Article
http://dx.doi.org/10.4217/OPR.2017.39.2.125 June 2017
동해 울진 원전 온배수배출구 주변 해역에서의 대형저서동물 군집구조
권순현 1,2 · 유옥환 1,2* · 이형곤 1
1한국해양과학기술원 생태기반연구센터 (15627) 경기도 안산시 상록구 해안로 787
2한국해양대학교 해양과학기술전문대학원 (49112) 부산광역시 영도구 태종로 727
Community Structure of Macrobenthos around the Thermal Discharge Area of the Uljin Nuclear Power Plant in the East Sea, Korea
Soon Hyun Kwon 1,2 , Ok Hwan Yu 1,2* , and Hyung-Gon Lee 1
1Marine Ecosystem and Biological Research Center, KIOST Ansan 15627, Korea
2Ocean Science and Technology School, Korea Maritime University Busan 49112, Korea
Abstract : This study analyzed the community structure of macrobenthic organisms in the subtidal area suffering under the influence of thermal discharge from the Uljin nuclear power plant during 2012 −2013 and reviewed the temporal change in the faunal composition of the macrobenthic community using data from previous studies since the start of the plant operation in 1989. A total of 319 species were collected in 2012−2013, with a mean density of 3,330 ind./m
2and a mean biomass of 131.96 g/m
2. These results were similar to those obtained in 2006−2007. The dominant species were not different from each other before and after the power plant operations began, but the faunal density near the power plant drainage port was significantly higher compared to those densities obtained from other stations. Spiophanes bombyx and Polydora sp., which is known as a pollution indicator species, was dominant only in the drainage port area.
The study area consisted of two subregions with different macrobenthic communities depending on the water depth, which was similar to the results of the 2006−2007 survey. The macrobenthic community structures before and after the operation of the power plant in the coastal area were not changed, but there were seasonal differences in the long-term macrobenthic community structure which were strongly related to the thermal discharge from the power plant.
Key words : macrobenthos, community structure, long-term variation, thermal discharge, Uljin Nuclear Power Plant, East Sea
1. 서 론
우리나라 연안에 건설되어 있는 원자력발전소는 발전에 이용하지 못한 폐열을 재활용하기 위하여 연간 수백억톤
이상의 해수를 복수기에 유입시킨 후, 배수구를 통하여 취 수구의 수온보다 평균 7
oC 이상의 온배수를 방출하고 있 다(KORDI 2007). 더군다나 최근의 전력수요 증가와 함께 연안에 많은 발전소가 건설 중이거나 계획 중에 있어 온 배수의 방출량은 점차 증가할 것으로 여겨진다. 온배수 배 출량 증가는 해양생물의 서식에 중요한 인자로 작용하고
*Corresponding author. E-mail : [email protected]
있는 해수온의 변화에 영향을 주어 온배수의 영향이 미치 는 지역에서의 해양생태계는 주변 지역에서의 해양생태계 와 차이가 커질 것으로 여겨진다(Barnett 1972; Laws 1981). 대형저서동물은 부유생물에 비해 생활사 기간 동안 서식 공간 이동이 거의 없으며, 외부의 환경변화에 회피할 수 있는 능력이 거의 없는 생활 방식을 가지기 때문에, 온 배수 배출에 의한 해양생태계 변화를 파악하기 위해 대형 저서동물을 이용한 연구가 많이 진행되어졌다(Bamber and Spencer 1984; Bamber 1990; Lardicci et al. 1999;
Riera et al. 2011).
대형저서동물 군집은 크게 물리적인 요인과 생물적 요 인에 영향을 받는다(Mclennan and Miller 1990; Smythe and Sawyko 2000; Bedulina et al. 2010; Teixeira et al.
