arterial injury ApoE KO mice were subjected to left femoral arterial injury to induce abnormal proliferation and migration of SMCs. The mice were daily administered with the talin modulator or. Wash out Treatment 2. p-FAKY397 FAK p-MEK1/2 MEK1/2. 4 (hours). C. 24 hours. 2.0. AKT GAPDH. Fold change of intensity (Normalization to ERK1/2). p-AKT. p-FAKY397 *. 1.5 1.0 0.5 0. p-ERK1/2 ERK1/2. After wash out 48 hours. 2.0 1.5. Fold change of intensity (Normalization to MEK1/2). B. After 24 hours of treatment. p-ERK1/2 *. 1.0 0.5 0. Fold change of intensity (Normalization to AKT). Before treatment. Fold change of intensity (Normalization to FAK). A. 72 hours. 4 3. p-MEK1/2. *. 2. *. Treatment 2 hours 4 hours. 1 0 2.0. p-AKT. 1.5 1.0. * *. Treatment 2 hours 4 hours. 0.5 0. Figure 3. Reversible effects of a talin modulator. (A) HAoSMCs were treated with a talin modulator for 24 hours and then cultured in fresh media for 24, 48, and 72 hours. Scale bars represent 100 μm. (B, C) Western blot analyses of FAK, MEK1/2, ERK1/2, and AKT phosphorylation after treatment, and at 2 and 4 hours after wash out of the talin modulator. All band intensities were normalized to GAPDH, and the phosphorylated forms were then normalized to the total form of the respective forms. AKT = protein kinase B; ERK1/2 = extracellular signal-regulated kinase 1/2; FAK = focal adhesion kinase; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; HAoSMC = human aortic smooth muscle cell; MEK1/2 = mitogen-activated protein kinase kinase 1/2. *p<0.05 vs. treatment. All experiments were repeated at least 3 times independently.. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 620.
(9) Effects of a Talin Modulator on SMCs. vehicle control for 28 days. There was no significant change in body weight among groups throughout the duration of the experiment (data not shown). The blood flow on the ventral side of mice was examined using laser Doppler imager post-surgery (day 0), and at day 7, 14, and 28 (Figure 4A). At 28 days, oral administration of the talin modulator significantly. Day 7. Day 14. Day 28. Talin modulator Vehicle control. Blood flow. 600. * ,†. 500. *. 400 300 200 100. at or. l tro m. od. ul. on. 2. ul. at or. l. od m. Ta l. icl ec. Sh. am. 0. TNF-α. 5 4. *. 3. * ,†. 2 1 0. Ve hi. Sh am cle co Ta nt lin ro l m od ul at or. Talin modulator. * ,†. 4. Ve h Sham. Injured left femoral artery Vehicle control. Concentration (pg/mL). Elastic Uninjured right femoral artery. αSMA. D. in *. 6. tro. Talin modulator. 8. on. Sham. Injured left femoral artery Vehicle control. IL-6. 10. in. Uninjured right femoral artery. Concentration (pg/mL). E. Ta l. Ve h. icl ec. Sh. am. 0. H&E. C. B Average of flux mean at day 28. Day 0. Sham. A. Figure 4. Improvement of the blood flow and suppression of neointimal formation by a talin modulator after femoral arterial injury. (A) Representative images of the ventral side of a mouse measured by laser Doppler imager post-surgery (day 0) and at day 7, 14, and 28 after femoral arterial injury. (B) Quantitative analysis of the blood flow at day 28 after injury and oral administration of the talin modulator. (C) Histological images of left and right femoral arteries showing the reduced neointima after administration of the talin modulator. (D) Immunohistological staining of α-SMA (DAB, brown) to visualize SMCs within the neointima. (E) Serum levels of inflammatory cytokines such as IL-6 and TNF-α after 28 days of injury and administration as measured by ELISA. All scale bars represent 100 μm. α-SMA = alpha-smooth muscle actin; DAB = 3,3′-diaminobenzidine; ELISA = enzyme-linked immunosorbent assay; H&E = hematoxylin and eosin; IL-6 = interleukin-6; SMC = smooth muscle cell; TNF-α = tumor necrosis factor alpha. *p<0.05 vs. sham; †p<0.05 vs. vehicle control. Sham (n=5); vehicle control (n=5); talin modulator (n=6).. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 621.
