서 론
전 인구의 5-15%에 이환될 정도로 흔한 질환인 요석은 체외충격파쇄석술이나 내비뇨기과학적인 치료법 등 다양 한 치료법으로 치료가 가능하지만 재발이 흔하여1 예방이
필요하나 현재까지 효과적인 내과적 예방법이 개발되어 있 지 않아 효과적인 예방적 치료제의 개발이 필요한 실정이 다.
요석 형성에 있어서 수산의 중요성이 널리 알려지게 되 었는데 물리화학적으로 칼슘농도 변화보다 수산농도 변화 가 수산칼슘 결정체 형성에 훨씬 중요하며 실제로 수산농
대장균의 개발
Development of Recombinant Escherichia coli expressing oxc and frc Gene of Oxalobacter formigenes
Yong Hyun Park, Byong Chang Jeong, Cheol Kwak, Ji Eun Oh, Bong Sub Kim, Eui Chong Kim1, Hyeon Hoe Kim
From the Departments of Urology and1Laboratory Medicine, Seoul National Uni- versity College of Medicine and Clinical Research Institute, Seoul National Uni- versity Hospital, Seoul, Korea
Purpose: Oxalobacter formigenes(O. formigenes) is an obligate anaerobe, which may be important in the prevention of stone formation.O. formigenes degrades oxalates using oxalyl-CoA decarboxylase and formyl- CoA transferase encoded by theoxcandfrcgenes, respectively. Attempts were made to develop recombinant Escherichia coli(E. coli) expressing both the oxc and frc genes of O. formigenenes.
Materials and Methods: After the extraction of total RNA from O.
formigenes, a reverse transcriptase-polymerase chain reaction (RT-PCR) was carried out using primers synthesized according to the oxcand frcgenes reported in GenBank. The cloned cDNA encoding oxalyl-CoA decar- boxylase and formyl-CoA transferase was introduced into the pET-22b (+) plasmid vector. The constructs were verified by restriction analysis and DNA sequencing. The plasmid vector containing the cDNA fragment was transformed into competent E. coli BL21 (DE3). The recombinant E. coli was then analyzed using SD-SPAGE for the protein expressions of oxc and frcgene products, and visualized by staining with Coomassie Blue.
Results: Restriction enzyme and sequence analyses showed the gene cloned into the pET-22b (+) plasmid vector was identical to the reported oxcand frcgenes. After the transformation into the competentE. coli, the SDS-PAGE analysis showed the recombinantE. coliexpressed the proteins migrating at 66 and 50KD, which was identical to the reported weight of oxalyl-CoA decarboxylase and formyl-CoA transferase.
Conclusions: A recombinant E. coli, expressing oxc and frc genes, was successfully produced. Further studies may be necessary to investigate their enzymatic activities on the degradation of oxalate in the development of a new therapeutic strategy for the prevention of stone formation.
(Korean J Urol 2007;48:206-211)
Key Words: Oxalobacter formigenes, Escherichia coli, oxc and frc
대한비뇨기과학회지 제 48 권 제 2 호 2007
서울대학교 의과대학 비뇨기과학교실,
1검사의학교실
박용현 정병창 곽 철 오지은
김봉섭 김의종1 김현회
접수일자:2006년 10월 9일 채택일자:2006년 12월 22일
교신저자: 김현회
서울대학교병원 비뇨기과 서울시 종로구 연건동 28
110-744
TEL: 02-2072-2425 FAX: 02-742-4665 E-mail: [email protected] 본 논문은 2006년도 대한비뇨기과학회 추계학술대회에서 우수논문상을 수상한 논문임.
본 논문은 2005년도 서울대학교병원 일반 연구비(04-2005-037-0)의 보조로 이루 어진 논문임.
206
수산분해효소인 수산 보효소 A 탈카르복실라아제 (oxalyl CoA decarboxylase)와 포밀산 보효소 A 전이효소 (formyl CoA transferase)를 부호화하는 oxc와 frc 유전자의 발현이 장 내 수산을 분해하여 요중 수산 농도를 낮출 수 있다는 가정하에 oxc와 frc 유전자를 발현하는 재조합 대장균을 제 작하고자 하였다.
