저작자표시-비영리-동일조건변경허락 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. l 이차적 저작물을 작성할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 동일조건변경허락. 귀하가 이 저작물을 개작, 변형 또는 가공했을 경우 에는, 이 저작물과 동일한 이용허락조건하에서만 배포할 수 있습니다.
이학 석사학위 논문
ERK1/
2에 의한 미토콘드리아의
형태조절 기전 규명
아 주 대 학 교
대 학 원
의생명과학과/
분자의학전공
김 영 화
ERK1/
2에 의한 미토콘드리아의
형태조절 기전 규명
지도교수
정 선 용
이 논문을 이학 석사학위 논문으로 제출함.
2011년
8월
아 주 대 학 교
대 학 원
의생명과학과/
분자의학전공
김 영 화
김영화의 이학 석사학위 논문을 인준함.
심사위원장
정
선
용
인
심 사 위 원
김
완
기
인
심 사 위 원
박
찬
배
인
아 주 대 학 교
대 학 원
2011년 06월 23일
- 국문요약 -
ERK1/2 에 의한 미토콘드리아의 형태조절 기전 규명
미토콘드리아는 세포 내 에너지 생산뿐만 아니라 세포의 삶과 죽음을 포함한 세포의 운명 결정과 직결된 다양한 생명활동에 관여하고 있으며, 미토콘드리아의 형태(morphology) 조절이 세포의 항상성(homeostasis) 유지에 필수적이라는 사실이 차츰 밝혀지고 있다. 미토콘드리아의 형태는 Mitofusin 1(Mfn1), Mitofusin 2(Mfn2), Optic atrophy protein 1(Opa1), Dynamin related protein 1(Drp1), Fission protein 1(Fis1) 등의 미토콘드리아 형태조절 단백질(mitochondria-shaping protein)이 관여하는 미토콘드리아 융합-분열 기구(mitochondrial fusion-fission machinery)에 의해 정교하게 조절되고 있다. 이러한 형태조절 단백질들이 어떻게 분자 수준에서 조절 되어 그들의 역할을 수행하는지에 관해서는 아직 명확하지 않다. 본 연구는 동일한 종류의 primary 배양세포에서 ERK1/2 의 활성화 정도에 비례하여 미토콘드리아가 짧아지는 현상의 발견으로부터 연구를 시작하게 되었다. ERK1/2 는 외부의 다양한 자극에 반응하여 세포성장, 증식, 분화 및 세포의 생(生)과 사(死)를 조절하는 세포 내 필수 kinase 로 최근의 연구에서 ERK1/2 가 미토콘드리아 외 막과 내막에 모두 국재(localization) 한다고 보고되었다. 이는 ERK1/2 에 의한 신호전달과 미토콘드리아 간에 직접적인 상관관계가 있음을 시사한다.따라서, 본 연구에서는 ERK1/2 의 활성화와 미토콘드리아 형태조절과의 상관관계 규명을 연구목표로 하였다. 우선, ERK1/2 활성을 저해 시켜서 미토콘드리아 형태에 미치는 영향을 관찰 하여 ERK1/2 의 활성화와 미토콘드리아 형태조절간의 연관성을 확인해 보았다. 여러 종류의 세포에 MEK/ERK 특이적 저해제인 PD98059 를 처리하여 ERK1/2 의 활성을 억제한 결과, 시험한 모든 세포에서 미토콘드리아가 길어지는 것을 확인 하였다. 이러한 ERK1/2 의 활성 저해에 의한 미토콘드리아의 형태변화가 미토콘드리아 형태조절 단백질과 직접적으로 연관되어 있는지 분석한 결과, 미토콘드리아 융합(fusion)조절에 관여하는 Mfn2 단백질의 특이적 증가를 확인하였다. 다음으로, ERK1/2 활성 저해에 의해 Mfn2 가 증가된 원인을 밝히기 위한 실험을 진행하였다. Mfn2 의 증가가 mRNA 수준에서 유전자 발현(expression)의 증가에 의한 것인지 RT-PCR 방법으로 확인하였고, 또한 Mfn2 의 증가가 단백질 수준의 안정성(stability) 증가에 의한 것인지 단백질 분해 경로에 관여하는 유비퀴틴화(ubiquitination) 분석을 통하여 확인하였다. 결과적으로 PD98059 처리에 의한 ERK1/2 의 활성 저해는 유비퀴틴화 감소로 인한 Mfn2 단백질 수준의 안정성을 증가시킨다는 사실을 확인하였다. Mfn2 의 유비퀴틴화 감소 원인을 확인 하기 위하여 지금까지 Mfn2 의 유비퀴틴화에 관여하는 것으로 보고된 2 종류의 E3 ubiquitin ligase 인 MARCH-V 와 Parkin 을 분석한 결과, PD98059 처리에 의한 ERK1/2 활성 저해는 MARCH-V 의 발현 감소를 유도 한다는 중요한 사실을 밝혔으며, 이는 MARCH-V 의 발현 감소가 Mfn2 의 유비퀴틴화 감소에 영향을 미쳤음을 시사한다.
결론적으로, ERK1/2 의 활성 변화는 E3 ubiqiutin ligase 인 MARCH-V 의 발현 량에 영향을 미치고, 이로 인해 미토콘드리아 형태조절 단백질 Mfn2 의 안정성의 변화를 가져와 결과적으로 미토콘드리아의 형태 변화를 초래한다는 사실을 밝혔다. 본 연구 결과는 세포의 항상성 유지에 필수적이라고 알려진 미토콘드리아의 형태조절이 ERK1/2 신호전달 경로에 의해 조절되고, 그 균형이 유지되고 있음을 시사한다. 본 연구는 미토콘드리아 형태조절에 의해 매개 되는 다양한 생명현상의 이해 증진에 기여 할 것으로 생각된다. 핵심어: 미토콘드리아, 미토콘드리아 형태, 미토콘드리아 융합/분열, ERK1/2, Mfn2, MARCH-V, 유비퀴틴화
차
례
국문요약 ··· ⅰ 차례 ··· ⅳ 그림차례 ··· ⅴ 표차례 ··· ⅶ . Ⅰ 서론 ··· 1 . Ⅱ 재료 및 방법 ··· 6 A. Cell culture ··· 6B. Reagent treatment and Western blot analysis··· 6
C. Ras activation assay ··· 7
D. Mitochondria fractionation ··· 7
E. Total RNA extraction and cDNA synthesis ··· 8
F. RT-PCR ··· 9 G. Immunoprecipitation analysis ··· 11 H. Confocal microscopy ··· 11 . Ⅲ 결과 ··· 13 . Ⅳ 고찰 ··· 29 . Ⅴ 결론 ··· 33 참고문헌 ··· 34 ABSTRACT ··· 36
그림
차례
Fig. 1. Various shapes of mitochondrial morphology ... 4
Fig. 2. Mitochondria-shaping proteins involved in mitochondrial fusion and fission
machinery………..………..5
Fig. 3. Mitochondrial morphology and endogenous ERK1/2 activation level in the three types of primary cultured cells ……….…...……….……….…... 14
Fig. 4. Mitochondrial morphology changes caused by inhibition of ERK1/2 activity in the Ras-activated primary cells………..………… …...……..…16
