저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
I
gE-r
eact
i
vi
t
y andpr
oi
nf
l
ammat
or
y cyt
oki
ne
-i
nduci
bi
l
i
t
y ofvar
i
ousr
ecombi
nanthousedust
mi
t
eal
l
er
gensi
ncl
udi
ng novelr
Derf4
by
Chung-RyulKi
m
Maj
ori
n Mol
ecul
arMedi
ci
ne
Depar
t
mentofBi
omedi
calSci
ences
TheGr
aduat
eSchool
,Aj
ou Uni
ver
si
t
y
I
gE-r
eact
i
vi
t
y andpr
oi
nf
l
ammat
or
y
cyt
oki
ne-i
nduci
bi
l
i
t
y ofvar
i
ousr
ecombi
nanthouse
dustmi
t
eal
l
er
gensi
ncl
udi
ng novelr
Derf4
by
Chung-RyulKi
m
A Di
sser
t
at
i
on Submi
t
t
ed t
oTheGr
aduat
eSchoolof
Aj
ou Uni
ver
si
t
y i
n Par
t
i
alFul
f
i
l
l
mentoft
heRequi
r
ement
sf
or
TheDegr
eeofPh.
D.ofBi
omedi
calSci
ences
Super
vi
sedby
Ho-J
oon Shi
n,Ph.
D.
Maj
ori
n Mol
ecul
arMedi
ci
ne
Depar
t
mentofBi
omedi
calSci
ences
TheGr
aduat
eSchool
,Aj
ou Uni
ver
si
t
y
Thi
scer
t
i
f
i
est
hatt
hedi
sser
t
at
i
on
ofChung-RyulKi
m i
sappr
oved.
SUPERVI
SORY COMMI
TTEE
SunPar
k
Ho-J
oon Shi
n
Myung-HeeKwon
Kyongmi
n Ki
m
Tai
-Soon Yong
TheGr
aduat
eSchool
,Aj
ou Uni
ver
si
t
y
December
,18t
h,2015
-Abstract
-I
gE-r
eact
i
vi
t
y andpr
oi
nf
l
ammat
or
y cyt
oki
ne-i
nduci
bi
l
i
t
y
ofvar
i
ousr
ecombi
nanthousedustmi
t
eal
l
er
gens
i
ncl
udi
ng novelr
Derf4
House dustmite (HDM) is the mostimportant inducer ofallergy and asthma in the world.More than thirty three group ofmite allergens have been reported from HDMsand storagemites(SMs).Theaccuratediagnosis ofmiteallergy isavery importantforeffectivetreatment.Fordiagnosisand immunotherapy ofmite allergy,the crude extractofmites have been used, which may cause side effects. Recently, specific allergens selectively developed using molecularbiologicaltechnology cameintothespotlight.This study intendedtoexaminediagnosticandimmunotherapeuticpotentialsofthe recombinantallergensfrom housedustmiteDermatophagoidesfarinae.
Thirteen D.farinae-derived recombinantallergens were produced using
Escherichia coliorPichia pastoris expression system.Native Derf4 was also purified forcomparison.rDerf1(89.2%)and rDerf2(96.4%)showed high reactivity to IgE in Korean HDM-sensitized patient’s sera.rDer f 3 (61.9%),nDerf4 (75.8%),rDer f5 (64.9%),rDerf7 (43.8%),rDer f21 (66.0%),and rDerf 23(43.3%)exhibited moderateIgE reactivity them.IgE reactivityofrDerf10(24.7%),rDerf13(3.6%),rDerf15(4.1%),rDerf18 (32.0%)and rDerf20 (38.7%)displayed low IgE reactivity.IgE reactivities againstDerf2 were shown in allpatients with asthma,rhinitis oratopic dermatitis.Each serum recognized with 2 to 12D.farinae proteins,and 190
of194(97.9%)recognizedwiththreeormoreproteins.
A cross-reactivity ofrecombinantgroup 5 and 21 allergens from three mite species (D.farinae,Tyrophagus putrescentiae,and Blomia tropicalis) wasevaluated.Cross-reactivity between rDerf23and rDerp 23 werealso examined by using inhibition-ELISA.IgE reactivity to rDerf5 was highly correlatedwith doseofrDerf21(r=0.733).rTyrp5exhibitedthehighest levelofcorrelation with rTyrp 21 (r= 0.950).rDerf21 exhibited a high inhibitory effect against rTyr p 5 (93.0%),rTyr p 21 (92.1%),rBlo t 5 (87.0%),and rBlot21(70.3%),implying cross-reactivity acrossmitespecies. rDerp23effectivelyinhibitedaIgE bindingtorDerf23.
Notonly ability ofIgE responsesbutalso capability to induceinflammatory cytokine production is importantproperties ofallergens.In this study,IL-6 and IL-8 production in BEAS-2B cells (bronchialepithelialcell-line) were examined by challenging oftherecombinantmiteallergens.rDerf4,rDerf 5,rDerf10and rDerf21induced IL-6orIL-8secretionsin BEAS-2B.In particular,rDer f21 could trigger IL-6 and IL-8 production in BEAS-2B throughTLR4-dependentsignaling.
In summary,Derf1 and Derf2 was shown to be the mostpotent allergens.Derf3,Derf4,Der5,Derf7,Derf21andDerf23werefound tobethemajorallergenssensitizedmorethan 50% ofHDM allergy patients. Derf21showed cross-reactivity toT.putrescentiaeandB.tropicalisgroup 5 and 21 allergens.In addition,Derf21 wasfound to triggerofIL-6 and IL-8productioninabronchialepithelialcelllinethroughaTLR4.
The information on allergenicity and IgE reactivities of various mite allergens shown in this study will be useful for development of a component-resolveddiagnosisandeffectivetreatmenttoHDM allergy.
Key words: house dust mite, storage mite, allergy, IgE reactivity, cross-reactivity, component-resolved diagnosis, recombinant allergen,
TABLE OF CONTENTS
ABSTRACT ··· i
TABLE OF CONTENTS ···iv LIST OF FIGURES ··· vi
LIST OF TABLES···ix I.INTRODUCTION ··· 1
1.Overview ofhousedustmiteallergens ··· 1
2.Component-resolveddiagnosis ··· 2
3.Cross-reactivityofmiteallergens··· 3
4.Recombinantmiteallergensindiagnosisandtreatment ··· 4
5.Immuneresponseonairwayepithelialcellbymiteallergens ··· 6
6.Aimsofthisstudy ··· 7
II.MATERIALS AND METHODS ···11
1.Subjectsandserum samples ···11
2.PreparationofnativeDerf4andotherrecombinantmiteallergens ·11 1)PurificationofnativeDerf4(nDerf4) ···11
2)CloningofDerf4fullcDNA sequence ···13
3)ExpressionofrecombinantallergensinE.coliusingthepET expressionvector ···14
4)ExpressionofrecombinantallergensinP.pastorisusingthe pPIC9expressionvector ···15
5)Removalofendotoxinfrom therecombinantallergens(proteins) ···17
3.IgE reactivityofrecombinantmiteallergens ···17
4.InhibitionELISA and3D proteinstructuremodeling ···18
5.StimulationofBEAS-2B cellswithrecombinantmiteallergens ···19
2)Determinationofcytokineproduction ···19
III.RESULTS ···23
1.IgE reactivityprofileofKoreanmite-sensitizedallergicpatientsto recombinantD.farinaeallergens ···23
1)IgE reactivitytoD.farinae,T.putrescentiaeandB.tropicalis extracts ···23
2)PreparationofnativeandrecombinantDerf4 ···26
3)IgE reactivitiesandcorrelationsofthirteenD.farinaeallergens ····38
2.Evaluationofcross-reactivityamongmiteallergens ···50
1)Cross-reactivitiesofsixrecombinantgroup5and21mite allergens ···50
2)Cross-reactivitybetweenrDerf23andrDerp23 ···63
3.Productionofproinflammatorycytokines,IL-6andIL-8,inBEAS-2B bystimulationofrecombinantallergensderivedfrom HDM, D.farinae ····68
IV.DISCUSSION ···76
V.CONCLUSION ···82
REFERENCE ···83
LI
ST OF FI
GURES
Fig.1.IgE reactivitiestoD.farinae,T.putrescentiaeandB.tropicalis
crudeextracts···24 Fig.2.CorrelationsamongtheIgE reactivitiesofD.farinae,T.
putrescentiaeandB.tropicaliscrudeextracts ···25 Fig.3.ThenucleotideanddeducedaminoacidsequencesofDerf4 ···27 Fig.4.cDNA sequencealignmentofDerf4isoform clones ···28 Fig.5.Aminoacidssequencealignmentof5mitespeciesgroup4
allergen ···29 Fig.6.VectormapandSDS-PAGE analysisofnativeandrecombinant
Derf4 ···31 Fig.7.AmylaseactivityofD.farinaecrudeextract,nDerf4,and
rDerf4 ···32 Fig.8.SDS-PAGE andWesternblottingofnativeandrecombinant
Derf4 ···35 Fig.9.IgE reactivityofD.farinae-sensitizedhumanseratonDerf4
andrDerf4anditscorrelation ···36 Fig.10.IgE reactivityofD.farinae-sensitizedhumanseraand
correlationstonDerf4andGermancockroach(B.germanica)
α-amylase ···37 Fig.11.SDS-PAGE profileofrecombinantD.farinaeallergens ···39 Fig.12.IgE reactivitiestorecombinantD.farinaeallergens ···41 Fig.13.CorrelationsofIgE reactivitiestorDerf1,rDerf2and
rDerf3withD.farinaeextract ···42 Fig.14.IgE reactivityprofilestoD.farinaeallergensdepending
onclinicalsymptoms ···44 Fig.15.IgE reactivityprofilestoD.farinaeallergensdepending
onage ···47 Fig.16.Profileofallergennumberagainstoneserum ···49 Fig.17.SequencecomparisonofrecombinantDerf5andothergroup5
andgroup21allergens ···51 Fig.18.SDS-PAGE ofrDerf5,rDerf21,rTyrp5,rTypp21,
rBlot5andrBlot21 ···54 Fig.19.TheIgE reactivitiestosixrecombinantgroup5and21
allergens ···56 Fig.20.CorrelationsbetweentheIgE reactivitiesofrecombinant
allergens ···57 Fig.21.CompetitiveIgE inhibitionELISA ofgroup5and21allergento
crudeextract ···60 Fig.22.Inhibitionanalysesofsixrecombinantallergens ···61 Fig.23.Predicted3D-structureofrecombinantgroup5and21
allergens ···62 Fig.24.SequencecomparisonandSDS-PAGE ofrDerf23and
rDerp23 ···64 Fig.25.InhibitionELISA ofrDerf23andrDerp23 ···67 Fig.26.SDS-PAGE profileofD.farinae,D.pteronyssinusandT.