대상 및 방법 1. O. formigenes의 배양
장 내 세균인 O. formigenes 균주인 OxB (ATCC 35274)를 사용하였다. 배양 배지로 Allison 등3의 방법에 의하여 me- dium B broth (ATCC culture medium 1514)를 제조하여 사용 하였다. Medium B broth를 24시간 동안 혐기성 방 (Bactron Anaerobic/Environmental ChamberR)에서 배양한 후 O. for- migenes를 접종하였다.
2. O. formigenes로부터 RNA의 추출
O. formigenes를 medium B broth에서 이틀간 배양한 후 배 양액을 4oC에서 5분간 300xg로 원심분리하여 상층액을 덜 어내었다. 이때 세포수가 1x109을 넘지 않도록 하였다.
Lysozyme/TE buffer (400μg/ml) 100μl를 첨가하여 잘 혼합 하여 준 후 실온에서 3-5분간 배양하였다. RLT buffer 350μl 를 더하고 잘 혼합하여 준 후 최고 속도로 2분간 원심분리 한 후 상층액을 새 튜브로 옮기고 무수 알코올 250μl를 더 하였다. Rneasy mini column (QiagenR)으로 옮긴 후에 8,000 xg 이상에서 15초간 원심분리하고 RW1 buffer 700μl를 더 한 후 8,000xg 이상에서 15초간 원심분리하였다. 다시 RPE buffer 500μl를 더하고 8,000xg 이상에서 15초간 원심분리 후 다시 RPE buffer 500μl를 더한 후 8,000xg 이상에서 2분 간 원심분리하였다. Collection tube를 새 것으로 바꿔준 후에 RNase- free water 30-50μl를 더하고 1분간 원심분리하였다.
1.3Kbp): 5x buffer 5μl, dNTP 10mM, primer oxcF/frcF 1μl, oxcR/frcR 1μl, enzyme mix 1μl (OmniscriptTM reverse tran- scriptase, SensiscriptTM reverse transcriptase, HotStar TaqR DNA polymerase), template DNA 2μl, RNase-free water 14μl 반응액에서 50oC에서 30분, 95oC에서 15분간 반응시키고 94oC 1분, 54oC 1분, 72oC 1분씩 25 주기로 반응시킨 후 72oC 에서 10분간 반응시키고 전기영동을 통하여 확인하였다.
4. 발현벡터에 연결 삽입 후 제한효소 처리 및 염기서열 분석
역전사중합효소연쇄반응법을 이용하여 획득한 oxc와 frc cDNA fragment를 pET-22b(+) plasmid vector에 도입하여 oxc와 frc 발현벡터 (pCR-oxc & pCR-frc)를 제작하였다. Auto- mated DNA sequencer (ABI PrismR)를 이용해 삽입한 cDNA fragment의 염기서열을 확인하였다.
5. 대장균에 형질전환
발현벡터를 배지가 포함된 비병원성의 competent E. coli BL 21 (DE3) (NovaGenR)에 넣어주고 4oC에서 5분 동안 방치 한 후 30초간 42oC의 열충격 (heat shock)을 가하고 다시 2분 간 4oC에 방치시켜 형질전환하였다.
6. SDS-PAGE 분석
형질전환된 대장균을 isopropyl-β-D-thiogalactopyranoside (IPTG)가 1mM 포함된 Luria Bertani (LB) broth에서 37oC에서 4시간 동안 발현시키고, 단백질을 추출하여 10% SDS-PAGE 로 전기영동시킨 후 Coomassie Blue 염색법으로 확인하였다.
결 과
1. oxc와 frc 유전자의 cloning
O. formigenes로부터 RNA를 추출한 후 oxc 유전자와 frc
유전자의 coding sequence에 대한 primer를 제작 이용하여 역전사중합효소연쇄반응법을 이용하여 획득한 oxc와 frc의 cDNA 절편 (각각 1,704, 1,284bp)을 pET-22b(+) 벡터 (Fig.
3)에 삽입한 후 자동염기서열분석기를 이용하여 염기서열을 분석한 결과, 유전자 은행에 있는 자료와 본 실험에서 얻어 진 oxc와 frc 유전자의 염기서열은 동일하였다 (Fig. 1, 2).