2012). 물리적 요인에는 저층의 수심, 수온, 퇴적상 등이 있으며, 생물적 요인에는 서식 공간이나 먹이에 대한 종 내/종간 경쟁과 포식자 등이 있다(Gray 1981). 대형저서동 물의 공간 분포는 물리적 요인에 의해 결정되며, 특히 퇴 적상은 대형저서동물의 공간 분포를 결정하는 중요한 요 인으로 알려져 있다(Buchanan et al. 1978; Weston 1988;
Hoey et al. 2004). 더군다나, 연안에서의 인위적인 교란은 여러 요인과 함께 대형저서동물의 군집구조를 복잡하게 변화시킬 수 있다(Pearson and Rosenberg 1978; Gray 1981; Hartley 1982; Bamber and Spencer 1984). 또한 인 위적인 교란이 지속되어지면, 대형저서동물의 군집구조에 영향을 주는 요인들은 일반적으로 현상과 다른 패턴을 보 일 수 있다(Pearson and Rosenberg 1978; Bamber and Spencer 1984).
대형저서동물의 군집구조는 온배수에 개체수의 증감, 우점종의 변화 등 영향을 받는 것으로 여겨지고 있다 (Snoeijis and Prentice 1989; Suresh et al. 1993; Kailasam and Sivakami 2004). 그러나 일부 해역에서의 자연적 환 경 변화 폭이 온배수에 의한 변화 폭보다 크기 때문에, 대 형저서동물의 군집이 온배수의 영향을 받는지 파악하기 어렵다고 알려져 있다(Lardicci et al. 1999). 온배수 지역 근처에서는 급변한 환경변화로 인하여 기회주의 종들이 우점하는 것으로 알려지고 있다(Mahadevan 1980; Bamber and Spencer 1984). 그러나 일부 연구에서는 기회주의 종 들이 전혀 출현하지 않았다(Lardicci et al. 1999).
동해 연안의 해안선은 남북 방향으로 단조롭거나 암반 과 사질 해빈이 반복적으로 나타나며, 수심 경사가 급격히 나타나고 있는 특징을 보인다(Kim et al. 2001). 이와 같 은 환경 특징으로 인하여, 동해안의 대형저서동물 군집은 크게 수심과 퇴적상의 영향을 받는 것으로 알려지고 있다 (Paik et al. 2007; Yu et al. 2011). 동해안 울진 원자력발 전소는 1988년에 1호기 운전을 시작으로, 2005년에 6호 기를 가동하여, 현재까지 6개호기에서 온배수가 지속적으
로 방출되고 있다(KHNP 2009). 울진 원자력발전소에서 나오는 온배수는 해안선을 따라 남북방향으로 흐르기 때 문에, 온배수에 의한 저서동물의 변화는 온배수의 영향이 있는 얕은 수심에서 나타나고 있다(Yu et al. 2011, 2013).
또한 온배수 배출 부근 지역에서는 온수종과 기회종의 특 성을 동시에 가지는 대형저서동물이 높게 나타나고 있으 며(Yu et al. 2011), 발전소 정비 등의 이유로 각 호기 별 가동 정지가 있을시 대형저서동물의 군집구조 변화는 단 기간 동안 있는 것으로 알려져 있다(Yu et al. 2013). 또한 발전소 온배수의 배출이 소규모 환경 교란을 일으켜 대형 저서동물의 군집 조성에 변화를 주는 것으로 보고되었다 (Choi et al. 2000). 지금까지 우리나라 온배수 지역에서의 대형저서동물 군집 연구는 강릉, 월성, 기장, 울진 등의 지 역에서 단기간에 걸쳐 이루어졌다(Choi et al. 2000; Seo et al. 2009; Kim et al. 2011; Yu et al. 2011, 2013). 더군 다나 온배수 영향이 큰 지역에서 지속적인 온배수가 장기 간 방류되었을 때, 대형저서동물의 종 조성과 군집구조가 어떻게 변하는 지에 대한 장기 변동 연구는 매우 제한적 이다.
이러한 관점에서 본 조사는 동해 연안의 원자력발전소 인근 해역에서 발전소 가동 10년 이후의 대형저서동물의 종 조성과 군집구조에 변화가 있는지를 분석하는데 초점 을 두었고, 발전소 가동 전과 가동 이후의 대형저서동물의 종 조성과 군집구조의 변화를 분석하였다.