putrecentiaecrudeextracts ···69 Fig.27.Threespeciesmiteextracts-inducedCCL20,IL-6andIL-8
secretioninBEAS-2B cells ···70 Fig.28.rDerf4-inducedIL-6andIL-8secretioninBEAS-2B cells ····71 Fig.29.rDerf21-inducedIL-6andIL-8secretioninBEAS-2B cells ·· 72 Fig.30.rDerf21-inducedIL-6andIL-8secretioninBEAS-2B cells
withTLR4antagonist ···73 Fig.31.rDerf5-inducedIL-6andIL-8secretioninBEAS-2B cells ····74 Fig.32.rDerf10,rDerf13,rDerf23andrDerp23-inducedIL-6
LI
ST OF TABLES
Table1.ListofHDM allergens ···9 Table2.Listofcloningprimersequences ···21 Table3.α-amylaseactivityofD.farinaecrudeextract,nDerf4,and
rDerf4 ···33 Table4.PrevalenceagainstD.farinaeallergensperclinicaldiagnosis ···45 Table5.PrevalenceagainstD.farinaeallergensperage ···48 Table6.Sequenceidentityofgroup5andgroup21allergensfrom
D.farinae,T.putrescentiae,andB.tropicalis ····52 Table7.Characteristicsofpatients’serausedinthecompetitive
ELISA ···59 Table8.Characteristicsofpatients’serausedinthecompetitive
I
.I
NTRODUCTI
ON
1.Overview ofhousedustmiteallergens
TheHDMsandSMsarethemajorcauseofrespiratory allergicdiseases (Yu etal,2014;Fernández-Caldas etal,2008).These mite allergens were classified according to a Linnean system ofnomenclature thatis maintained by the World Health Organization (WHO) and the InternationalUnion of Immunological Societies (IUIS) Allergen Nomenclature Sub-Committee (Radaueretal,2014).Morethan 34differentgroupsofmiteallergens,which elicits specific IgE responses in sensitized patients were identified in
Dermatophagoides spp. (Jacquet, 2011a; Fernández-Caldas et al, 2007; GregoryandLloyd,2011;http://www.allergen.org).
Allergennomenclatureisfollows:combinationofthefirstthreelettersof the genus name,the first letter of the species name and a number.For example,in the majorhouse dustmite allergen “rDerf1”,“r” stands for recombinant(“n” fornative),“Der” forthe firstthree letters ofthe genus (Dermatophagoides),“f”forthe firstletterofthe species (farinae)and the number “1” for the numbering of the allergen (usually in the order of reporting). Notwithstanding reporting order,similar allergens from related speciesusethesamenumbering.HDM α-amylaseallergensfrom thespecies
Dermatophagoides pteronyssinus,Euroglyphus maynei,and B.tropicalis are individually referred to as Derp 4,Eurm 4,and Blo t4,respectively,or collectively asthegroup4miteallergens.Withisoallergens,derivedfrom the samespeciesdemonstrating morethan 67% sequenceidentity,a suffix (e.g., Der p 1.01,Der p 1.02) is used.For isoallergens that differ by a few residues,an additionaltwo digits are added (e.g.,Derp 1.0101)to identify
them (BreitenederandChapman,2014).
HDM allergenscould bedivided into groupson thebasisofbiochemical properties and functions where these are known or can be deduced. It includes six ‘families’:the proteases (mite allergen groups 1,3,6 and 9), glycosidases(groups4,12,15and18),transferases(groups8and 20),small α-helicalproteins (groups 5,7 and 21),muscle proteins (10,11,26,33 and 34),thebindingproteins(2,13,14,23,24and30)andunclassified(16,17,19, 22,25,27,28,31and32)(BessotandPauli,2011;http://www.allergen.org).
An allergen isconsidered “major”when theallergen elicitsareaction in 50% ormore ofan allergic population and “minor”when less than 50% of theallergicpopulation respondstotheallergen.Traditionally,majorallergens are considered more important,butsometimes otherallergens can be more importantforitsabilitytoelicitsevereclinicalsymptoms.
2.Component-resolveddiagnosis
Component-resolved diagnosis(CRD)isconsidered usefulin diagnosisof allergy recently.Itutilizes purified native orrecombinantallergens to detect specific IgE (sIgE) reactive to individualallergen molecules (Treudler and Simon,2013).Traditionalin vitro diagnosis ofallergic diseases have been based on detecting sIgE to totalextracts which contain both allergenic and non-allergenic components.In CRD,purified single allergens are used to establish a sensitization profilein theindividualallergicpatient,and thereby clinically relevantinformation can be added.In some cases,single allergens mayevenyieldahigherdiagnosticsensitivitythannaturalextracts(Carvalho etal,2013;Valentaetal,1999;Yasuharaetal,2001).TheadvantageofCRD is as follows: first,Increasing the accuracy of tests (detecting clinically
relevant allergenic components from allergenic sources);second,Predicting cross-reactivity to proteinswith similarstructures;third,Assessing risk and type of clinical reactions; fourth, Providing information important to the dietarymanagementofpatients,e.g.foodsourcescontaining relevantallergen; fifth,Selecting thepatientssuitabkeforspecificimmunotherapy (SIT)(Lloyd, 2014).CRD have become ofgrowing importance in clinicalinvestigation of IgE-mediatedallergies.
3.Cross-reactivity ofmiteallergens
The cross-reactivity between epitopes results from their structural similarity and may inducethecross-binding oftheIgE and allergicresponse in a patientprimarily non-sensitized allergenic protein (Jenkins etal,2005). Usually these epitopes are formed by the polypeptide chain ofthe allergen. Allergenic determinants in D.farinae are shared with othermites belonging to the Pyroglyphidae family and are highly cross-reactive with other
Dermatophagoides species (Thomas etal,2002).Also,cross-reactivity has been reported between allergens ofHDMs and otherinvertebrates such as crabs,shrimps and snails (Sidenius,2001).Unlike with Dermatophagoides
species, there seems to be a limited degree of cross-reactivity with nonpyroglyphid mites (storage mites) (Ferrandiz et al,1998;Thomas and Smith,1998).
In general,as a number ofspecific allergens in D.pteronyssinus are homologous with otherallergens,varying degrees ofcross-reactivity can be expected(Thomasetal,2010).Forexample,Derp4andEurm 4share90% sequenceidentity,andwerealsocalculatedtobeapproximately 50% identical toinsectandmammalianα-amylases(Millsetal,1999).
Tropomyosin,group10miteallergen,isawell‑known pan-allergen that causes cross-reactivity among mites and a variety of invertebrates (eg. shrimps and crabs)(Reese etal,1999;Jeong etal,2006),even to parasites (eg.Anisakisnematodesandroundworms)(Gamezetal,2011;Nieuwenhuizen and Lopata,2013;Acevedo and Caraballo,2011).Group 2 miteallergens are theprimary allergensresponsibleforthecross-reactivity between HDMsand SMs(Parketal,1999;Liaoetal,2010).
There is about 40% sequence identity between group 5 and 21 mite allergens, and expected specific IgE to be cross-reactive. However, no correlation between Blo t 5 and Der p 5 in eliciting IgE responses was observed in B.tropicalis exposed and unexposed individuals,despite 43% identityattheaminoacidlevel (Arrudaetal,1995).Similarly,inastudyon Malaysian and Taiwanesepatients,low correlation ofIgE reactivity between Blo t5 and Derp 5 wasobserved (Taiwanese,r=0.452;Malaysian,r=0.346) as wellas limited cross‑reactivity (Kuo etal,2003).Otherstudy showed thatBlo t21 was nothighly cross‑reactive to Blo t5 despite a certain amount of sequence and structuralidentity (Gao et al,2007).Der p 21 exhibitedlow IgE cross-reactivitytoDerp5,Lepd5andBlot5according totestsusing serafrom Derp21-sensitizedpatients (Weghoferetal,2008). In addition,Blo t21 wasreported to have low to moderatecross-reactivity with Blo t5,Derp 5 and Derf21 (Tan etal,2012).However,intensities and frequencies of IgE cross-reactivity between group 5 and 21 mite allergensarestilluncertain.
4.Recombinantmiteallergensin diagnosisandtreatment
mainly performed by using crude extracts from whole mite bodies with or withoutfecalpellets.These allergen extracts contain a variety ofallergenic and nonallergenic components and their allergen contentis reported to be variabletoalargeextentby thequality oftheraw materialandthemethod ofextractionandpurification.Asfordiagnosticpurposes,positivereactionsto a given allergen extractwillthus provide the information on the source of allergens thatan allergic subjectis sensitized withoutidentity ofallergenic components.Likewise,extract-based immunotherapy cannot reflect to the individualpatient’ssensitizationpattern.
Moreover,the use offormulations with known therapeutic doses ofthe importantallergennotonly improvetheefficacybutwouldallow reproducible investigationstomadetoestablish optimalprotocols(Vrtalaetal,2014).The poorbalance ofthe concentrations ofdifferentallergens causes the inherent problem oftrying to achieve therapeutic doses with an allergen presentin low concentration when allergensathigherconcentrationshavetheability to cause anaphylactic side effects. Furthermore, allergen extracts contain molecules eliciting inflammation like ceramides and lipids as well as ß-glucansandtheendotoxin.