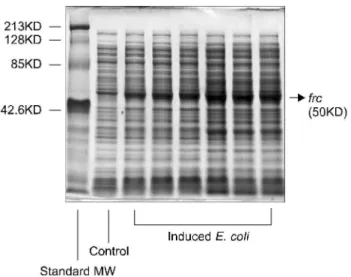
2. oxc와 frc 유전자 발현 재조합 대장균 획득 Competent E. coli에 상기 발현벡터를 형질전환시켜 개발 한 재조합 대장균의 SDS-PAGE 분석을 시행하고 Coomassie Blue 염색하였다. 그 결과 competent E. coli에 비해 각각 66, 50KD 크기의 band가 새로 관찰되었으며 이는 oxc와 frc 단 백의 크기와 동일하였다 (Fig. 4, 5).
고 찰
요로 결석은 가장 흔한 비뇨기과 질환 중의 하나로, 한국 인의 요로 결석의 평생 유병률은 남자는 6.0%, 여자는 1.8%
로 평균 3.5%에 이른다.4 요로 결석의 시점 유병률 또한 1998년 현재 남자는 1.9%, 여자는 0.3%로, 한국 전체 인구 의 0.9%가 요로 결석을 앓고 있다고 추정되며 이를 1998년 한국 인구로 환산할 때 국내에는 약 40만명의 요로 결석 환 자가 존재한다.4많은 요로 결석의 원인 중에서도 수산칼슘 석은 한국인의 요로 결석의 약 81.6%를 차지하고 있는 가장 흔한 결석의 원인이며,5추적 관찰하였을 때 1년에 약 10%, 5년에 35%, 10년이 지나면 50% 이상의 높은 재발률을 보이 고 있다.6
수산칼슘석에서 요중 수산칼슘석의 결정화에는 수산 농 도의 변화가 칼슘 농도의 변화보다 15배 이상 중요하다고 알려져 있으며,2요로 결석 환자에서 요중 수산 배설의 감소 가 새로운 요석 발생의 위험을 줄이는 가장 중요한 인자들 중의 하나로 평가받고 있다.7
정상 장 내 세균인 O. formigenes는 위장관 내에서 수산을 분해하여 에너지원으로 사용하는 그람 음성의 혐기성 세균3 으로, 위장관 내에서 수산을 분해하기 때문에 요석의 중요 한 성분인 수산의 위장관을 통한 흡수를 감소시킨다.8 O.
Fig. 1. The coding sequence of the oxc gene.
formigenes가 수산의 위장관을 통한 흡수를 조절할 수 있다 는 것은 매우 흥미로운 사실이지만, 그동안 O. formigenes에 대한 연구는 균주 배양의 어려움으로 인해 제한되어 있었 다. 그러나 O. formigenes가 수산의 이화작용에 필요한 수산- 보효소 A 탈카르복실라아제 (oxalyl-CoA decarboxylase)를 암호화하는 독특한 유전자인 oxc를 발현한다는 사실과 이 유전자의 cloning이 완성되어 염기 서열이 알려지게 되면서
균 검출에 많은 발전이 있었다.9 여러 O. formigenes 균주들 의 oxc 유전자 염기서열을 분석한 결과 5' end 부위에 독특 하면서도 높게 보존되어 있는 영역이 있음이 밝혀졌고, 이 를 통해서 속 특이적 (genus specific) PCR primer의 제작이 가능하게 되어 중합효소연쇄반응법을 통한 균 동정에 이용 하게 되었다.10,11
이러한 분자생물학적 방법을 이용하여 저자들은 국내 요 Fig. 2. The coding sequence of the frc gene.
Fig. 3. The structure of pET-22b (+) vector, including the T7 pro- motor (A) and the structure of the constructed oxc/frc expression vec- tor [named pET-22b(+) - oxc/frc]
(B).