Fig. 5. Mitochondrial morphology changes caused by inhibition of ERK1/2 activity in the cell lines……...18
Fig. 6. Dose-dependent increase of Mfn2 level and mitochondrial elongation caused by inhibition of ERK1/2 activity in HT0180 cells ………..…...20
Fig. 7. Time-dependent increase of Mfn2 level and mitochondrial elongation caused by inhibition of ERK1/2 activity in HT0180 cells ………...………..22
Fig. 8. Effect of the cellular localization of Mfn2 by inhibition of ERK1/2 activity in HT01080 cells ………..…….24
Fig. 9. RT-PCR of the mitochondria-shaping genes by treatment of ERK1/2…………... 26 Fig. 10. Decrease of the MARCH-V-mediated ubiquitination of Mfn2 by inhibition of
표
차례
Table. 1. Primer sequences used for amplification of the mitochondria-shaping genes ………...……….…….9
I. 서 론
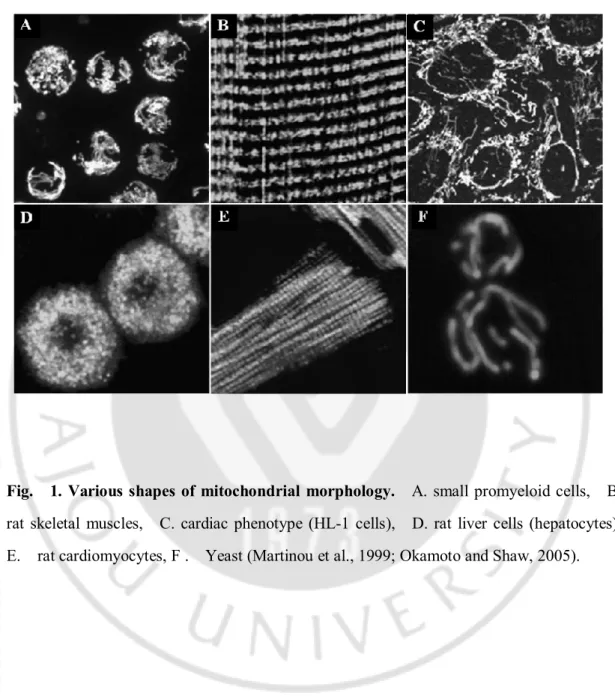
미토콘드리아는 이중 지질의 막과 그 속의 바탕 질(matrix) 등으로 구성되어 있으며 고유의 유전체(genome)를 갖고 있다. 미토콘드리아는 세포의 종류에 따라 작은 공(sphere) 모양이나 짧은 막대(rod) 모양에서부터 긴 관(tubule) 모양까지 그 형태는 매우 다양하다 (Fig. 1)(Kuznetsov et al., 2009). 미토콘드리아는 microtubules 과 microfilaments 를 따라 끊임없이 움직이며, 융합(fusion)과
분열(fission)을 통해 자신의 수와 형태를 조절한다. 미토콘드리아의
형태(morphology)는 세포 내 에너지 생산 외에도 증식(proliferation), 분화(differentiation), 사멸(apoptosis), 노화(aging), 발생(development), reactive oxygen species(ROS) 대사 등의 다양한 세포기능과 밀접한 관계가 있다는 사실이 차츰 밝혀지고 있다(Amati-Bonneau et al., 2009; Grandemange et al., 2009). 특히, 세포 사멸이 일어나는 동안 미토콘드리아 형태는 단편화(fragmentation) 되어 있으며, 인위적으로 형태조절 단백질을 억제시키면 사멸이 일어나거나 세포사멸 신호에 대해 감수성(sensitivity)의 차이를 보인다고 알려져 있다(Martinou et al., 1999).
이러한 미토콘드리아 형태는 미토콘드리아 형태조절 단백질(mitochondria-shaping protein)이 관여하는 미토콘드리아 fusion-fission machinery 에 의해 정교하게
조절되며, 미토콘드리아의 융합(fusion)에 관여하는 단백질로서 Mitofusin 1(Mfn1),
Mitofusin 2(Mfn2), Optic atrophy protein 1(Opa1)이 밝혀졌으며, 분열(fission)에
관여하는 단백질로서 dynamin related protein 1(Drp1)과 fission protein 1(Fis1)이 밝혀졌다(Anesti and Scorrano, 2006; Chan, 2006a). 미토콘드리아 형태조절 단백질 중 Mfn1, Mfn2, Opa1, Drp1 은 large GTPase domain 을 가지고 있고, Fis1 은 단백질
결합과 관련된 3 개의 tetratrico-peptide repeat(TPR) motif 를 가지고 있다(Fig. 2)(Cheung et al., 2007; Hom and Sheu, 2009). 미토콘드리아 형태조절 단백질들의 작용 기전에 대한 가설로 미토콘드리아 융합의 경우, 이웃한 미토콘드리아의 외 막에 있는 Mfn1, Mfn2 의 coiled-coil domain 이 서로 homo 또는 hetero 형태로
결합함으로써docking 이 일어나 N-terminal GTPase 활성에 의해 외 막이 융합되며,
내막에 있는 Opa1 이 내막의 융합에 관여한다는 가설이 있다. 분열의 경우는 미토콘드리아의 외 막에 있는 Fis1 의 TPR motif 에 large GTPase domain 을 가진 Drp1 이 결합하여 미토콘드리아 절단부위의 foci 에서 미토콘드리아를 두 개로 절단한다는 가설이 있다. 하지만, Fis1 은 Drp1 의 막 이동에 관련이 없다는 보고가 있고, 각각 다른 막에 존재하는 Mfn1, Mfn2 와 Opa1 을 연결하는 단백질이 발견되지 않아 아직 정확한 기전은 밝혀지지 않았다(Grandemange et al., 2009; Castanier and Arnoult, 2010). 미토콘드리아 형태조절 유전자에 돌연변이(mutation)가 발생하면 미토콘드리아의 융합과 분열을 반복하는 형태조절의 균형이 깨지게 되어 결과적으로 neurodegenerative diseases 나 제 2 형 당뇨와 같은 다양한 질환이 유발된다고 보고되어 있다(Detmer and Chan, 2007; Wang et al., 2009).
이렇듯 미토콘드리아 형태조절 단백질들의 역할과 기능에 대해서는 많은 연구가 이루어지고 있으나, 미토콘드리아 형태조절 단백질들이 세포 내 어떠한 신호전달 경로에 의해 조절되어 미토콘드리아 형태를 조절하는지에 대한 연구는 그 중요성에 비하여 매우 부족한 실정이다. 본 연구의 목표는 미토콘드리아 형태조절 단백질들이 세포 내 어떠한 신호전달 경로에 의해 조절되는지 그 분자 기전을 규명함으로써 세포의 항상성 유지에 필수적인 미토콘드리아 형태조절이 어떻게 균형을 유지하는지를 밝히는
것이다. 본 연구에서는 신경섬유종증 환자로부터 분리한 동일한 종류의 세가지 Ras 활성화 primary 세포를 비교 분석한 결과, 세포의 성장, 증식, 분화 및 세포의 생(生)과 사(死)를 조절하는 세포 내 대표적 신호전달 kinase 인 ERK1/2 의 활성화 정도에 비례하여 미토콘드리아 형태가 짧아져 있음을 발견하였고, ERK1/2 활성화와 미토콘드리아 형태조절 과의 상관관계를 알아보기 위해 MEK/ERK 특이적 저해제인 PD98059 를 처리하여 ERK1/2 의 활성 저해가 미토콘드리아 형태조절 단백질들의 발현 및 미토콘드리아 형태에 미치는 영향을 관찰하였다.