Recombinantallergens can be used to overcome the limitations ofthe naturalextract,asstand-alonereagentsorasamixturereplacing an extract, andtodeterminetheallergencomponentprofileofanindividualfordiagnostic or treatment purposes (Jutel et al, 2012). The concept and terms of recombinant allergen based component-resolved diagnostics (CRD) and immunotherapy (CRIT) was created in 1999 (Valenta et al,1999). This approachadvocatestheuseofwell-definedallergencomponents,fordiagnosis ofIgE-mediatedallergy.
individualallow closerexaminationsofcross-reactions,moleculesofrisk and prediction ofsymptom prognosis(Thomas,2015).Accumulation ofinformation on each allergen is very useful for development of accurate allergy diagnostictoolsandeffectiveimmunotherapeutics.
5.Immuneresponseon airway epithelialcellby miteallergens
TheHDM hasboth directand indirecteffects on theairway epithelium (Gandhietal,2013).Mediators released following house dustmite-mediated activation of mast cells or T cells,such as histamine,prostaglandins or IL-13,have multiple effects on epithelialcells (Thomas et al,2010).The epithelium therefore may translate and/or propagate HDM-induced signals from immune cells and amplify the inflammatory response withoutthe need for direct interactions with HDM components (Jacquet, 2013). However, HDMscanalsodirectlyactivatetheairwayepithelium andcausemanyofthe changesseeninallergicairwayinflammation.
Exposure to mite allergens triggers inflammation in atopic patients, including with allergicasthma,rhinitisand atopicdermatitis(Jacquet,2011b). Theairway epithelium isthefirsttissueto beencountered by theantigens. The epithelium expresses many pattern recognition receptors (PRRs) that mediatean innateimmuneresponsetomite(ParkerandPrince,2011).Innate cellactivation is mainly mediated via a limited numberofgermline encoded receptors,the PRRs,which have broad specificity and recognize conserved molecularstructuresuniquetoclassesofinfectiousmicrobes(Jacquet,2011a). The mostimportantcontaminants ofhouse dustand HDM extractare toll-likereceptor(TLR)agonistlipopolysaccharides(LPS)andβ-glucans.LPS is one of the major cause of mite-induced airway inflammation which is
mediated by LPS/TLR4 dependent signal pathway (Hammad et al,2009; Hongjiaetal,2010).HDM-derivedβ-glucan actsthroughTLR2anddectin-1 (Ryu etal,2013;Nathan etal,2009).Thesemolecules arealso endogenous danger signals called damage-associated molecular patterns (DAMPs) (Jacquet,2011a;Thomas,2010).
Theproteinaseactivity ofallergensin themiteextractcan alsoplay an importantrole in allergic sensitization and inflammation.Group 1 (cysteine protease),3,6 and 9 (serineproteinase)may exerttheireffectsthrough the activation of a receptor expressed on the epithelium, called proteinase activated receptor(PAR),mainly PAR-2 (Jeong etal,2008;Roelandtetal, 2008;Postetal,2014,Shietal,2010,Sunetal,2001).
Mite extracts were shown not only to induce increases in the permeability of bronchial mucosa but also to trigger the release of pro-inflammatory cytokinessuch asIL-6,IL-8,monocytechemotacticprotein (MCP)-1 (CCL20),GM-CSF and IL-1β in activated airway epithelialcells (Kauffman etal,2006;Osterlund etal,2009;Wan etal,1999,Wan etal, 2001). Similar pro-inflammatory cytokine up-regulation was observed in primary bronchialepithelialculturesfrom healthy individualsorHDM-allergic patients(GregoryandLloyd,2011).
6.Aimsofthisstudy
Although D.farinae is a dominantspecies ofHDM in Korean domestic environments,studies on the immunology ofeach D.farinae allergen and relationship with allergens from other sources are stillunder investigation. Identification of the recombinant mite allergens potentially useful for the developmentofCRD and SIT isimportantin mite-induced allergicdiseases.
Thepurposeofthisstudy istoevaluateallergenicity and cross-reactivity of recombinantallergens from D.farinae,T.putrescentiae and B.tropicalis.In thisstudy,anumberofrecombinantallergensofD.farinae,D.pteronyssinus, T.putrescentiae and B.tropicalis were produced in E.coliorP.pastoris
expression system toprovidebasicinformation onthedevelopmentofuseful recombinantmiteallergens.TheirIgE reactivity with Korean mite-sensitized allergypatient’ssera,andthelevelofsensitizationtoeachallergendepending on theclinicalsymptomsand agewereanalyzed.Then,IgE cross-reactivity between group 5 and 21 mite allergens was examined.IgE cross-reactivity betweenDerf23andDerp23,thenewlyidentifiedHDM allergenswasalso studied.Finally,productionofpro-inflammatorycytokinebystimulationofthe recombinantmiteallergensin BEAS-2B cellswasinvestigated.Theseworks in this study showed that these recombinant mite allergens are effective candidate molecules for development of CRD and SIT to mite allergic diseases.
Table1.ListofHDM allergens
Group Biochemicalfunction MW(kDa) Species1 IgE binding2
1 Cysteineprotease 27 Dp,Df,Dm,DsEm,Bt 80
2 NPC2family 15 Dp,Df,Ds,Em,Ld,Tp,Gd,As 80
3 Trypsin 29 Dp,Df,Ds,Em,Bt,Tp 16-100
4 Alphaamylase 60 Dp,Df,Em,Bt,As 40-46
5 Unknown 14 Dp,Df,Bt,Ld 50-70
6 Chymotrypsin 25 Dp,Df,Bt 40
7 Unknown 30-31 Dp,Df,Ld 50
8 GlutathioneS-transferase 32 Dp,Df,Bt 20-40 9 Collagenolyticserineprotease 29 Dp,Df 90
10 Tropomyosin 37 Dp,Df,Bt,Ld,Tp 50-95
11 Paramyosin 98 Dp,Df,Bt 80
12 Unknown 15 Bt 50
13 Fattyacidbindingprotein 14 Df,Bt,Ld,Tp,As 10-20
14 Apolipophorin 177 Dp,Df,Em 90
15 Chitinase 98/109 Dp,Df 70
16 Gelsolin/villin 53 Df 50
17 Calcium bindingprotein 53 Df 35
18 Chitin-bindingprotein 60 Dp,Df,Bt 55
19 Anti-microbialpeptidehomologue 7 Bt 10
Table1.ListofHDM allergens(continued)
Group Biochemicalfunction MW(kDa) Species1 IgE binding2
21 Unknown 14 Dp,Df,Bt 40-50
22 Unknown Df 50
23 Peritrophin-likeprotein 8 Dp,Df 70-80
24 Ubiquinol-cytochromecreductasebindingproteinhomologue 13 Df 50*
25 Trisephosphateisomerase 34 Df 75.6*
26 Myosinalkalilightchain 18 Df 29.4*
27 Serpin 48 Df 42.1*
28 Heatshockprotein 70 Df 11.5*
29 Peptidyl-prolylcis-transisomerase 16 Df 21*
30 Ferritin 16 Df 63.4*
31 Cofilin 15 Df 31*
32 Secretedinorganicpyrophosphatase 35 Df 15.4*
33 Alpha-tubulin 52 Df 25*
34 TroponinC 18 TP 10.6
1.Species designated by initials:Dp,Dermatophagoides pteronyssinus;Df,Dermatophagoidesfarinae;Em,Euroglyphusmaynei;Ds,Dermatophagoides siboney;Dm,Dermatophagoides microceaus;Ld,Lepidoglyphus destructor;Bt,Blomia tropicals;Tp,Tyrophagus putrecentiae;Gd,Glycophagus domesticus;As,Ascarissiro.
2.Bindingfrequency(% patients,variationduetodifferentstudies)
*
I
I
.Mat
er
i
al
sandmet
hods
1.Subjectsandserum samples
Serum samples were obtained from patients attending the Allergy‑Asthma Clinic atSeverance Hospital,YonseiUniversity College of Medicine in Seoul,Korea. Allergy was diagnosed on history of allergic reactionsand positiveskin prick testing.Sera from patientsweretested for IgE antibodiesspecificto HDM (D.farinae)allergensusing theImmunoCAP system (Thermo Fisher Scientific, Uppsala, Sweden) and enzyme-linked immunosorbent assay (ELISA). Sera from 194 HDM‑sensitized subjects (males/females,103:91;average age,25.4 years;age range,4-67 years)and 20 healthy controls were used to assess the IgE reactivity ofrecombinant proteins.Serum samples were collected after consentof the patients,and experiments using the collected sera were approved by the Institutional Review Board (no.4-2009-0180).The study was approved by the Ethics CommitteeofSeveranceHospital(YonseiUniversityCollegeofMedicine).
2.Preparation ofnativeDerf4andotherrecombinantmite allergens
1)Purification ofnativeDerf4(nDerf4)
(1)PurificationofnDerf4
acetate,pH 5.2,containing 5 mM CaCl2,10% glyceroland proteaseinhibitor
cocktailsetIII(Calbiochem,San Diego,CA,USA).Ethanolwasadded to a finalconcentration of40%,keeping thesamplesat4℃,and themixturewas centrifuged(8,000g,10min,4 ˚C).Thesupernatantwastreatedwith0.2% glycogen for 5 min at 4 ˚C,and the α-amylase-glycogen complex was collectedbycentrifugation(8,000g,10min,4˚C).Theprecipitatewaswashed with the extraction buffer containing 40% ethanolThe finalsedimentwas incubated with rotation (2 h,room temperature) in 50 mM MES,pH 6.0, containing 5 mM CaCl2 and 10% glycerol,and dialyzed againstthe same
buffer.ThepurityoftheisolatedDerf4wasconfirmedby10% SDS-PAGE, and its concentration was determined by Bradford assay (Bio‑Rad Laboratories,Hercules,CA,USA).
(2)N-TerminalSequencing
Purified nDerf4 was transferred onto PVDF membrane and visualized by Western blotting with Derf4 reacted patient’s sera and/orPonceau S staining.ThePVDF membranewith theexactmolecularmasswereexcised andsubjectedtoEdmandegradationusing aProcise491HT proteinsequencer (Applied Biosystems,USA) at the Korea Basic Science Institute (Seoul, Korea).