석 환자의 약 37%, 정상인의 약 77%에서 O. formigenes가 기생하고 있음을 발견하여 정상인에 비하여 요석 환자군에 서 O. formigenes의 검출률이 유의하게 감소되어 있음을 확 인하였다.11또한 요석 환자들의 분변에서 DNA를 추출하여 oxc 유전자를 표적으로 정량 중합효소연쇄반응법을 시행한 결과 O. formigenes의 상대적 균수와 24시간 요중 수산치 간 에 역상관성이 있음을 밝혀 O. formigenes가 요석의 형성과 인과관계가 있음이 밝혀졌다.12 이러한 연구 결과들은 O.
formigenes를 이용한 예방적 치료제 개발이 뚜렷한 내과적 치료가 없는 요석 환자들의 재발 방지에 도움을 줄 수 있을 것으로 생각되는 과학적인 근거이다.
현재까지도 O. formigenes를 Probiotics로써 요석의 치료에 이용하려는 연구는 진행 중에 있다. Sidhu 등13은 과수산뇨 증을 유발시킨 Sprague-Dawley 백서를 대상으로 한 실험에 서 2주간 O. formigenes를 먹였을 때 섭취를 시작한지 2일 경과 후부터 혈중 수산의 농도가 O. formigenes 섭취량에 비 례하여 감소함을 확인하였다. 그러나 O. formigenes의 섭취 를 중단한지 5일 경과 후부터 실험 대상 백서의 분변에서 O. formigenes의 검출률이 감소하여 정량 중합효소연쇄반응 법으로 측정되지 않을 정도의 수준에 이른다는 사실도 함 께 확인되었다. 이것은 요석의 예방적 치료제로서 O. for- migenes의 가능성과 방법상의 한계를 함께 보여주는 결과 이다.
Kwak 등14은 O. formigenes를 위장관 내에 투여하기 위해 반드시 선행되어야 할 O. formigenes의 순수 분리 및 배양을 시도한 바 있다. 인체 분변을 대상으로 중합효소연쇄반응
에 양성 반응을 보인 집락들에 대하여 계대배양을 시행한 결과, 점차 중합효소연쇄반응이 음성으로 전환되다가 5차 계대배양 시에는 양성 반응을 보이는 집락들이 관찰되지 않음을 확인하였다. 따라서 O. formigenes의 순수 분리 및 동정은 현재까지 시행이 어려운 과정으로 생각하며, 이를 극복하기 위해서는 O. formigenes의 수산분해능력을 다른 방법으로 발현시키려는 시도가 필요하다.
O. formigenes는 에너지원으로 전적으로 수산만을 사용하 는데, 수산을 이화시키기 위하여 포밀산-보효소A 전이효소 (formyl-CoA transferase)와 수산-보효소A 탈카르복실라아제 (oxalyl-CoA decarboxylase)를 사용한다. O. formigenes는 oxlT 유전자에 의하여 부호화되는 oxlT라는 transporter에 의하여 수산과 포밀산의 교환을 통하여 수산을 세포 내로 받아들 인다.15 세포 내로 유입된 수산은 먼저 포밀산-보효소A 전 이효소에 의하여 활성화되고, 이후 수산-보효소A 탈카르복 실라아제에 의하여 이산화탄소와 포밀산으로 탈카르복실 화되고, 이로 인해 oxlT는 더 많은 수산을 세포 내로 유입할 수 있게 된다.
최근 수산 분해 관련 단백들을 조절하는 유전자로 수산- 보효소A 탈카르복실라아제를 부호화하는 oxc 유전자9와 포 밀산-보효소A 전이효소를 부호화하는 frc 유전자16가 밝혀 졌다. 저자들은 이를 이용하여 수산분해균주인 O. formi- genes의 주요 수산분해효소인 수산-보효소A 탈카르복실라 아제와 포밀산-보효소A 전이효소를 부호화하는 oxc와 frc 유전자를 발현하는 재조합 대장균을 개발하였다. 하지만 아직 oxc 유전자가 발현하는 수산-보효소A 탈카르복실라아 Fig. 4. SDS-PAGE analysis of the induced oxc gene recombinant
E. coli using Coomassie blue staining. The new band located at 66KD was shown after the induction of recombinant E. coli compared with the control (non-recombinant E. coli). Standard MW: standard molecular weight.