(10) Effects of a Talin Modulator on SMCs. improved the blood flow compared to that in the sham and vehicle control groups (Figure 4B). Moreover, the wire injury induced increased neointimal formation in the left femoral arteries of the sham group compared to right arteries. Interestingly, mice that had been administered the talin modulator exhibited markedly reduced neointima in femoral arteries compared to that in the sham and vehicle control groups (Figure 4C). Decreased expression of α-SMApositive SMCs was observed by administration of the talin modulator compared to that in the sham and vehicle control groups (Figure 4D). In addition, at 28 days after administration of the talin modulator changes in the expression of proinflammatory cytokines including IL-6 and TNF-α were observed. Talin modulator-administered mice exhibited significantly lower serum levels of IL-6 and TNF-α than the sham and vehicle control groups (Figure 4E).. DISCUSSION The present study demonstrated that a newly synthesized talin modulator exhibited antiproliferative and anti-migratory effects in HAoSMCs, and its effects with respect to the regulation of downstream signaling pathways were reversible. Our study also revealed that oral administration of the talin modulator improved the blood flow and neointimal formation after femoral arterial injury in ApoE KO mice. Talin has been implicated in linking integrin-mediated pathways related to cell proliferation and cell cycle progression. Talin-deficient epithelial cells were unable to recruit other focal adhesion proteins, and showed decreased incorporation of BrdU and 5-ethynyl-2′deoxyuridine, and reduced PHH3 expression via modulating FAK phosphorylation.9) The autophosphorylation of FAK at Y397 induces the phosphorylation of other tyrosine residues including Y925 which interacts with GRB2 of the MAPK pathway.15) It has been reported that mutation of FAK at Y925 reduced cell proliferation, migration, and metastasis along with reduced activation of ERK1/2.17) In addition, FAK has been found to be upstream of the PI3K/ AKT signaling pathway involved in cell survival and apoptosis through its binding sites at Y397.18) In accordance with previous reports, our findings demonstrated that significantly decreased phosphorylation of FAK at both Y397 and Y925 sites (Figure 2E and F) due to treatment with the talin modulator resulted in subsequent downregulation of MAPK and PI3K signaling molecules (Figure 2G and H). These downregulations resulted in a concentration-dependent decrease in cell proliferation and migration, and lower expression of PHH3 in talin modulator-treated cells (Figure 1C-H). Several studies have focused on inhibiting neointimal formation through suppressing the signaling pathways involved in SMC proliferation and migration.3)19)20) In particular, Son et al.21) reported that inhibition of FAK decreased SMC migration and neointimal thickening after balloon injury by reducing the phosphorylation of MAPKs and AKT. Similarly, the present study showed that administration of the talin modulator significantly suppressed neointimal formation and SMC expression within the neointima after arterial injury (Figure 4C and D), via downregulating signaling pathways related to cell proliferation and migration (Figure 2E). A previous study had revealed the inhibitory effects of DMSO on SMC proliferation by suppressing DNA synthesis.22) It has also been reported that 0.5% DMSO treatment showed a partial inhibition of SMC proliferation. In our study, DMSO was used as a solvent for the talin modulator and treatment with 0.5% DMSO as vehicle control partially, but not significantly affected cell proliferation relative to that in the negative control (Figure 1E and F, Figure 2G). https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 622.
(11) Effects of a Talin Modulator on SMCs. Moreover, there were significant differences in the blood flow of injured femoral arteries and in the proinflammatory cytokine levels after administration of DMSO (vehicle control) relative to that in the sham group (Figure 4B and E). These observations are supported by a previous study which reported that injection of 40% DMSO prevented the decrease in the blood flow and thrombotic occlusion in mouse carotid arteries after injury.23) In summary, this study showed that the talin modulator could attenuate neointimal formation after femoral arterial injury. The inhibitory effects of the talin modulator are mediated through suppression of SMC proliferation and migration via modulation of focal adhesion and downstream signaling molecules of talin. Based on our study, the talin modulator may be an effective anti-atherosclerotic drug by suppressing neointimal hyperplasia for the treatment of cardiovascular diseases.. ACKNOWLEDGMENTS The authors thank Chi-Yeon Park and Ji-Hyun Choi for their technical support.. REFERENCES 1. Bennett MR, Sinha S, Owens GK. Vascular smooth muscle cells in atherosclerosis. Circ Res 2016;118:692-702. PUBMED | CROSSREF. 2. Kim JH, Lim IR, Joo HJ, et al. Fimasartan reduces neointimal formation and inflammation after carotid arterial injury in apolipoprotein E knockout mice. Mol Med 2019;25:33. PUBMED | CROSSREF. 3. Nagayama K, Kyotani Y, Zhao J, et al. Exendin-4 prevents vascular smooth muscle cell proliferation and migration by angiotensin II via the inhibition of ERK1/2 and JNK signaling pathways. PLoS One 2015;10:e0137960. PUBMED | CROSSREF. 4. Huttenlocher A, Horwitz AR. Integrins in cell migration. Cold Spring Harb Perspect Biol 2011;3:a005074. PUBMED | CROSSREF. 5. Wozniak MA, Modzelewska K, Kwong L, Keely PJ. Focal adhesion regulation of cell behavior. Biochim Biophys Acta 2004;1692:103-19. PUBMED | CROSSREF. 6. Sun Z, Costell M, Fässler R. Integrin activation by talin, kindlin and mechanical forces. Nat Cell Biol 2019;21:25-31. PUBMED | CROSSREF. 7. Ye F, Kim C, Ginsberg MH. Reconstruction of integrin activation. Blood 2012;119:26-33. PUBMED | CROSSREF. 8. Goult BT, Yan J, Schwartz MA. Talin as a mechanosensitive signaling hub. J Cell Biol 2018;217:3776-84. PUBMED | CROSSREF. 9. Wang P, Ballestrem C, Streuli CH. The C terminus of talin links integrins to cell cycle progression. J Cell Biol 2011;195:499-513. PUBMED | CROSSREF. 10. Harburger DS, Calderwood DA. Integrin signalling at a glance. J Cell Sci 2009;122:159-63. PUBMED | CROSSREF. 11. Chen HC, Appeddu PA, Parsons JT, Hildebrand JD, Schaller MD, Guan JL. Interaction of focal adhesion kinase with cytoskeletal protein talin. J Biol Chem 1995;270:16995-9. PUBMED | CROSSREF. 12. Calalb MB, Polte TR, Hanks SK. Tyrosine phosphorylation of focal adhesion kinase at sites in the catalytic domain regulates kinase activity: a role for Src family kinases. Mol Cell Biol 1995;15:954-63. PUBMED | CROSSREF. 13. Kleinschmidt EG, Schlaepfer DD. Focal adhesion kinase signaling in unexpected places. Curr Opin Cell Biol 2017;45:24-30. PUBMED | CROSSREF. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 623.