Fig. 1. Various shapes of mitochondrial morphology. A. small promyeloid cells, B.
rat skeletal muscles, C. cardiac phenotype (HL-1 cells), D. rat liver cells (hepatocytes), E. rat cardiomyocytes, F . Yeast (Martinou et al., 1999; Okamoto and Shaw, 2005).
A.
B.
Fig. 2. Mitochondria-shaping proteins involved in mitochondrial fusion and fission machinery. A. Mitochondrial fusion is regulated by Mfn1, Mfn2 and Opa1 and
mitochondrial fission is regulated by Drp1 and Fis1. B. Structure of proteins involved mitochondrial fusion and fission machinery (Chan, 2006b; Hom and Sheu, 2009).
.
Ⅱ 재료
및 방법
A. Cell culture
HT1080 (human epithelial fibro sarcoma), T24 (human epithelial bladder carcinoma) 를 1% antibiotics(GIBCO BRL, U.S.A), 10% FBS(Sigma Aldrich INC., U.S.A) 를 첨가한 RPMI 1640 medium(Hyclone Laboratories INC., U.S.A)에서 37 , 5% CO2 ℃ 조건으로 배양하였다. 신경섬유종증 환자로부터 분리한 Ras 가 활성화된 3 종류의 세포(normal, benign, malignant)를 1% antibiotics(Gibco BRL, U.S.A.), 15% FBS(Hyclone Laboratories INC., U.S.A.)를 첨가한 Dulbecco's modified Eagle's medium(Hyclone Laboratories INC, U.S.A.)에 배양하였다.
B. Reagent treatment and Western blot analysis
세포를 동일하게 무 혈청 배양액에 1 시간 starvation 시킨 뒤 MEK/ERK inhibitor 인 PD98059 (Calbiochem) 를 24 시간 동안 처리한 후 harvest 하여 Bio-rad Lowry protein assay reagent kit(Bio-rad laboratories, Hercules, CA, U.S.A.)방법으로 정량 분석하여 SDS-PAGE(Sodium dodecyl sulfate polyacrylamidegel electrophoresis) loading buffer(5X)와 섞은 후, 100℃에서 10 분간 가열하였다. 이를 각각 20-50 ㎍씩 loading 한 후 8-15% SDS-PAGE gel 에 전기영동 하여 분리하고, Polyvinylidene fluoride (PVDF, Immobilon-P membrane (0,45 ㎛), Millipore Corporation, U.S.A.)에 흡착 이동시켜, 다음과 같이 각각의 특이항체로 Western blotting 을 통해 발현 량을 확인하였다. 1 차 항체로는 ERK, phospho-ERK(Thr202/Tyr204), pJNK, pAKT, Parkin,
Total Ras 1:1000(cell signaling, Thecnology, Inc. Beverly, MA., USA.), Mitofusin1, OPA1, DLP1 1:1000(BD Transduction biotechnology, Inc.), Mitofusin2, Fis1 1:1000(Epitomics Inc.), MARCH5 1:500(Abcam, Chicago, IL., U.S.A.) 2 차 항체로는 peroxidase 가 conjugation 된 goat anti-rabbit antibody, goat anti-mouse antibody 1:5000 (Santa cruz Biotechnology, INC, U.S.A.)를 이용하여 반응시킨 뒤 WEST-ZOL Plus(Intron Biotechnology, Korea)를 처리하여 필름에 현상하였다.
C. Ras activation assay
세포 내 활성화된 Ras 형태인 GTP-Ras의 수준을 알아보기 위해 Ras activation assay kit(Upstate Biotech, NY)의 실험 방법을 이용하여 확인하였다. Assay는 cell lysate 로부터 Ras-GTP를 분리하기 위하여 affinity precipitation을 이용하였고, ell lysate는 Raf-1 RBD fusion 단백질을 결합한 agarose와 함께 incubation 하였다. Agarose bead는 원심 분리하여 모은 후 lysis buffer로 washing 하였다. 최종적으로 sample buffer를 첨가하여 95℃에서 5분간 가열 후 Western blot analysis를 시행하였 다.
D. Mitochondria fractionation
세포 내 미토콘드리아를 분리하기 위하여 PBS 로 배양액을 완전히
제거하고 PBS 2ml 을 넣어 세포를 회수하여 4℃로 맞춘 Micro centrifuge 에서 1200rpm 2 분 spin down 시키고 다시 PBS 를 넣어 pipeting 한 뒤 동일한 조건에서 spin down 하여 모아진 pellet 에 10mM HEPES(pH 7.4), 150mM NaCl, Protease inhibitor cocktail(Sigma Aldrich Co.), NaF(sodium fluoride, Sigma Aldrich Co.), Na3VO4
(sodium van date, Sigma Aldrich Co.)를 혼합하여 ice 에서 30 분간 반응 뒤 세포를 모아 Homogenizer 로 약 200 회 dunce 한다. 4℃로 맞춘 Micro centrifuge 에서 3000rpm(800g)에서 5 분 spin down 하여 상층 액만 회수하여 (pellet; unbroken of cell nucleus) 6000rpm(3000g) 에서 10 분 동안 spin down 하여 상층 액은 cytosol 로써 회수하여 사용하고 pellet 은 미토콘드리아로 protein lysis buffer 를 사용하여 lysis 한 뒤 Bradford protein assay(Bio-rad laboratories, Hercules, CA, U.S.A.)방법으로 정량 분석하여 SDS-PAGE(Sodium dodecyl sulfate polyacrylamidegel electrophoresis) loading buffer(5X)와 섞은 후, 100℃에서 10 분간 가열하였다.
E. Total RNA extraction and cDNA synthesis
배양한 세포를 Trizol™ reagent(Invitrogen Co., U.S.A.)를 이용하여 RNA 를 추출하고 그 중, RNA 3 ㎍을 이용하여 DNase I(Invitrogen Co., U.S.A.)3U 을 처리하여 남아있는 gDNA 를 제거한 후 cDNA 를 합성하였고, 합성한 cDNA 을 이용하여 PCR 로 증폭하여 확인하였다. 사용한 primer 는 RNA 가 PCR 의 주형이 될 수 없으며, intron 영역은 gDNA 에만 존재하는 특성을 이용하여 제작하였다. PCR 결과 gDNA 가 완전히 제거된 것으로 확인이 된 RNA 를 ReverAid™ First Strand cDNA Synthesis Kit(Fermentas)를 이용하여 cDNA 를 합성하였으며 이용하여 cDNA 를 합성하였으며, cDNA 합성 여부를 확인하기 위하여 GAPDH cDNA 에서
두 개의 exon 이 만나는 부분에 특이적인 primer set (F,
5´-GCCTCAAGATCATCAGCAATGCCT-3´; R, 5´-AGACCACCTGGTGCTCAGTGTAG-3´)를 제작하였고, cDNA 주형으로 하여 PCR 로 증폭하여 확인하였다.