(3)α-amylaseassay
α-amylase assay of native and recombinant Der f 4 was performed according to the protocoldescribed in the Amylase assay kit’s usermanual (Abcam,Cambridge,UK).
2)Cloning ofDerf4fullcDNA sequence
(1)5’and3’RapidamplificationofcDNA Ends
The 5’rapid amplification ofcDNA ends (RACE)-Ready cDNA and 3’ RACE-Ready cDNA were generated by following the protocoldescribed in the Smart RACE cDNA amplification kit’s user manual (Clontech Laboratories,PaloAlto,CA,USA).TotalRNA wasisolatedfrom frozen mite bodies using TRIzolreagent (Invitrogen Life Technologies,Carlsbad,CA, USA),according to themanufacturer'sinstructions.First‑strand cDNA was synthesizedfrom 5μg totalRNA withspecificprimersandreagentsprovided by the kit in the presence of Moloney murine leukemia virus (MMLV) reversetranscriptase.Subsequently,the5’RACE wasperformedusing the5’ UniversalPrimerMix (ClontechLaboratories)andagene-specificprimer1,5’ -TTG TTC ATC ACC ACT ACG TGT TCG TAT ATC-3’,which was designed based on theconsensusnucleotidesequencesofothergroup 4mite allergens.However,2 moregene-specific primers (GSP 2 and GSP 3)were usedforthegenerationof3’RACE fragments.Positiveandnegativecontrols werepreparedaccording totheprotocolofthemanufacturer.GSP 2,5’-GGT TAT GGT GGT GTA CAG GTT TCA CC-3’,and GSP 3,5’-GGC CGC ATT ATT GAA TTC CGT TTT TAT AAA-3’,weredesignedbasedonthe nucleotidesequencesofgroup4allergensofmitespecies.Theconditionsfor PCR amplification were as follows:Pre-denaturation at95˚C for5 min,30 cycles of denaturation at 94˚C for 30 sec,annealing at 55˚C for 30 sec, extension at 72˚C for 1 min and,following the finalcycle,an additional extension at72˚C for5 min.ThePCR‑amplified cDNA sequencesencoding
allergens were cloned into the pCR4‑TOPO vector (Invitrogen), and sequenced.Clone 1 was isolated from the 5’RACE productwith GSP 1, whereasclones2and 3wereisolated from the3’RACE productwith GSPs 2and 3.TheirDNA sequenceswereconfirmed tobethe3’end,themiddle fragment and the 5’end of Der f 4 cDNA,by overlapping homologous sequencesamongtheseclones.
(2)GenerationofFull-LengthDerf4
The template DNA was the 5’ and 3’ RACE products previously generatedfrom thetotalmRNA ofthemites.Thesequencesofclones1and 3 were used to design the forward primer:5’-CTC GAG AAA AGA GAG GCT GAA GCT GAC AGT AAA TTC AGT AAT CCA CAT TTT ATT GGT-3’and the reverse primer:5’-GCG GCC GCT TAA TGG TGA TGA TGG TGA TGA GAT TCC ACA CGA GCA CCA AT-3’,respectively. Subsequently,the PCR products were purified with the Expin gelSV kit (GeneAll Co.,Seoul,Korea) and ligated into pCR4-TOPO cloning vector (Invitrogen)andthensequencedbyGeneTechCorporation(Daejeon,Korea).
3)Expression ofrecombinantallergensin E.coliusing thepET expression vector
Frozen D.farinae,T.putrescentiaeandB.tropicalismiteswereobtained from the Arthropods ofMedicalImportance Resource Bank (AMIB)atthe DepartmentofEnvironmentalMedicalBiology,YonseiUniversity College of Medicine (Seoul,Korea).TotalRNA was isolated from frozen mite bodies using TRIzol reagent (Invitrogen), according to the manufacturer's
instructions.First‑strandcDNA wassynthesizedfrom 5μg totalRNA using avian myeloblastosisreversetranscriptase(Promega,Madison,WI,USA)and an oligo (dT18) primer. Primers were designed based on the published
full‑length nucleotide sequence of each allergens. The primers were synthesized by GeneTech.The conditions for PCR amplification were as follows:Pre-denaturation at94˚C for5min,30cyclesofdenaturation at94˚C for30 sec,annealing at55˚C for30 sec,extension at72˚C for1 min and, following the finalcycle,an additionalextension at 72˚C for 5 min.The PCR‑amplified cDNA sequences encoding allergens were cloned into the pCR4‑TOPO vector (Invitrogen),sub-cloned into the pET-28a expression vector (Novagen, San Diego, CA, USA), and transformed into E. coli
RosettaTM 2 (DE3)cells (Novagen).The expression ofrecombinantproteins was induced by addition of 1 mM isopropyl-1-thio-β-galactopyranoside (IPTG,BioShop CanadaInc.,Burlington,ON,Canada)to theE.coliculture. Overexpressedrecombinantproteinwaspurifiedfrom theinsolublefraction of the cell lysate using Ni-sepharose excel resin (GE Healthcare, Uppsala, Sweden) under denaturing conditions (20 mM sodium phosphate,500 mM NaCl,500 mM imidazole,6 M urea,pH 7.4).The concentration of each recombinantprotein was determined by a Bradford assay.To confirm the purity, all the purified recombinant proteins were separated by 15% SDS‑PAGE,followed by staining with Coomassie Blue R250 (Amresco, Solon,OH,USA).
4) Expression of recombinant allergens in P.pastoris using the pPIC9expression vector
amplified by PCR using theprimersets(Table3)containing theXhoIand Not I restriction enzyme sites,respectively.The expression of the PCR productin P.pastoriswasperformed according to methodsdescribed in the manufacturer’smanual,version E.Briefly,thegel-purified PCR productwas digested with XhoIand NotI(TakaraBioInc.,Shiga,Japan),ligated with pPIC9 vector,and transformed into E.coliDH5α.The plasmid wasisolated andlinearizedwithSacI(Takara)andthentransformedintotheyeaststrain SMD1168 by lithium chloride method for protein expression.In order to identifythehighlyexpressedclones,anindividualcolonywasinoculatedin50 mlof buffered glycerolcomplex medium (1% yeast extract,2% peptone, 1.34% yeastnitrogen base with ammonium sulfate,1% glycerol,0.4 mg/L biotin and 0.1 M potassium phosphate,pH 6.0) and grown at 30˚C in a rotary-shaking incubator at 230 rpm. The cells were harvested by centrifuging at1,500g for5min atroom temperaturewhen theOD 600nm reached 2.0–6.0. The cell pellet was resuspended in 10 ml of buffered methanolcomplex medium (which is identicalto buffered glycerolcomplex medium exceptthatglycerolisreplacedwith 0.5% methanol)forinduction of therecombinantexpression.Thereafter,themedium containing theexpressed recombinantprotein was centrifuged at8,000 g for30 min at4˚C.Forthe purification ofrecombinantallergen,theculturesupernatantwassubjected to ammonium sulfate (80% saturation)precipitation at4˚C.The precipitatewas dissolved in distilled waterand then dialyzed on a Nisepharoseexcelresin (GE Healthcare, Uppsala, Sweden) in binding buffer (20 mM sodium phosphate,pH 7.4,containing 0.5M NaCland5mM imidazole).Recombinant allergenswerepurifiedfrom theculturesupernatantusing Nisepharoseexcel resin in elution buffer(20 mM sodium phosphate,pH 7.4,containing 0.5 M NaCland 0.5 M imidazole).The concentration ofeach recombinantallergen
was determined using a Bradford assay.The purified recombinantallergens wereseparatedby SDS-PAGE (rDerf1,3,5,and21:15%;rDerf4:10%, rDerf23and rDerp 23:16.5%),followed by staining with CoomassieBlue R250.
5)Removalofendotoxin from therecombinantallergens(proteins)
Following purification, all recombinant allergens for BEAS-2B cell stimulation were filtered through an Acrodisc Mustang E membrane (Pall Corporation, Ann Arbor, MI, USA) to remove endotoxin. Removal of endotoxin was confirmed using the LAL chromogenic endotoxin quantitation kit (Pierce Biotechnology, Rockford, IL, USA), which is an assay that qualitativelydetectsendotoxin(resultsnotshown).
3.IgE reactivity ofrecombinantmiteallergens
Wells on microtiter plates were coated with 100 ul/well of the recombinant protein (2 ug/mlin 50 mM sodium carbonate,pH 9.6),and washed with phosphate-buffered saline (PBS) containing 0.05% Tween 20 (PBST).For blocking, 1% bovine serum albumin (BSA; EMD Millipore, Kankakee,IL,USA) in PBST was added to the wells for 1 h at room temperature.And then,100 μlof patient's sera diluted at 1:9 in PBST containing 1% BSA wereaddedtoeach wellsandincubatedfor1hatroom temperature. Serum IgE antibodies reacting recombinant proteins were detected using biotinylated goat anti‑human IgE (Vector Laboratories, Burlingame, CA, USA) and streptavidin‑peroxidase (Sigma‑Aldrich, St.
Louis,MO,USA).Colorwas developed using 3,3',5,5'‑tetramethylbenzidine (Kirkegaard and Perry Laboratories,Gaithersburg,MD,USA)asasubstrate. Following thestop ofcolordevelopmentby adding 0.5M H2SO4,absorbance
at 450 nm was measured using Tecan sunrise microplate reader (Tecan, Salzburg,Austria)and the Magellan CE software.The cut‑offvalue was determined as the mean absorbance plus twice the value ofthe standard deviationof20negativecontrols.