(12) Effects of a Talin Modulator on SMCs. 14. Bouchard V, Demers MJ, Thibodeau S, et al. Fak/Src signaling in human intestinal epithelial cell survival and anoikis: differentiation state-specific uncoupling with the PI3-K/Akt-1 and MEK/Erk pathways. J Cell Physiol 2007;212:717-28. PUBMED | CROSSREF. 15. Tomakidi P, Schulz S, Proksch S, Weber W, Steinberg T. Focal adhesion kinase (FAK) perspectives in mechanobiology: implications for cell behaviour. Cell Tissue Res 2014;357:515-26. PUBMED | CROSSREF. 16. Lim IR, Joo HJ, Jeong M, et al. Talin modulation by a synthetic N-acylurea derivative reduces angiogenesis in human endothelial cells. Int J Mol Sci 2017;18:E221. PUBMED | CROSSREF. 17. Kaneda T, Sonoda Y, Ando K, et al. Mutation of Y925F in focal adhesion kinase (FAK) suppresses melanoma cell proliferation and metastasis. Cancer Lett 2008;270:354-61. PUBMED | CROSSREF. 18. Xia H, Nho RS, Kahm J, Kleidon J, Henke CA. Focal adhesion kinase is upstream of phosphatidylinositol 3-kinase/Akt in regulating fibroblast survival in response to contraction of type I collagen matrices via a beta 1 integrin viability signaling pathway. J Biol Chem 2004;279:33024-34. PUBMED | CROSSREF. 19. Kim J, Jang SW, Park E, Oh M, Park S, Ko J. The role of heat shock protein 90 in migration and proliferation of vascular smooth muscle cells in the development of atherosclerosis. J Mol Cell Cardiol 2014;72:157-67. PUBMED | CROSSREF. 20. Daniel JM, Prock A, Dutzmann J, et al. Regulator of G-protein signaling 5 prevents smooth muscle cell proliferation and attenuates neointima formation. Arterioscler Thromb Vasc Biol 2016;36:317-27. PUBMED | CROSSREF. 21. Son JE, Lee E, Jung SK, et al. Anthocyanidins, novel FAK inhibitors, attenuate PDGF-BB-induced aortic smooth muscle cell migration and neointima formation. Cardiovasc Res 2014;101:503-12. PUBMED | CROSSREF. 22. Katsuda S, Okada Y, Nakanishi I, Tanaka J. Inhibitory effect of dimethyl sulfoxide on the proliferation of cultured arterial smooth muscle cells: relationship to the cytoplasmic microtubules. Exp Mol Pathol 1988;48:48-58. PUBMED | CROSSREF. 23. Camici GG, Steffel J, Akhmedov A, et al. Dimethyl sulfoxide inhibits tissue factor expression, thrombus formation, and vascular smooth muscle cell activation: a potential treatment strategy for drug-eluting stents. Circulation 2006;114:1512-21. PUBMED | CROSSREF. https://e-kcj.org. https://doi.org/10.4070/kcj.2019.0421. 624.
(13)
수치

관련 문서
In vitro cell migration in APE or JAG1 siRNA-treated M059K cell line Fig,11 Down-regulation of JAG1 induces S phase arrest in
(A) Before repair of RCT shows fatty degeneration of supraspinatus (stage 2) and infraspinatus (stage 3) muscle and atrophy (Grade 2) (B) After 18months later
Effect of cell migration of JMJD6 transcript variants over- expressed MCF-7 cells using Transwell.. Effect of cell migration of JMJD6 transcript variants
These results suggest that the bilobalide inhibits the cell proliferation and induces the apoptotic cell death in FaDu human pharyngeal squamous cell carcinoma via both
Objective: The purpose of this study is to use it as a basis for the health care guidelines for the muscle mass and cardiovascular disease of Korean population by
Taken together, this present study show that VSMC proliferation is blocked by CPP343 treatment, and this anti-proliferative effect is correlated with cell-cycle
They also showed significantly improved cell migration and proliferation with respect to negative control in the scratch - wound healing assay using confluent monolayers
Conclusion: In conclusion, the relaxant effects of neuromuscular blockers on the uterine smooth muscle may be transmitted via nicotinic acetylcholine receptors