F. RT-PCR
세포에서 추출한 total RNA 에서 합성된 cDNA 합성 여부를 확인하기 위하여 GAPDH cDNA 에서 두 개의 exon 이 만나는 부분에 특이적인 primer set(Table. 1)를 제작하였고, cDNA 를 주형으로 하여 유전자 특이 primer 를 이용하여 95 2℃ 분, 95℃ 30 초, 60℃ 40 초, 72 ℃ 40 초 에서 28 cycle 반복하여 PCR(MJ Research Co., U.S.A.)로 증폭된 산물을 1.5% agarose gel 에 전기영동 하여 EtBr 로 염색하여 transiluminator 로 확인하였다.
Table 1. Primer sequences used for amplification of the mitochondria-shaping genes.
Gene Primer sequences
GAPDH
F, 5’- GCC TCA AGA TCA TCA GCA ATG CCT-3’ R, 5’- AGA CCA CCT GGT GCT CAG TGT AG-3’ OPA1
F, 5’- GCT GCG CAG ACC ATG AAT TCA GGA AC-3’ R, 5’- GCT GTC CTC CGC AAA GTC ATT CCA C-3’ MFN1
F, 5’- GTG CTGGCT AAG AAGGCG ATT ACT GCA -3’ R, 5’- GAT AGC ACC TCA CCA ATG ATG GAA AGC T -3’ MFN2
F, 5’- GAA TGC ATC CCC ACT TAA GCA CTT TGT C -3’ R, 5’- CCA GAG GGC AGA ACT TTG TCC CAG AGC -3’ DLP1
F, 5’- AGG TCA TGC CGT GAA CCT GCT AGA T-3’ R, 5’- GCC TAC TAG CTC ACT CTG AAG AGT GTC-3’ FIS1
F, 5’- CCG TAA AGG CAT CGT GCTGCT CGA G-3’ R, 5’- TGT TCT GGG GCT CTG TCT GCA GCA AC-3’
G. Immunoprecipitation analysis
PD98059 를 처리한 세포에 RIPA buffer(Sigma Aldrich Co., U.S.A.)와 PMSF (Phenylmethanesulfonyl fluoride, Sigma Aldrich Co.), Protease inhibitor cocktail(Sigma Aldrich Co.), NaF(sodium fluoride, Sigma Aldrich Co.), Na3VO4 (sodium vanadate, Sigma Aldrich Co.)를 혼합하여 ice 에서 30 분간 반응 한 후, 세포를 모아 4 ℃ 에서 30 분 incubation 한다. 4℃로 맞춘 Micro centrifuge 에서 12000g 10 분 spin down 시키고 얻어진 시료를 Protein A/G PLUS Agarose beads(Santa Cruz Biotechnology, INC.) 20 ㎕ 넣고 4℃에서 1 시간 반응시켜 pre-clearing 한다. 1 시간 뒤 4℃ Micro centrifuge 에서 12000g 10 분 spin down 후 상층 액만 옮겨 Bio-rad Lowry protein assay reagent kit(Bio-rad laboratories, Hercules, CA, U.S.A.)방법으로 정량 분석하여 500 ㎍의 단백질을 cell lysis buffer 로 total 500 ㎕을 맞추고 Mitofusin2 antibody (1:100 Epitomics Inc)를 넣어 4℃에서 2 시간 정도 반응 한 뒤 Agarose beads 20 ㎕ 넣고 4℃에서 over night 반응한다. 4℃로 맞춘 Micro centrifuge 에서 2600g 5 분 spin down 시키고 pellet 을 500 ㎕의 cell lysis buffer 로 5 번 washing 한다. pellet 에 20 ㎕의 2X SDS loading buffer 를 넣고 잘 섞어 준 뒤 100℃에서 5 분간 가열하고 Micro centrifuge 에서 2600g 5 분 spin down 시켜 이를 각각 loading 한 후 PVDF membrane(Millipore Corporation, U.S.A.)에 흡착 이동시켜 다음의 항체를 이용하여 Western blot analysis 를 시행하여 분석하였다.
H. Confocal microscopy
미토콘드리아 형태관찰 및 Mfn2의 세포 내 위치(cellular localization)를 확인하기 위하여, Poly-D-lysin(Sigma Aldrich, U.S.A.)코팅한 cover slip에 세포를
(4X105) 키운 뒤, 무 혈청(Serum free) 배양액에 1시간 동안 starvation하고, PD98059를 24 시간 동안 처리하였다. 공 초점 현미경으로 관찰하기 위해, 배양액에 250nM MitoTracker(Molecular Probes™, U.S.A.)를 넣어 20분간 처리하여 미토콘드리아를 염색한 뒤 PBS로 10분씩 3회 세척하고 MeOH:Acetone을 1:1로 섞어 -20 ℃에서 5분간 처리하여 세포고정(fixation)을 하였다. 세포 내 위치를 확인을 위해 1% BSA(bovine serum albumin)로 상온에서 1시간 동안 blocking 한 뒤, 일차 항체를 1% BSA가 녹아있는 PBS에 희석하여 4℃에서 12시간 이상 반응시키고 PBS로 씻어준 후 FITC(fluorescein isothiocynate-Cappel ICN Pharmaceuticals, Aurora, OH)가 결합되어 있는 2차 항체를 1시간 반응시킨 다음 관찰하였으며, 미토콘드리아 형태관찰을 위해 세포 고정 후 0.1M Glycine으로 실온에서 3분씩 3회, 0.1% Triton X-100 실온에서 10분씩 3회 처리하여 permeabilization 한다. PBS로 10분간 세척 후 Vectashield in DAPI(Vector Laboratories Co., U.S.A)로 mounting과 동시에 핵을 염색하여 샘플을 완성하였다. 이 후 공 초점 현미경(Zeiss LSM710 confocal laser scanning microscope; CLSM, Germany)을 사용하여 40x weter, 60x oil lens 조건으로 관찰하였다.
III. 결 과
A. Ras 활성화 primary 세포에서의 ERK1/2 활성화와 미토콘드리아 형태
상관관계
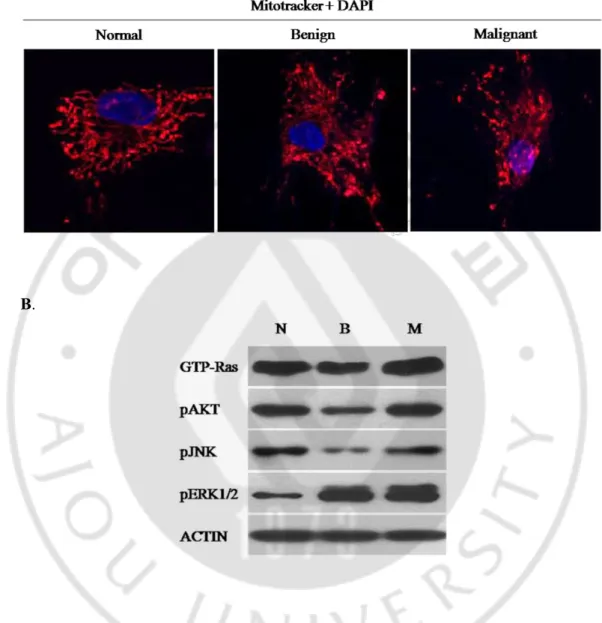
신경섬유종증 환자에서 분리 배양한 Ras 가 활성화된 3 종류의 primary 세포 (normal, benign, malignant)에서 세포 내 ERK1/2 의 활성이 증가 될수록 미토콘드리아의 형태가 짧아져 있다는 사실을 발견하였다(Fig. 3). 이 결과로부터 ERK1/2 의 활성화와 미토콘드리아 형태조절과의 상관관계를 밝히기 위한 연구를 진행하였다.