4.Inhibition ELISA and3D protein structuremodeling
Theinhibitoryeffectsofeachallergenwereexaminedusingacompetitive ELISA approach.Forthese experiments,recombinantprotein was suspended in acoating buffer(2μg/ml,50mM carbonatebuffer,pH 9.6)and added to thewellsofELISA plates.Afterblocking with 1% BSA in PBST for1h at room temperature,wellswereincubated with selected serum samples(1:9;a pooled serum of10 subjects)thathad been pre-incubated overnightat4˚C with solutionscontaining variousconcentrations(0.001,0.01,0.1,1.0or10.0μ g/ml) of recombinant proteins.IgE antibodies were detected as described above.Thepercentageofinhibition wascalculated as(1‑Ai/A0)×100,where
Ai stands for absorbance at 450 nm with an inhibitor and A0 for the
absorbanceat450nm withoutan inhibitor.Theseassayswereconducted in duplicate.
A 3D modelofsix group 5and 21miteallergenswerecreated with the SwissModelserverusing theallergen sequences(GenBankaccession No:Der f 5,ABO84970; Der f 21,ABO84964; Tyr p 5,AAX34057; Tyr p 21, AAX34059; Blo t 5, AAD10850; Blo t 21, ABH06344) and the nuclear
magneticresonance(NMR)structureofBlot5(PDB:2JMH)wasusedasa template.
5.Stimulation ofBEAS-2B cellswith recombinantmiteallergens
1)Cellculture
Human bronchialepithelialcellline BEAS-2B cells (ATCC,Manassas, VA) were cultured in a humidified atmosphere of5% CO2 at37˚C using
bronchialepithelialcellgrowth medium (BEGM;Lonza,Walkersville,MD, USA) consisting of bronchialepithelialbasalmedium (BEBM;Lonza) and growth factor supplements (human recombinant epidermal growth factor, bovine pituitary extract, hydrocortisone, insulin, transferrin, epinephrine, retinoicacid,and triiodothyronine).In each experiment,BEAS-2B cellswere seededatacelldensity of5× 104cells/well(24wellplate)andculturedfor 48 h in BEGM.Before treatment,cellculture media was changed with serum-freeBEBM overnight.
2)Determination ofcytokineproduction
Cellswerestimulatedwith100μg/mlofD.farinaeextract,rDerf4,rDer f5,rDerf10,rDerf13,rDerf21,rDerf23 and rDerp 23 for24 h in BEBM.Ascontrols,cellswerealsoincubatedwithmedium aloneorwiththe Pam3CSK4 TLR2 ligand (50 ng/ml)(Invivogen,San Diego,CA,USA)and LPS TLR4ligand (100ng/ml)(Invivogen).When appropriate,BEAS-2B cells weretreatedfor1hat37°C withanti-hTLR2monoclonalantibody(50μg/ml,
eBiosciences,San Diego,CA,USA),MAB2-hTLR4 (5 μg/ml,Invivogen), isotype control(50 μg/ml,eBiosciences)or LPS-RS TLR4 antagonist(100 ng/ml,Invivogen)priortotheaddition ofallergen.Culturesupernatantswere collected,centrifugedfor5minat10,000g,andassayedforCCL20,IL-6and IL-8withELISA kit(RnD systems,Minneapolis,MN,USA)according tothe manufacturer’sprotocol.
Table2.Listofcloning primersequences
Allergen Primer Direction
PrimerSequence Expression
Host(Vector) Derf1 forward 5′-CTCGAGAAAAGAGAGGCTGAAGCTGACA-3′ P.pastoris(pPIC9)
reverse 5′-GCGGCCGCTTAATGATGATGATGATGGTGAGAT-3′
Derf2 forward 5′-GGATCCATGATTTCCAAAATCTTGTGCCTT-3′ E.coli(pET-28b) reverse 5′-CTCGAGTTAATCACGGATTTTACCATGG-3′
Derf3 forward 5′-CTCGAGAAAAGAACACCGATTCTTCCATCATCACCA-3′ P.pastoris(pPIC9) reverse 5′-GCGGCCGCTTACTGTGAACGTTTTGATTCAATCC AATCGACA-3′
Derf5 forward 5′-GGATCCATGAAATTCATCATTGCTATTGCTG-3′ E.coli(pET-28a) reverse 5′-CTCGAGTTAAACTTCAATCTTCTTCACACGTTGCTC-3′
Derf5 forward 5′-CTCGAGAAAAGAGAGGCTGAAGCTGAACCAAAAAAACATGATTATCAAAATGAAT-3′ P.pastoris(pPIC9) reverse 5′-GCGGCCGCTTAATGATGATGATGATGGTGAACTTCAATCTTCTTCACAC G-3′
Derf7 forward 5′-AGATCTATGATGAAATTTTTGTTGATTG-3 E.coli(pET-28a) reverse 5′-GTCGACTTAATTTTTTTCCAATTCACG-3′
Derf10 forward 5′-GGATCCATGGAGGCCATCAAGAATAAAATGCAG-3′ E.coli(pET-28b) reverse 5′-CTCGAGTTAATAACCAGTAAGTTCGGCAA-3′
Derf13 forward 5′-GGATCCTATGGCAAGCATTGAAGGTAAA-3′ E.coli(pET-28a) reverse 5′-CTCGAGTTAGATTCGTTTATATGTTCGAAC-3′
Derf15 forward 5′-GGATCCATGAAAACCATATATGCAATACTT-3′ E.coli(pET-28a) reverse 5′-CTCGAGTTATTCGCCTATACAAGTCAATT-3′
Derf18 forward 5′-AGATCTATGACTCGATTCTCTTTGACTG-3 E.coli(pET-28a) reverse 5′-GTCGACTTAACAACCGGGTATTTTGGTATG-3′
Derf20 forward 5′-GGATCCATGGTCGATCAAGCTACCCTGA-3′ E.coli(pET-28b) reverse 5′-GTCGACTTACAATGATTTTTCCATTTTAATCAATTCA-3′
Table2.Listofcloning primersequences(continued)
Allergen Primer Direction
PrimerSequence Expression
Host(Vector) Derf21 forward 5′-GGATCCATGAAATTCATTATTTTCTGTGCCA-3′ E.coli(pET-28a)
reverse 5′-CTCGAGTTAATCATCCGATTTTACAGCTTTCACCTT-3′
Derf21 forward 5′-CTCGAGAAAAGAGAGGCTGAAGCTTTTATTGTTGGTGACAAAAAAGAAGAT GAATG-3′ P.pastoris(pPIC9) reverse 5′-GCGGCCGCTTAATGATGATGATGATGGTGATCATCCG ATTTTACAGCTTTC-3′
Derf23 forward 5′-CTCGAGAAAAGAGAGGCTGAAGCTGATATTGATCATGATGATGATCC-3′ P.pastoris(pPIC9) reverse 5′-GCGGCCGCTTAATGGTGATGATGGTGATGTGTACATGTTAATTCTTTTTC ATTC-3′
Tyrp5 forward 5′-GGATCCATGAAGTTCG CCATTCTCGC-3′ P.pastoris(pPIC9) reverse 5′-CTCGAGTTAGCGAGTCTTGACAGCCT-3’
Tyrp21 forward 5′-GGATCCATGAAGTTCGCCATCGTTCTTATTG-3′ E.coli(pET-28a) reverse 5′-CTCGAGTTATTGGGTTTGAATATCCTTCACTTTTTG-3′
Blot5 forward 5′-GGATCCATGAAGTTCGTCATCGCCCT-3’ E.coli(pET-28a) reverse 5′-GTCGACTTAGACCT TGATGGCGTTCACT-3′
Blot21 forward 5′-GGATCCATGAAATTTATCATCGCATTGGCTG-3′ E.coli(pET-28a) reverse 5′-GTCGACTTATTCGGAATCTTGGACTCGCTTT-3′
Derp23 forward 5′-CTCGAGAAAAGAGAGGCTGAAGCTGCCAATGATAATGATGATGATC-3′ P.pastoris(pPIC9) reverse 5′-GCGGCCGCTTAATGGTGATGATGGTGATGAGTGCATGTTTCTTCATCTTC A-3′
I
I
I
.RESULTS
1.IgE reactivity profileofKorean mite-sensitizedallergicpatientsto recombinantD.farinaeallergens
1)IgE reactivity toD.farinae,T.putrescentiaeandB.tropicalis
extracts
IgE reactivities of three species of mite extract (D. farinae, T. putrescentiae, B.tropicalis) were tested using ELISA to mite-sensitized patients’ as a preliminary experiments. Extracts of D. farinae, T. putrescentiae,B.tropicaliswerereacted with 194of194(100%),110of194 (56.7%)and126of194(64.9%)housedustmite-sensitizedpatientssera(Fig. 1).Thereactivity againstD.farinaehaslow correlation with theothers,but
T.putrescentiaeandB.tropicalisshowedhigh correlation with eachother(r = 0.953)(Fig.2).This resultshowed thatallergens from T.putrescentiae
Fig.1.IgE reactivities to D.farinae (Derf),T.putrescentiae (Tyrp) andB.tropicalis(Blot)crudeextracts.IgE reactivitieswereevaluatedby ELISA analysis,using serafrom 194HDM sensitizedpatientsand 20normal control sera.Bold lines indicate the average value of positive reactions. Dottedlineindicatethecut-offvalues.
Fig. 2. Correlations among the IgE reactivities to D. farinae, T. putrescentiaeandB.tropicaliscrudeextracts.
2)PreparationofnativeandrecombinantDerf4
(1)cDNA andproteinsequenceanalysisofDerf4
Previous studies reported an importance of group 4 mite allergen in sensitization orIgE reactivity.A full-length cDNA sequenceofDerf4and recombinant Der f4 production,however,have not been reported.A full cDNA sequenceofDerf4wasclonedinthisworkforthefirsttime.
TheclonedDerf4cDNA was1,901bpin length containing a91bpof 5’non-translating region,open reading frame coding for a mature protein with 525aminoaicdswith astopcodon TAA atnucleotidepositions1,576– 1,578.Itspredicted molecularweightwasestimated to beapproximately 60.5 kDa(Fig.3).EightdifferentcDNA sequencesofDerf4wereidentified(Fig. 4)
A protein sequence homology search was performed using the Advance Blastsearch.Deduced amino acid sequence ofDerf4 has 88,86,65 and 66% sequencehomology with thegroup4allergensofD.pteronyssinus(Der p4),E.maynei(Eurm 4),T.putrescentiae(Tyrp4)andB.tropicalis(Blo t4),respectively(Fig.5).