Fig. 5. SDS-PAGE analysis of the induced frc gene recombinant E. coli using Coomassie blue staining. The new band located at 50KD was shown after the induction of recombinant E. coli compared with the control (non-recombinant E. coli). Standard MW: standard molecular weight.
O. formigenes로부터 RNA를 추출한 후 oxc와 frc 유전자 의 coding sequence에 대한 primer를 제작 이용하여 역전사 중합효소연쇄반응법에 의하여 cDNA를 획득하여 발현벡터 에 삽입한 후 제한효소와 sequencing reaction을 통하여 염기 서열을 확인하였고, competent E. coli에 상기 발현벡터를 형 질전환시켜 기능 유전자를 발현할 수 있는 재조합 대장균 을 획득하는 데 성공하였다. 추후 추가적인 연구를 통하여 수산분해효소 발현 재조합 대장균이 수산분해능을 가짐을 확인하고, 요석 형성 동물모델에서 치료 효과를 확인할 수 있다면, 이를 이용한 치료가 요석 형성 및 요석 재발을 막을 수 있는 유용한 방법이라는 것을 확증할 수 있을 것이라 생각한다.
REFERENCES
1. Ahlstrand C, Tiselius HG. Recurrences during a 10-year fol- low-up after first renal stone episode. Urol Res 1990;18:397-9 2. Scheid CR, Koul H, Hill WA. Oxalate ion and calcium oxalate crystal interactions with renal epithelial cells. In: Coe FL, Favus MJ, Pak CY, Parks JH, Preminger GM, editors. Kidney stones: medical and surgical management. Philadelphia: Lip- pincott-Raven; 1996;129-43
3. Allison MJ, Dawson KA, Mayberry WR, Foss JG. Oxalobacter formigenes gen nov., sp. Nov.,: oxalate degrading anaerobes that inhibit the gastrointestinal tract. Arch Microbiol 1985;141:1-7 4. Kim HH, Jo MK, Kwak C, Park SK, Yoo KY, Kang DH, et
quence, and gene expression of the oxalyl-coenzyme A decar- boxylase gene, oxc, from the bacterium Oxalobacter formi- genes. J Bacteriol 1994;176:2468-72
10. Sidhu H, Holmes RP, Allison MJ, Peck AB. Direct quanti- fication of the enteric bacterium Oxalobacter formigenes in human fecal samples by quantitative competitive-template PCR. J Clin Microbiol 1999;37:1503-9
11. Kwak C, Jeong BC, Lee JH, Kim HK, Kim EC, Kim HH.
Molecular identification of Oxalobacter formigenes with the polymerase chain reaction in fresh or frozen fecal samples.
BJU Int 2001;88:627-32
12. Kwak C, Kim HK, Kim EC, Choi MS, Kim HH. Urinary oxalate levels and the enteric bacterium Oxalobacter formi- genes in patients with calcium oxalate urolithiasis. Eur Urol 2003;44:475-81
13. Sidhu H, Allison MJ, Chow JM, Clark A, Peck AB. Rapid reversal of hyperoxaluria in a rat model after probiotic ad- ministration of Oxalobacter formigenes. J Urol 2001;166:
1487-91
14. Kwak C, Oh JE, Kim HK, Choi MS, Kim EC, Kim HH.
Isolation and purification of the oxalate-degrading bacterium Oxalobacter formigenes in human feces. Korean J Urol 2003;44:785-90
15. Anantharam V, Allison MJ, Maloney PC. Oxalate: formate ex- change. The basis for energy coupling in Oxalobacter. J Biol Chem 1989;264:7244-50
16. Sidhu H, Ogden SD, Lung HY, Luttge BG, Baetz AL, Peck AB. DNA sequencing and expression of the formyl coenzyme A transferase gene, frc, from Oxalobacter formigenes. J Bac- teriol 1997;179:3378-81
![Fig. 3. The structure of pET-22b (+) vector, including the T7 pro-motor (A) and the structure of the constructed oxc/frc expression vec-tor [named pET-22b(+) - oxc/frc]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5244395.131868/4.892.85.535.154.721/structure-vector-including-motor-structure-constructed-expression-named.webp)