A.
B.
Fig. 3. Mitochondrial morphology and endogenous ERK1/2 activation level in the
three types of primary cultured cells. A. Confocal microscopy for mitochondrial
morphology in the three ras-activated cells (normal, benign and malignant) stained with mitotracker (red). The mitochondria were gradually fragmented from normal to malignant primary cells. B. Endogenous levels of pERK1/2 were gradually increased from normal to the malignant cells.
B. Primary 세포에서 ERK1/2 활성화 저해에 의한 미토콘드리아 형태 변화
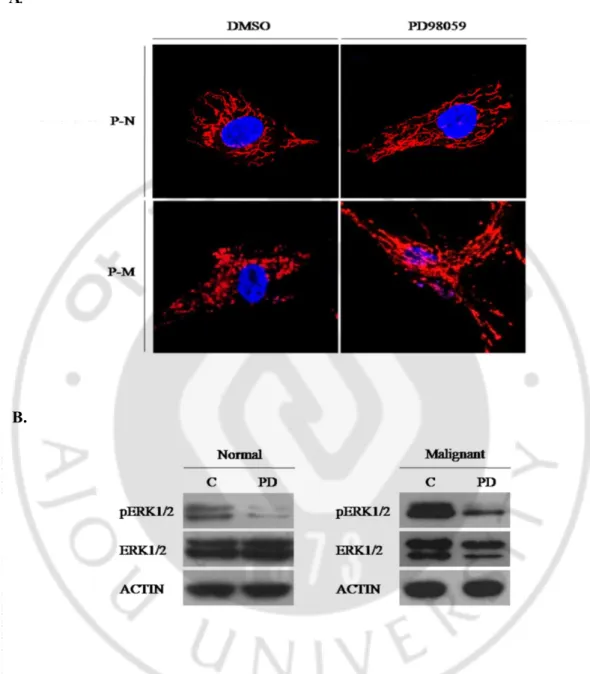
ERK1/2 활성화 정도와 미토콘드리아 형태의 차이가 확실한 두 종류의 primary 세포(normal, malignant)에 MEK/ERK 저해제인 PD98059 80μM 를 처리하여 ERK1/2 활성화를 억제한 결과, ERK1/2 활성화의 감소에 따라 미토콘드리아의 형태가 길어지는 것을 관찰하였다(Fig. 4).
A.
B.
Fig. 4. Mitochondrial morphology change caused by inhibition of ERK1/2 activity in the Ras-activated primary cells. A. Confocal microscopy for mitochondrial
morphology in the primary normal and malignant cells after treatment of 80μM of PD98059(P-N; primary normal cell, P-M; primary malignant cell). B. Western blot analysis of ERK1/2, pERK1/2 and ACTIN in the normal and malignant cells. pERK1/2 was decreased after treatment of PD98059.
C. 세포 주에서 ERK1/2 활성화 저해에 의한 미토콘드리아 형태 변화
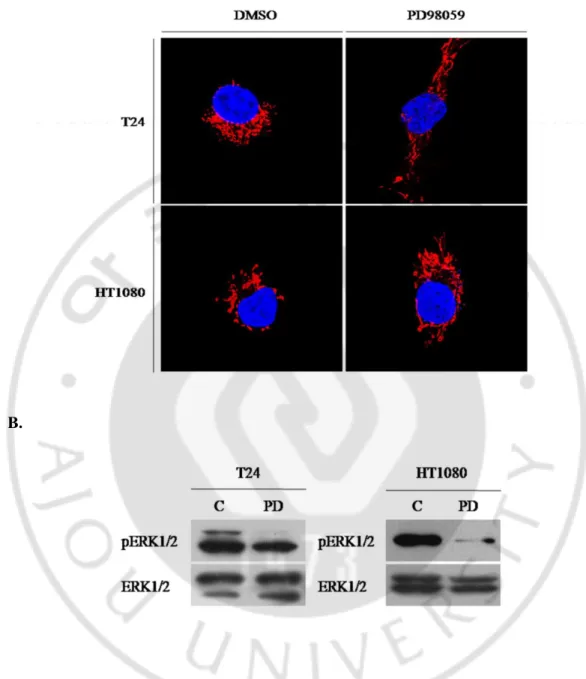
다른 세포에서도 ERK1/2 활성 저해에 따른 미토콘드리아 형태 변화를 확인 하기 위하여, HT1080 과 T24 세포 주에 PD98059 60μM 을 24 시간 동안 처리하여 ERK1/2 활성을 저해 하였다. 시험한 두 세포 주 모두 ERK1/2 의 활성 저해에 의해 미토콘드리아의 형태가 길어지는 것을 관찰할 수 있었다(Fig. 5).A.
B.
Fig. 5. Mitochondrial morphology change caused by inhibition of ERK1/2 activity in the cell lines. A. Confocal microscopy for mitochondrial morphology in the cell lines
after treatment of 60μM of PD98059(left; T24 cell, right; HT1080 cell). B. Western blot analysis of ERK1/2, pERK1/2 in both cells. pERK1/2 was decreased after treatment of PD98059.
D. ERK1/2 활성의 농도 별 저해에 의한 미토콘드리아 형태변화와
Mfn2 단백질 량 변화
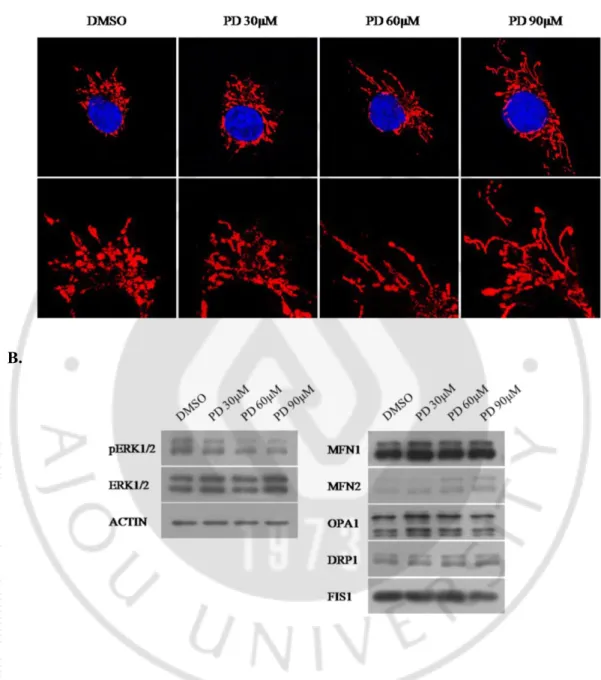
HT1080 세포 주에 PD98059 를 이용하여 농도 별로 ERK1/2 활성을 저해 하였더니 PD98059 농도가 증가함에 따라 미토콘드리아의 형태가 길어지는 것을 관찰할 수 있었다. 이때 다른 미토콘드리아 형태조절 단백질(Mfn1, Opa1, Drp1, Fis1)의 양적 변화는 관찰되지 않았고, Mfn2 의 단백질 량이 증가된 것을 확인 하였다(Fig. 5).A.