Fig.3.The nucleotide and deduced amino acid sequences ofDer f4. Stop codon is indicated by 3 asterisks.Its estimated molecular mass and isoelectricalpointare60.46kDaand6.11.
Fig.5.Amino acids sequence alignmentof5 mite group 4 allergens: Der f 4 (Gene bank accession number KM016828), Der p 4 (D. pteronyssinus,accession number AAD38942.1,identity 88%),Eur m 4 (E. maynei,AAD38943.1,86%),Tyrp 4 (T.putrescentiae,ABM53754.1,65%) Blo t4 (B.tropicalis,AAQ24543.1,66%).The N terminus ofthe mature enzymeislabeled+1andtheaminoacidnumbering isbasedontheDerf4 sequence.UnderlinesindicatepredictedN-glycosylationsites.
(2)ExpressionandenzymaticanalysisofrDerf4
The rDer f 4 produced from E. coli and P. pastoris resolved at approximately 61.2and 58.5kDaon an SDS-PAGE gel,respectively (Fig.6). nDerf4 was purified to homogeneity from the whole body extractofD.
farinae using an optimized procedure for affinity precipitation with oyster glycogen.ThenDerf4migratedasasinglebandof58kDaonSDS-PAGE (Fig.6,D).N-terminalamino acid sequence (DSKFSNPHFIG)ofnDerf4 identified by Edman sequencing (data notshown)corresponded to theamino acidsequencededucedfrom Derf4cDNA (Fig.3).
Toevaluatebiochemicalnature,α-amylaseactivity ofnDerf4and rDer f4weretestedusing acommercialamylaseassay kit.P.pastorisexpressed rDerf4 had α-amylase activity (36.9 mU/nmol)similarto nDerf4 (38.5 mU/nmol) (Table 3),whereas rDer f 4 from E.coli had no α-amylase activity (Fig.7).This resultimplies thatrDerf4 expressed in P.pastoris
Fig.6.Vectormap and SDS-PAGE analysisofnativeand recombinant Derf4.Expressionvectordiagram ofDerf4forE.coli(A)andP.patoris
(B).SDS-PAGE ofrDerf4(C)andnDerf4(D).M,molecularsizemarker; E,rDerf4E.coli–expressed form;P,rDerf4P.patoris–expressed form; CE,D.farinaecrudeextract;N,nDerf4.
Fig.7.Amylaseactivity ofD.farinae crudeextract,nDerf4 (purified Derf4),andrDerf4(producedin P.pastoris).
Table 3.α-amylase activity ofD.farinae crude extract,nDerf4,and rDerf4. Sample Name D.farinae crude extract nDerf4 rDerf4 (producedin P. pastoris) rDerf4 (producedin E. coli) Activity (mU/㎍) 0.04 0.71 0.63 N.D. Activity (mU/nmol) 38.5 36.9
Unitdefinition :Oneunitofα-amylaseistheamountofα-amylasethat cleavesethylidene-pNP-G7togenerate1.0μmolofnitrophenolpermin at pH 7.20at25˚C.
(3)AllergenicpropertiesofnativeandrecombinantDerf4
IgE reactivity to both nDer f 4 and rDer f 4 was demonstrated by Western blotwith poolofD.farinae-sensitized sera (Fig.8).Only single bandwasobtainedforeachtestedallergen.While146ofthe194sera(75.3%) testedwerefoundtobereactivetonDerf4and62(32.0%)reactedtorDer f4 (Fig.9).Thereisno corelation ofIgE reactivity between nDerf4 and rDerf4. Additionally,thecorelation ofIgE reactivity with α-amylasefrom other species, German cockroach (Blattella germanica). B. germanica α
-amylase showed a 6.2% (12/194)ofIgE reactivity showing no correlation (Fig.10).Theseresultsshowed thatDerf4 arehighly reactiveallergen in Koreanmiteallergysubject,notcross-reactivetocockroachα-amylase,Blag 11.
Fig.8.SDS-PAGE and Western blotting ofnativeand recombinantDer f 4. (A) SDS-PAGE of native and recombinant Der f 4.Proteins were separated underreducing conditionson 10% polyacrylamide gels and stained with Coomassiebrilliantblue.(B)Immunoblotting ofnativeand recombinant Derf4 with Derfsensitized patients sera.M,molecularmass marker;N, nDerf4;P,rDerf4(expressedinP.pastoris).
Fig.9.IgE reactivity ofD.farinae-sensitizedhuman sera(A)tonDerf 4 and rDerf4 (expressed in P.pastoris)and its correlation (B).The cut-offvalues(dashedlines)weredeterminedasthemeanplustwo‑foldthe standarddeviationofthecontrolseravalues.
Fig.10.IgE reactivity of D.farinae-sensitized human sera (A) and correlations (B)to nDer f4 and German cockroach (B.germanica)α
-amylase.The cut-offvalues (dashed lines)were determined as the mean plustwo‑foldthestandarddeviationofthecontrolseravalues.
3)IgE reactivitiesandcorrelationsofthirteenD.farinaeallergens
(1)Expressionandpurificationofrecombinantallergensfrom D.farinae
forIgE reactivitytest
Thirteen differentallergensfrom D.farinae wereproduced using E.coli
orP.pastorisexpressionsystem.Derf1,Derf3,Derf4,Derf5,Derf21 andDerf23wereproducedinP.pastoris,andtheothers(Derf2,Derf7, Derf10,Derf13,Derf15,Derf18and Derf20)in E.coli.They were purifiedwithanaffinitychromatography,Ni-sepharoseresin(Fig.11).
Fig. 11. SDS-PAGE profile of recombinant D. farinae allergens. Coomassie-stained10% (rDerf4),12% (rDerf15and18)15% (rDerf1,2, 3,5,7,10,13,20and21)and16.5% (rDerf23)SDS-PAGE gelcontaininga molecularweightmarkerandeachpurifiedrecombinantallergen.
(2)MeasurementofIgE reactivitiesto13D.farinaeallergens
IgE reactivitiesto purified D.farinae allergenswereexamined by using ELISA.rDerf1,3and23producedfrom P.pastorisexpressionsystem,and theothersfrom E.coliwereexamined.However,nDerf4wasused,since rDerf4showsalow reactivitytoIgE ofallergicsera.
D.farinae extractwasused as a positivecontrol.While173 ofthe194 sera(89.2%)testedwerefoundtobeIgE reactivetorDerf1,187(96.4%)to rDerf2,120 (61.9%)to rDerf3,147 (75.8%)o nDerf4,126(64.95%)to rDerf5,85(43.8%)torDerf7,48(24.7%)torDerf10,7(3.6%)torDerf 13,8(4.1%)torDerf15,62(32.0%)torDerf18,75(38.7%)torDerf20, 128 (66.0%) to rDer f 21,and 84 (43.3%) to rDer f 23 (Fig.12).IgE reactivity to D.farinae extractwas highly correlated with those to major allergen,Derf1(r=0.649),Derf2(r=0.721)and Derf3(r=0.528)(Fig.13). TheseresultssuggestthatDerf1andDerf2arevery importantallergens in diagnosisofmiteallergy,followed by Derf3,Derf4,Derf5,Derf7, Derf21andDerf23.
Fig.12.IgE reactivities to recombinant D.farinae allergens.IgE reactivities were evaluated by ELISA analysis,usingserafrom 194HDM sensitizedpatientsand20normalcontrolsera.
(3)CorrelationofIgE reactivitiesandclinicalsymptoms
IgE reactivity of D.farinae allergens versus clinical symptoms was investigated.Serafrom asthmaandrhinitispatientswerehighly IgE reactive to various D.farinae allergens.Der f1 showed a high IgE reactivity to various clinicalsymptoms,exceptcough.Der f2 reacted to allsera from asthma and rhinitis showing a highestreactivity againstsera with allergy. Derf3,Derf4,Derf5,Derf21andDerf23reactedto40% ormorein asthma and rhinitis patient’s sera (Fig.14,Table 4).These results suggest thatmajormite allergens,Derf1 and Derf2,have a highly sensitization rate in mite allergic asthma and rhinitis patients,butrelationship ofmite allergenandclinicalsymptomsisunclear.
Table4.PrevalenceagainstD.farinaeallergensperclinicaldiagnosis.
Allergens Ast(n=61)hma RhiAllernigitics (n=73) Asthma+ Allergic Rhinitis (n=17) Atopic dermatitis (n=7) Cough (n=5) Ur(tn=8)icaria Other diagnosis (n=23) Derf1 98.4 84.9 88.2 100.0 60.0 87.5 82.6 Derf2 100.0 94.5 100.0 100.0 80.0 87.5 95.7 Derf3 63.9 60.3 52.9 71.4 40.0 75.0 60.9 Derf4 77.0 71.2 76.5 85.7 80.0 62.5 82.6 Derf5 75.4 57.5 58.8 71.4 60.0 50.0 69.6 Derf7 39.3 47.9 47.1 57.1 60.0 12.5 43.5 Derf10 29.5 24.7 29.4 42.9 20.0 12.5 13.0 Derf13 1.6 4.1 5.9 0.0 0.0 0.0 8.7 Derf15 3.3 4.1 5.9 14.3 0.0 0.0 4.3 Derf18 32.8 26.0 47.1 14.3 60.0 12.5 39.1 Derf20 37.7 43.8 35.3 28.6 60.0 25.0 30.4 Derf21 72.1 57.5 70.6 71.4 80.0 25.0 69.6 Derf23 34.4 49.3 58.8 42.9 60.0 37.5 34.8
*Other diagnosis:conjunctivitis,tonsillopharyngitis,angioedema,drug allergy and notdiagnosed.