B.
Fig. 6. Dose-dependent increase of Mfn2 level and mitochondrial elongation caused by inhibition of ERK1/2 activity in HT0180 cells. A. Confocal microscopy
for mitochondrial morphology in HT1080 cells. PD98059 treatment induced mitochondrial elongation in a dose-dependent manner. B. Western blot analysis of mitochondrial shaping-proteins, ERK1/2, pERK1/2 and ACTIN. The PD98059 treatment induced a specific increase of Mfn2 level in a dose-dependent manner.
E. 시간 별 ERK1/2 저해에 의한 미토콘드리아 형태 변화와 Mfn2 단백질 량
변화
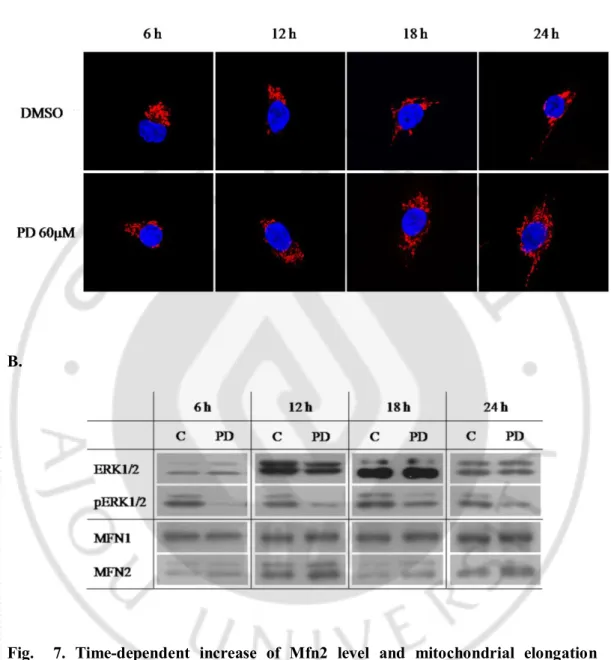
HT1080 세포 주에 PD98059 를 60μM 처리하여 6, 12, 18, 24 시간 동안 ERK1/2 activity 를 저해 하였더니 시간이 지남에 따라 미토콘드리아의 형태가 더 길어지는 것을 관찰할 수 있었고, 또한 미토콘드리아 형태조절 Mfn2 의 단백질 량도 비례하게 증가되는 것을 확인하였다(Fig. 6).
A.
B.
Fig. 7. Time-dependent increase of Mfn2 level and mitochondrial elongation caused by inhibition of ERK1/2 activity in HT0180 cells. A. Confocal microscopy
for mitochondrial morphology in HT1080 cells. PD98059 treatment induced mitochondrial elongation in a time-dependent manner. B. Western blot analysis of mitochondrial shaping-proteins, ERK1/2, pERK1/2 and ACTIN. The PD98059 treatment induced a specific increase of Mfn2 level in a time-dependent manner.
F. ERK1/2 의 활성 저해에 따른 Mfn2 의 세포 내 국재
PD98059 에 의한 ERK1/2 활성 저해에 따른 Mfn2 의 세포 내 국재(localization)의 변화 여부를 관찰하였다. PD98059 를 처리한 세포로 부터 cytosol 과 미토콘드리아를 분획하여 Western blot analysis 를 실시한 결과(Fig. 9A)와 PD98059 를 처리한 세포에 Mfn2 항체를 이용하여 면역 세포 화학 방법으로 Mfn2 단백질을 염색하여 공 초첨 현미경으로 관찰한 결과(Fig. 9B)로 부터 PD98059 처리에 의한 Mfn2 단백질의 세포 내 국재는 변화가 없음을 확인하였다.
A.
B.
Fig. 8. Cellular localization of Mfn2 on the mitochondria after inhibition of ERK1/2 activity. A. Mfn2 was localized in the mitochondria fraction but not in the cytosol fraction
after treatment of PD98059(C; cytosol, M; mitochondria). B. Confocal microscopy for Mfn2 localization after treatment of PD98059 in HT1080 cells by immunocytochemistry with anti-Mfn2 antibody.
G. PD98059 처리에 따른 MFN2 mRNA 발현 량
PD98059 처리에 의한 Mfn2 단백질 량의 변화가 전사(transcription) 수준에서 Mfn2 유전자의 mRNA 발현 량의 변화에 기인하는지 규명하기 위하여 PD98059 를 60μM 의 농도에서 24 시간 동안 ERK1/2 활성을 억제한 뒤 RNA 를 추출하여 각각의 특이적 primer 를 이용하여 RT-PCR 을 수행하였다. MFN2 를 비롯한 미토콘드리아 형태조절 유전자 모두 control 과 비교 하였을 때 mRNA level 의 차이가 없음을 확인하였다(Fig. 7).
Fig. 9. RT-PCR of the mitochondria-shaping genes by treatment of ERK1/2. RT-PCR
analysis result showed that mRNA expression level of MFN2 gene was not changed after treatment of PD98059.
H. PD98059 처리에 따른 Mfn2 단백질 안정성
PD98059 처리에 의한 Mfn2 단백질량의 변화가 Mfn2 단백질의 안정성(stability) 변화에 기인한 것인지 확인하기 위해 단백질 분해 경로에 관여하는 유비퀴틴화(ubiquitination)를 분석하였다. 우선, PD98059 처리에 의해 Mfn2
단백질의 유비퀴틴화가 감소되어 단백질의 안정성이 증가 된 것을
확인하였다(Fig. 10A). 자세한 기전을 확인하기 위하여 Mfn2 의 E3 ubiquitin ligase 인 MARCH-V 와 Parkin 을 분석한 결과 PD98059 처리에 의한 ERK1/2 저해는 MARCH-V 의 발현 감소를 유도한다는 사실을 밝혔다(Fig. 10B).
A.
B.
Fig. 10. Decrease of the MARCH-V-mediated ubiquitination of Mfn2 by inhibition of ERK1/2 activity. A. Immunoprecipitation analysis of ubiquitinated Mfn 1 and Mfn2
proteins in HT1080 cells. Ubiquitination level of Mfn2 was significantly decreased by PD98059 treatment. B. Western blot analysis of the E3 ubiquitin ligases, MARCH-V and Parkin, in HT1080 cells after treatment of PD98059.
IV. 고 찰
미토콘드리아는 세포 내 에너지 생산뿐만 아니라 세포의 삶과 죽음을 포함한 세포의 운명 결정과 직결된 다양한 생명활동에 관여하고 있으며, 미토콘드리아의 형태(morphology) 조절이 세포의 항상성(homeostasis) 유지에 필수적이라는 사실이 밝혀지고 있다. 미토콘드리아 형태조절은 미토콘드리아 형태조절 단백질(mitochondria-shaping protein)이 관여하는 미토콘드리아 fusion-fission machinery 에 의해 정교하게 조절되며, 이러한 미토콘드리아 형태조절 단백질들의 역할과 작용기전 대해서는 많은 선행연구가 이루어져 있으나 미토콘드리아 형태조절 단백질들이 세포 내 어떠한 신호전달 경로에 의해 조절되어 미토콘드리아를 융합(fusion) 또는 분열(fission) 시키는지에 관한 연구는 그 중요성에 비하여 부족한 실정이다. 본 연구의 필요성은 미토콘드리아 형태조절 단백질들을 조절하는 분자 기전을 규명 함으로서, 미토콘드리아 형태가 어떠한 기전으로 균형을 유지하며 나아가서, 미토콘드리아 형태조절에 의해 매개 되는 다양한 생명현상의 이해를 증진 시킬 수 있을 것이라 기대한다. 본 연구는 신경섬유종증 환자로부터 분리한 동일한 종류의 primary 배양세포에서 ERK1/2 의 활성화 정도에 비례하여 미토콘드리아가 짧아진다는 사실의 발견으로부터 연구를 시작하게 되었다. ERK1/2 는 외부의 다양한 자극에 반응하여 세포성장, 증식, 분화 및 사멸을 조절하는 세포 내 필수 kinase 로 최근 연구에서 ERK1/2 가 미토콘드리아 외 막과 내막에 모두 위치한다고 보고되었다.
이는 ERK1/2 에 의한 신호전달과 미토콘드리아 간에 직접적인 상관관계가 있음을 시사한다. 본 연구에서 ERK1/2 의 활성화와 미토콘드리아 형태조절과의 상관관계를 밝힘으로써 미토콘드리아 형태조절 단백질들이 세포 내 어떠한 신호전달 경로에 의해 조절되는지 그 분자 기전을 규명하고, 세포의 항상성에 중요한 미토콘드리아 형태조절이 어떻게 균형을 유지하는지 밝히고자 하였다. 우선, ERK1/2 의 활성화에 비례하여 미토콘드리아 길이가 짧아지는 현상에 기인하여 그 와 반대로 MEK/ERK 특이적 저해제인 PD98059 를 처리하여 ERK1/2 활성을 저해 시키고 미토콘드리아 형태에 미치는 영향을 관찰 한 결과 미토콘드리아의 형태가 길어지는 것을 확인 하였다. 신경섬유종증 환자로부터 분리한 primary 배양세포의 특이성과 primary 세포배양의 한계점을 보완하기 위하여 T24 와 HT1080 세포 주를 사용하여 primary 세포와 동일한 실험을 적용해 본 결과 ERK1/2 활성 저해에 따라 미토콘드리아의 길이가 길어지는 것을 확인 하였다. 이와 같은 결과를 통해 ERK1/2 의 활성화와 미토콘드리아 형태조절 간에 밀접한 연관성이 존재함을 확인 하였고, 이후 실험은 primary 세포와 T24 세포 주에 비하여 native 한 미토콘드리아 길이 짧고 그 수가 적당하여 ERK1/2 활성 저해에 따라 길어지는 미토콘드리아 형태관찰에 적합한 HT1080 세포 주를 사용하였다. 먼저, PD98059 를 농도 별로 처리 한 결과 농도 의존적으로 미토콘드리아 길이가 길어짐을 확인하였는데, 이는 ERK1/2 활성화 정도에 따라 미토콘드리아 길이가 조절된다는 것을 의미하고, 이러한 미토콘드리아의 형태변화가 미토콘드리아 형태조절 단백질과 관련되어 있는지 분석한 결과, 미토콘드리아 융합 조절에 관여하는 Mfn2 단백질이 PD98059 농도 의존적으로 증가되어 미토콘드리아
융합을 유도하였음을 확인하였다. 다음으로, 활성화 형태의 AKT 와 JNK 확인을 통해 PD98059 의 side effect 가 없고 미토콘드리아 형태 변화가 뚜렷한 60μM 로 농도를 고정한 후, ERK1/2 활성 저해에 따른 미토콘드리아 길이와 Mfn2 단백질 량의 변화를 시간 별로 확인 해 보았다. 60μM 의 PD98059 처리하고 18 시간 이후 미토콘드리아가 길어지고 Mfn2 단백질 역시 증가 되는 것을 관찰하였다. 활성화 형태의 ERK1/2 는 PD98059 처리 후 6 시간부터 억제 되었지만, 미토콘드리아 형태와 Mfn2 단백질 량의 변화는 없었는데, 이는
아마도 Mfn2 단백질의 turn over 시간의 영향에 의한 것으로 생각된다. ERK1/2 활성 저해에 따른 Mfn2 단백질의 특이적 증가가 미토콘드리아 형태를 길어지게 하는 사실을 각기 다른 실험을 통하여 반복 확인하였으며, Mfn2 는 미토콘드리아 외 막에 위치하여 제 역할을 수행하는 것으로 잘 알려져 있다. PD98059 처리가 Mfn2 의 세포 내 위치변화에 영향을 미치는지 미토콘드리아 분획 실험과 Mfn2 항체를 이용한 면역 세포 화학 방법을 통해 확인 해 보았는데, PD98059 처리 후에도 Mfn2 가 미토콘드리아에 위치하는 것을 확인하였다. ERK1/2 활성 저해에 의한 Mfn2 단백질 량 증가 원인 규명 연구를 위하여 Mfn2 의 증가가 mRNA 수준에서 유전자 발현(expression) 증가에 의한 것인지 알아보고자 RT-PCR 을 수행 하였으나 발현의 차이가 없었고, 다음으로 단백질 분해 경로에 관여하는 유비퀴틴화(ubiquitination) 분석을 통하여 Mfn2 의 증가가 단백질 수준의 안정성(stability) 증가에 의한 것인지 확인하였다. 결과적으로 PD98059 처리에 따른 ERK1/2 의 활성 저해에 의해 Mfn2 의 유비퀴틴화가 감소 되어 증가된 안정성에 의해 단백질이 증가되어 미토콘드리아가 길어졌으며 mRNA level 에는 변화가 없음을 확인하였다. Mfn2 의 유비퀴틴화 감소 원인을
확인 하기 위하여 지금까지 Mfn2 의 유비퀴틴화에 관여하는 것으로 보고된 2 종류의 E3 ubiquitin ligase 인 MARCH-V 와 Parkin 을 분석한 결과, PD98059 처리에 의한 ERK1/2 활성 저해는 MARCH-V 단백질의 감소를 유도 한다는 흥미로운 사실을 발견하였고, 이를 통해 Mfn2 의 유비퀴틴화 감소는 MARCH-V 의 감소로 인한 결과라 생각되고, 향후 MARCH-V 와 Mfn2 의 직접적인 연관성 관찰을 통해 MARCH-V 가 매개하는 ERK1/2 에 의한 Mfn2 조절을 확인 해 볼 계획이다. 결론적으로, ERK1/2 의 활성 변화는 E3 ubiqiutin ligase 인 MARCH-V 의 발현 량에 영향을 미치고, 이로 인해 미토콘드리아 형태조절 단백질 Mfn2 의 안정성의 변화를 가져와 결과적으로 미토콘드리아의 형태 변화를 초래한다는 사실을 밝혔다. 본 연구 결과는, 세포의 항상성 유지에 필수적이라고 알려진 미토콘드리아의 형태조절이 ERK1/2 신호전달 경로에 의해 조절되어 그 균형을 유지되고 있음을 시사한다. 이와 같은 연구결과를 토대로 미토콘드리아 형태조절에 의해 매개 되는 다양한 생명현상 이해 증진에 크게 기여 할 것이라 생각한다.