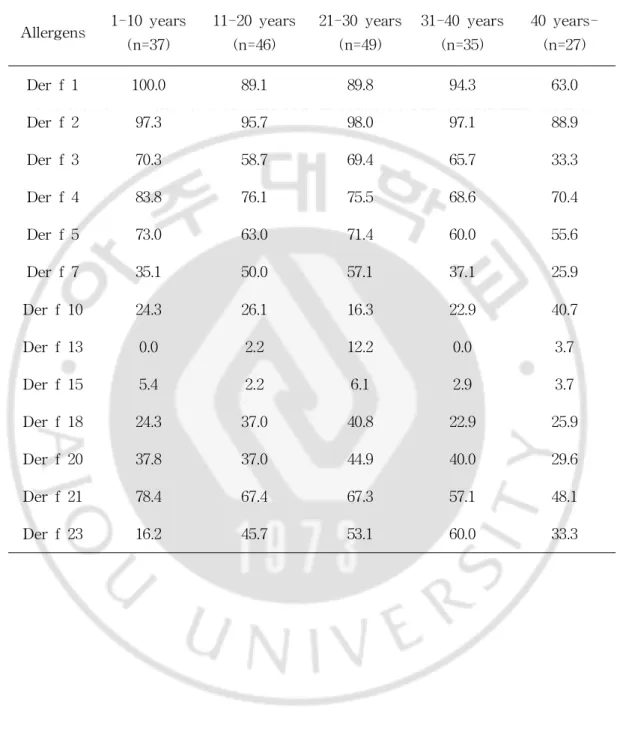
(4)IgE reactivitiescomparedwithasubject’sageandanalysisof sensitizationnumber
A relationship ofbetween IgE reactivity ofD.farinae allergensand age wasexamined.Derf1showeda100% IgE reactivity undertheageof10in mite allergy,and Der f 2 reacted 90% or more regardless of their ages (Fig.15,Table5).Asaresult,sensitizationtoeachallergenwasfoundnotto berelatedwiththeirages.
Mostofpatient’s sera (190 of194,97.9%)reacted to three ormore D.
farinae allergens.The two sera (2.1%)recognized to single allergen.These subjects were found to be sensitized with Der f4 only (Fig.16).Three subjectsreacted with twelvedifferentallergens.Onewasnotreacted rDerf 13 and theotherswererDerf15 only.Theseresultssuggestthatmostof patients with mite allergy in Korea were found sensitized with multiple allergens.
Table5.PrevalenceagainstD.farinaeallergensperage. Allergens 1-10years (n=37) 11-20years (n=46) 21-30years (n=49) 31-40years (n=35) 40year s-(n=27) Derf1 100.0 89.1 89.8 94.3 63.0 Derf2 97.3 95.7 98.0 97.1 88.9 Derf3 70.3 58.7 69.4 65.7 33.3 Derf4 83.8 76.1 75.5 68.6 70.4 Derf5 73.0 63.0 71.4 60.0 55.6 Derf7 35.1 50.0 57.1 37.1 25.9 Derf10 24.3 26.1 16.3 22.9 40.7 Derf13 0.0 2.2 12.2 0.0 3.7 Derf15 5.4 2.2 6.1 2.9 3.7 Derf18 24.3 37.0 40.8 22.9 25.9 Derf20 37.8 37.0 44.9 40.0 29.6 Derf21 78.4 67.4 67.3 57.1 48.1 Derf23 16.2 45.7 53.1 60.0 33.3
2.Evaluation ofcross-reactivity among miteallergens
1)Cross-reactivitiesofsixgroup5and21miteallergens
(1)SequenceanalysisofDerf5,Derf21,Tyrp5,Tyrp21,Blot5, andBlot21
Theaminoacidsequencesofthesixgroup5andgroup21allergens,Der f5,Derf21,Tyrp5,Tyrp21,Blot5,andBlot21,werecompared.As shown in Table 6,each allergen exhibits between 36.6 and 55.8% sequence identity to the othergroup 5 and 21 allergens.While Derf5 exhibited the highestlevelofidentitytoBlot5(44.4%),Derf21wasmostsimilartoTyr p5(41.6%),Tyrp5wasmostsimilartoBlot5(52.9%),andTyrp21was mostsimilartoBlo t21(55.8%).Thelowestlevelofidentity observed was betweenDerf5andBlot21(36.6%),whilethehighestwasbetweenTyrp 21 and Blo t 21 (55.8%) (Table 6).This high levelofsequence identity between allergenssuggeststherewould beacross-reactivity between group 5andgroup21allergens.Asdepictedin Fig.17,fouraminoacidsthatwere previously found to be crucialforIgE binding,areconserved in fiveofthe allergens:Der f21 (E-77,D-82,E-87,and E-92),Tyr p 5 (E-78,D-83, E-88,and E-93),Tyr p 21 (E-82,D-87,E-92,and E-97)Blo t5 (E-76, D-81,E-86,and E-91),and Blot21(E-74,D-79,E-84,and E-89).Derf5 (L-74,D-79,E-84,andE-89)wastheonlyallergeninwhichthebindingsite was notconserved (Fig.17).These results show thatgroup 5 and 21 mite allergenshaveahighsimilaraminoacidssequence,andcross-reactivitiesare expectedamongtheseallergens.
Fig.17.Sequencecomparison ofrecombinantDerf5and othergroup5 and group 21allergens.SequencealignmentofDerf5with Derf21,Tyr p 5,Tyrp 21,Blo t5 and Blo t21.Fourofthe charged residues were previously identified asthecrucialamino acidsforIgE reactivity in Blo t5 (marked by the asterisk *).The signalpeptide regions,predicted by the SignalP 4.1Server(http://www.cbs.dtu.dk/services/SignalP/),areunderlined.
Table 6.Sequence identity ofgroup 5 and group 21 allergens from D.
farinae,T.putrescentiae,andB.tropicalis. Aminoacid
sequence
identity (%) Derf5 Tyrp5 Blot5 Derf21 Tyrp21 Blot21
Derf5 100 38.0 44.4 36.7 40.3 36.6 Tyrp5 - 100 52.9 41.6 45.3 43.4 Blot5 - - 100 41.2 37.4 40.3 Derf21 - - - 100 39.3 37.5 Tyrp21 - - - - 100 55.8 Blot21 - - - 100
(2)Productionofrecombinantgroup5andgroup21allergens
RecombinantDerf5,Derf21,Tyrp5,Tyrp21,Blot5,andBlot21 were expressed in E.coli.The recombinantproteins were purified from E.
coliinclusion bodies by affinity chromatography using Ni-sepharose resin. Purified proteins were separated on 15% SDS-PAGE and visualized by CoomassieBrilliantBluestaining.Asexpected,thepurified proteinsmigrated slightly slower than that 15 kDa marker,which was consistent with the predicted17kDasizeoftheallergens(Fig.18).
Fig.18.SDS-PAGE ofrDerf5,rDerf21,rTyrp 5,rTyp p 21,rBlot 5 and rBlo t 21. Coomassie-stained 15% SDS-PAGE gel containing a molecularweightmarker(laneM)and3μgofpurifiedrecombinantallergens.
(3)AnalysisofIgE reactivitytorecombinantgroup5andgroup21 allergens
The IgE reactivity ofeach purified recombinantallergen was examined by ELISA,using serafrom 194Korean HDM allergicpatients.Derf2was used as a positive control.While 187 ofthe 194 sera (96.4%)tested were found to reactto rDerf2,126 (65.0%)reacted to rDerf5,128 (66.0%)to rDerf21,59 (30.4%)to rTyrp 5,80 (41.2%)to rTyrp 21,60 (30.9%)to rBlot5,and 42(21.7%)torBlot21(Fig.19).TheIgE reactivity ofrDerf 5alsocorrelatedstronglywiththatofrDerf21(r=0.733).TheIgE reaction torTyrp5exhibitedthehighestcorrelation withtheIgE reaction torTyrp 21 (r = 0.950).Italso correlated moderately with those ofrDer f5 (r = 0.563),rDerf21 (r= 0.613)and Blo t5(r= 0.742).TheIgE reactivity of rTyrp21correlatedthehighestwith thereactivitiesofrDerf5(r= 0.571), rDerf21(r=0.651)andBlot5(r=0.704),whilethereactivity ofrBlot5 correlated highly with those ofrTyrp 5 (r= 0.742),and rTyrp 21 (r= 0.704).However,theIgE reactivity ofrBlo t21 exhibited a poorcorrelation with thoseofrDerf5,rDerf21,rTyrp5,and rTyrp21(r= 0.270,0.305, 0.254,0.349,respectively)(Fig.20).These results demonstrate thatthe IgE reactivity ofallsix recombinantallergensexhibited atleastsomecorrelation tooneanother.
Fig. 19. The IgE reactivities to six recombinant group 5 and 21 allergens.They were evaluated by ELISA using sera from 194 house dust mite-sensitized patients and 20 normalcontrolsera.Each dotindicates an individualserum.The cut‑offvalues (dashed lines)were setas the mean plustwo‑foldthestandarddeviationofthecontrolsera.
Figure 20. Correlations between the IgE reactivities of recombinant allergens.A totalof194serum sampleswereusedin thedirecthuman IgE ELISA.
(4)AnalysisofIgE cross-reactivityamongrecombinantgroup5and group21allergens
In orderto investigatecross-reactivity,IgE inhibition wasperformed on eachallergen,using acompetitiveELISA approachandserum pooledfrom 10 allergy patients(Table7).Recombinantsix allergenswerenotblockedaIgE binding to mite extracts (Fig.21).Perhaps it is means that mite crude extractcontains a very low quantity ofgroup 5 and 21 allergens.The IgE reactivitiesagainstrDerf2,rDerf5and rDerf21 wereonly inhibited by selfsameallergens.rDerf21wasfound tobethestrongestinhibitorofIgE reactiontotheotherallergens,inhibiting 93.1% ofIgE reactivity torTyrp5, 92.1% torTyrp21,87.0% torBlot5,and70.3% torBlot21,ataninhibitor concentration of10 µg/mL (Fig.22).The IgE reactivity to rBlo t21 was inhibited upto35.3% by rDerf5,and up to43.6% by rTyrp21.rTyrp5 inhibited 76.7% ofIgE reactivity to rTyr p 21,while rTyr p 21 inhibited 86.0% ofthe IgE reaction to rTyrp 5.The IgE reactivity ofBlo t5 was inhibitedupto77.4% byrTyrp5and73.4% byrTyrp21.Additionally,rBlo t21wasinhibitedupto43.6% ofreactivitytorTyrp21.Inpredictionof3D structure,six allergenshaveavery similarform,and they haveaconserved structureofmajorepitopesites(Fig.23).Theseresultssuggestthateach of theallergenstestedexhibitssomedegreeofcross-reactivitytooneanother.