V. 결 론
본 연구에서는 미토콘드리아 형태조절 단백질들의 발현이 세포 내 어떤 신호전달 경로에 의해 조절되는지 그 분자 기전을 규명하고자 하였다. ERK1/2 의 활성화 정도에 비례하여 미토콘드리아가 짧아진다는 사실의 발견으로부터 연구를 시작하여, 여러 종류의 세포에 MEK/ERK 특이적 저해제인 PD98059 를 처리하여 ERK1/2 의 활성을 억제한 결과 시험한 모든 세포에서 미토콘드리아가 길어지는 것을 확인 하였다. 분자기전 연구를 통하여, ERK1/2 의 활성 변화는 미토콘드리아에 국재 하는 E3 ubiquitin ligase 인 MARCH-V 의 단백질 량에 영향을 미치고, 이로 인해 미토콘드리아 형태조절 단백질 Mfn2 의 안정성의 변화를 가져와 결과적으로 미토콘드리아의 형태 변화를 초래한다는 사실을 밝혔다.참고문헌
1.
Amati-Bonneau P, Milea D, Bonneau D, Chevrollier A, Ferre M, Guillet V,
Gueguen N, Loiseau D, de Crescenzo MA, Verny C, Procaccio V, Lenaers G,
Reynier P: OPA1-associated disorders: phenotypes and pathophysiology. Int
J Biochem Cell Biol 41: 1855-1865, 2009
2.
Anesti V, Scorrano L: The relationship between mitochondrial shape and
function and the cytoskeleton. Biochim Biophys Acta 1757: 692-699, 2006
3.
Castanier C, Arnoult D: [Mitochondrial dynamics during apoptosis]. Med Sci
(Paris) 26: 830-835, 2010
4.
Chan DC: Mitochondria: dynamic organelles in disease, aging, and
development. Cell 125: 1241-1252, 2006a
5.
Chan DC: Mitochondrial fusion and fission in mammals. Annu Rev Cell Dev
Biol 22: 79-99, 2006b
6.
Cheung EC, McBride HM, Slack RS: Mitochondrial dynamics in the
regulation of neuronal cell death. Apoptosis 12: 979-992, 2007
7.
Detmer SA, Chan DC: Functions and dysfunctions of mitochondrial
dynamics. Nat Rev Mol Cell Biol 8: 870-879, 2007
8.
Grandemange S, Herzig S, Martinou JC: Mitochondrial dynamics and cancer.
Semin Cancer Biol 19: 50-56, 2009
9.
Hom J, Sheu SS: Morphological dynamics of mitochondria--a special
emphasis on cardiac muscle cells. J Mol Cell Cardiol 46: 811-820, 2009
10.
Kuznetsov AV, Hermann M, Saks V, Hengster P, Margreiter R: The cell-type
specificity of mitochondrial dynamics. Int J Biochem Cell Biol 41: 1928-1939,
2009
11.
Martinou I, Desagher S, Eskes R, Antonsson B, Andre E, Fakan S, Martinou
JC: The release of cytochrome c from mitochondria during apoptosis of
NGF-deprived sympathetic neurons is a reversible event. J Cell Biol 144: 883-889,
1999
12.
Monick MM, Powers LS, Barrett CW, Hinde S, Ashare A, Groskreutz DJ,
Nyunoya T, Coleman M, Spitz DR, Hunninghake GW: Constitutive ERK
MAPK activity regulates macrophage ATP production and mitochondrial
integrity. J Immunol 180: 7485-7496, 2008
13.
Okamoto K, Shaw JM: Mitochondrial morphology and dynamics in yeast and
multicellular eukaryotes. Annu Rev Genet 39: 503-536, 2005
14.
Rasola A, Sciacovelli M, Chiara F, Pantic B, Brusilow WS, Bernardi P:
Activation of mitochondrial ERK protects cancer cells from death through
inhibition of the permeability transition. Proc Natl Acad Sci U S A 107:
726-731, 2010
15.
Wang X, Su B, Lee HG, Li X, Perry G, Smith MA, Zhu X: Impaired balance
of mitochondrial fission and fusion in Alzheimer's disease. J Neurosci 29:
9090-9103, 2009
16.
Zhu JH, Guo F, Shelburne J, Watkins S, Chu CT: Localization of
phosphorylated ERK/MAP kinases to mitochondria and autophagosomes in
Lewy body diseases. Brain Pathol 13: 473-481, 2003
- ABSTRACT -
Regulation of Mitochondrial Morphology by ERK1/2-Mediated
Control of Mfn2 Stability
Young Hwa Kim
Department of Biomedical Science The Graduate School, Ajou University
(Supervised by Associate Professor Seon-Yong Jeong)
Mitochondria are dynamic organelles. The major and essential role of mitochondria is to provide a myriad of services to the cell, including energy production and regulation of survival, apoptosis, metabolism, autophagy and aging. Mitochondria continuously move, fuse and divide in healthy cells. Recent studies revealed that the mitochondrial morphology is an importantdeterminant of mitochondrial function.The mitochondrial reticulum can be rapidly remodeledby dynamic mitochondrial fusion and fission events in response to the physiologicalrequirements of a cell. Mitochondrial fusion is regulated by the large GTPases mitofusins, Mfn1 and Mfn2 and optic atrophy protein 1 (Opa1) and mitochondrial fission is controlled by Fis1 and dynamin related protein 1 (Drp1) along with Mff, MTP18, GDAP1, DAP3, andpotentially other known and unknown proteins. Although the functions of the
mitochondrial shaping proteins are well known, the molecular mechanisms in regulation of mitochondria-shaping genes still remain to be clarified.
I have found that ERK1/2 activation is closely related with mitochondrial morphology in primary Ras-activated cells. The existence of ERK1/2 protein in outer membrane and inter membrane space of brain mitochondria has been reported. The ERK1/2 signaling is essential for cell growth, proliferation and survival in response to numerous cellular signals. These results have led to further study on the roles of ERK1/2 activation in regulation of mitochondrial morphology. I observed the elongation of mitochondria after inhibition of ERK1/2 activity by the MEK/ERK specific inhibitor PD98059 in primary cultured cells and cell lines. Western blot analysis of the mitochondria-shaping proteins revealed that the Mfn2 protein level was significantly increased in PD98059-treated cells. RT-PCR analysis of the MFN2 gene in PD98059-treated cells revealed that this increase of Mfn2 was not caused by the increased expression of MFN2 at the mRNA level. To elucidate the involvement of the protein stability in this increase of Mfn2 in PD98059-treated cells, I compared the ubiquitination patterns of Mfn2 protein between the PD98059-treated and non-treated cells. The ubiquitination level of Mfn2 protein in the PD98059-treated cells was much lower than that in the non-treated cells. Next, I investigated the expression levels of the E3 ubiquitin ligages, MARCH-V and Parkin, and found that the expression level of MARCH-V was significantly decreased in the PD98059-treated cells compared to the non-treated cells.
These results suggest that the expression level of MARCH-V may be regulated by ERK1/2 activation level, which leads to the regulation of the Mfn2 protein stability inked to mitochondrial morphology.
Key words: mitochondria, mitochondria morphology, mitochondria fusion/fission, ERK1/2, Mfn2, MARCH-V, ubiquitination