Table7.Characteristicsofpatients’serausedin thecompetitiveELISA No. Age (year) Sex (M/F ) Clinical symptoms ELISA results(A450nm)a
rDerf5 rDerf21 rTyrp5 rTyrp21 rBlot5 rBlot21
1 23 M rhinitis 2.916 3.281 2.739 3.251 0.239 0.723 2 14 M asthma 3.029 3.991 1.898 2.035 1.899 0.146 3 31 M rhinitis 2.128 2.285 0.420 1.544 0.071 1.519 4 25 F rhinitis 2.560 3.252 0.402 1.114 0.078 1.602 5 55 M asthma 2.343 1.804 1.404 1.641 0.800 0.074 6 9 M asthma 2.866 3.168 1.932 2.483 0.703 2.855 7 16 M asthma 2.559 3.113 2.084 2.495 0.785 0.071 8 12 M asthma 2.336 2.994 1.588 1.909 1.377 0.104 9 24 M rhinitis 2.838 2.999 1.798 2.214 1.884 0.090 10 16 M atopic dermatitis 2.623 3.289 2.190 2.672 1.578 0.087 a
The cut-offvalues were 0.070 forrDerf5,0.125 forrDerf21,0.172 for rTyrp 5,0.240 forrTyrp 21,0.086 forrBlo t5 and 0.080 forrBlo t21, respectively
Fig.21.CompetitiveIgE inhibition ELISA ofgroup5and21allergen to crude extract. It was conducted using pooled sera from 10 patients, pre-absorbedwith0.001,0.01,0.1,1,or100μg/mL ofeachallergen.
Fig.22.Inhibition analysesofsix recombinantallergens.CompetitiveIgE inhibition ELISA was conducted using pooled sera from 10 patients, pre-absorbed with 0.001,0.01,0.1,1,or100μg/mlofeach allergen (■:rDer f2,◆:rDerf5,◇:rDerf21,▲:rTyrp5,△:rTyrp21,●:rBlot5,○: rBlot21).
Fig. 23. Predicted 3D-structure of recombinant group 5 and 21 allergens.Themoleculeiscolored redforresiduesthatarepredictedasthe crucialaminoacidsforIgE reactivity.EssentialsitesofIgE binding displayed aredballandthinstick.
2)Cross-reactivitybetweenDerf23andDerp23
(1)ProductionofrDerf23andrDerp23
RecombinantDerf23and Derp23wereexpressed in P.pastoris.The recombinantproteins were purified from P.pastoriscultured supernatantby affinity chromatography. Purified proteins were then separated by 16.5% SDS-PAGE andvisualizedby CoomassieBrilliantBluestaining.Asexpected, thepurifiedproteinsmigratednear10kDamarker,whichwasconsistentwith thepredicted9.5kDasizeoftheallergens(Fig24).
Fig.24.Sequencecomparison and SDS-PAGE ofrecombinantDerf23 and Der p 23.(A)Sequence alignmentofDerf23 with Derp 23.The signalpeptide regions are underlined.(B)Coomassie‑stained 10-15% (left) SDS-PAGE and 16% (right)tricine SDS-PAGE gelcontaining a molecular weightmarker(laneM)and3μgofpurifiedrecombinantallergens.
(2)IgE cross-reactivitybetweenrecombinantDerf23andDerp23
IgE inhibition on D.farinaecrudeextract,rDerf23andrDerp23were performed with a pooled sera from 10 mite allergy patients using a competitive ELISA (Table 8).IgE reactivities against Der f extract were inhibited only by itself,and Derfextractwas notinhibited IgE binding to rDerf23and rDerp 23.rDerp 23wasfound tobethestrongestinhibitor ofIgE reaction totherDerf23,inhibiting 86.1% ofIgE reactivity torDerf 23,atan inhibitorconcentration of10µg/ml(Fig.25).IgE binding torDerp 23wasblocked(61.1%)byrDerf23at10µg/ml.Theseresultsshowedthat thereiscross-reactivitybetweenrDerf23andrDerp23..
Table 8. Characteristics of patients’ sera used in the competitive ELISA between rDerf23andrDerp23
No. Age (year) Sex (M/F) Clinical symptoms ELISA results(A450nm)a Derf
extract rDerf22 rDerp23 1 26 M Cough 1.848 0.675 0.740 2 20 M rhinitis 2.354 2.269 1.805 3 49 F Asthma 2.710 0.579 0.993 4 36 M Cough 2.841 0.876 0.752 5 20 M Asthma 2.032 0.808 2.292 6 63 M Urrhitinicartiisa, 2.286 0.984 1.747 7 19 F Atopic Dematitis 2.803 0.584 0.66 8 29 F DeAtmatopiictis 2.204 0.974 0.869 9 23 F DeAtmatopiictis 3.189 1.943 1.737 10 24 M rhinitis 2.374 1.291 1.631 aCut-offvalues were 0.125 forD.farinae extract,0.060 forrDerf23 and
Fig.25.Inhibition ELISA of rDer f 23 and rDer p 23.IgE binding capacity in D.farinae extract(A),rDerf23 (B)and rDerp 23 (C)was inhibited with a pooled sera pre-absorbed with a various quantity of D.
3. Production of proinflammatory cytokines, IL-6 and IL-8, in BEAS-2B cellsby stimulation ofrecombinantallergensderivedfrom HDM,D.farinae.
To determineifthe allergenscan induce a pro-inflammatory response in respiratory epithelial cells, secretion of mediators in 24 h cell culture supernatants from a BEAS-2B cells, was initially analysed by ELISA. Stimulation with mite extract showed that extract of D.farinae and D. pteronyssinus were effectively induced a production of IL-6 and IL-8 in BEAS-2B cells,but T.putrecentiae extract did not (Fig.26,27).After exposure to rDerf4 (Fig.28),rDerf21 (Fig.29)and rDerf5 (Fig.31) increased levels ofIL-6 and IL-8 from BEAS-2B cells,whereas rDerf10, rDerf13andrDerf23werelittleeffectonthecells(Fig.32).
BEAS-2B cells were cultured with rDer f 21 with/without additional anti-TLR2,anti-TLR4antibodiesandTLR4antagonistfor24hours.Although rDerf21 aloneinduced a secretion ofIL-6 and IL-8 in BEAS-2B cells,it wasconcentration-dependent,in comparison to medium controls(Fig.29).In the presence ofanti-TLR4 and TLR4 antagonist,IL-6 and IL-8 secretion wasreduced in comparison tothecontrol.Theseresultsshowed thatrDerf 21 stimulate the production of the IL-6 and IL-8 in BEAS-2B,and this reactionthroughaTLR4signalpathway.
Fig. 26. SDS-PAGE profile of D.farinae, D.pteronyssinus and T. putrecentiae crude extracts.The extracts (20 μg of D.farinae extract (DF),D.pteronyssinusextract(DP)and T.putrecentiae extract(TP))were runon10-25% SDS-PAGE gelandstainedwithCoomassieblue.
Fig.27.Three species mite extracts-induced CCL20,IL-6 and IL-8 secretion from BEAS-2B cells.BEAS-2B cells were incubated for 24 h withmedium aloneorwithD.farinae, D.pteronyssinusandT.putrescentiae
Fig.28.rDerf4-induced IL-6 and -8 secretion from BEAS-2B cells. BEAS-2B cellswereincubated for24 h with medium aloneorwith 10and 50 ug/mlrDer f4.As positive control,cells were also activated with D.
Fig. 29. rDer f 21-induced IL-6 (A) and IL-8 (B) secretion from BEAS-2B cells.BEAS-2B cellswereincubatedfor24hwithmedium alone orwith 100ug/mlrDerf21.Asapositivecontrol,cellswerealsoactivated with Pam3CSK4 (50 ng/ml) and LPS (100 ng/ml). Cells were also pre-incubated with a blocking anti-TLR2 antibody,anti-TLR4 antibody or isotypecontrolbeforerDerf21.IL-6andIL–8ELISA wasusedtoquantify levelsofIL-6andIL-8productioninculturesupernatants.
Fig. 30. rDer f 21-induced IL-6 (A) and IL-8 (B) secretion from BEAS-2B cells with TLR4 antagonist.BEAS-2B cellswereincubated for 24 h with medium alone or with 100 ug/mlrDer f 21.Cells were also pre-incubated with LPS-RS (TLR4 antagonist,100 ng/ml)before rDerf21. IL-6 and IL-8 ELISA wasused to quantify the levelsofIL-6 and IL-8 in culturesupernatant.
Fig. 31. rDer f 5-induced IL-6 (A) and IL–8 (B) secretion from BEAS-2B cells.BEAS-2B cellswereincubatedfor24hwithmedium alone orwith 100ug/mlrDerf5.Asa positivecontrol,cellswerealso activated with Pam3CSK4 (50 ng/ml) and LPS (100 ng/ml). Cells were also pre-incubated with blocking anti-TLR4 antibody before rDerf5.IL-6 and IL-8 ELISA was used to quantify levels ofIL-6 and IL-8 production in culturesupernatant.
Fig.32.rDerf10,rDerf13,rDerf23and rDerp23-inducedIL-6(A) and IL-8 (B)secretion in BEAS-2B cells.BEAS-2B cellswereincubated for24h with medium aloneorwith 100 ug/mlrecombinantallergens.Asa positive control,cells were also activated with Pam3CSK4 (50 ng/ml) and LPS (100ng/ml).IL-6and IL-8-specificELISA wasused toquantify levels ofIL-6andIL-8productioninculturesupernatant.