2. 재료 및 방법
생물채집
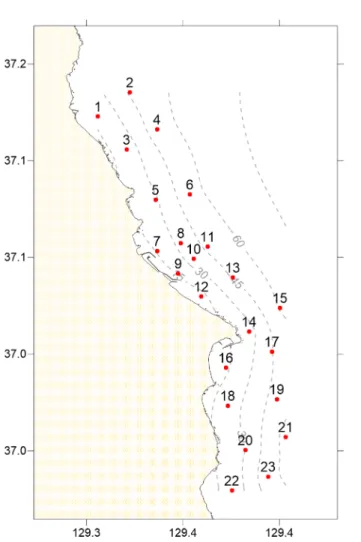
울진원자력발전소 주변에 서식하는 대형저서동물의 분 포특성을 파악하기 위해 2012년 8월과 2013년 2월 조사 를 실시하였다(Fig. 1). 본 연구의 조사 정점은 원자력발전 소를 중심으로 등 간격으로 설정된 23개의 연성기질 조하 대를 대상으로 하였다(Table 1). 울진 원자력 발전소 주변 의 해양생태계는 발전소를 기준으로 북쪽으로 5.5 km까 지 남쪽으로 9.7 km까지 발전소 온배수의 영향을 받는다 고 알려져 있다(MOF 2015). 그러므로 본 연구 지역은 일 부 정점(정점 1, 2, 3, 4, 22, 23)을 제외한 대부분의 조사 정점은 원자력 발전소의 온배수 영향을 받는 지역 안에 위치하였다(MOF 2015). 조사 정점은 수심 평균 30 m 보 다 낮은 지역과 깊은 지역으로 구분하여 울진 원자력발전 소를 중심으로 반경 11 km 내에 등 간격으로 위치하였으 며, 평균 수심은 35 m였다. Smith-McIntyre grab(면적:
0.1 m
2)를 사용하여 채집하였으며, 각 정점 당 2회씩 퇴적
물을 인양하였다. 인양된 퇴적물은 선상에서 그물코(Mesh
size) 1 mm 의 체(Sieve)를 사용하여 걸렀으며, 체에 걸린
모든 퇴적물은 10% 해수중성포르말린으로 고정하여 실험
실로 운반하였다. 고정된 시료는 실험실에서 생물체를 분 리한 후 동물 분류군별로 선별하였으며, 해부현미경하에 서 동정·계수하였다. 출현한 모든 대형저서동물은 종 수준 까지 동정하였으며, 명확한 동정이 어려우나 다른 종임을 구분할 수 있는 것들은 코드를 붙여 각기 다른 종임을 표 시하였다.
환경분석
환경요인의 측정은 대형저서동물의 채집과 동일한 시기 와 정점에서 이루어졌으며, 퇴적물온도계(SATO PC- 9200) 를 통해 퇴적물온도를 측정하였고, CTD(SBE-19)를 사용하여 수심, 저층 수온, 저층 염분, 저층 용존산소량 (DO)을 측정하였다. 퇴적물 내의 총 유기탄소량(TOC)과 퇴적물입도 분석을 위해 채니기에 인양된 퇴적물을 각 정 점 당 50 g을 채취한 후 냉동 보관하여 실험실로 운반하 였다. 운반된 퇴적물은 50
oC 에서 48시간 건조시킨 후 1 g 씩 채취하여 10%의 과산화수소로 유기물을 0.1 N 염산으
로 탄산염을 제거한 후 CHN 분석기를 통해 총유기탄소 량을 구하였다. 퇴적물입도분석을 위해서는 습식체질을 통해 조립질과 세립질로 분리한 다음 4 ø 이상의 세립질 은 입도 분석기(Sedigraph 5100D)를 통해 입자별 비율을 구하였다.
군집분석
각 정점으로부터 얻어진 대형저서동물의 종별·개체수 자료는 단위 면적당(m
2)으로 환산하여 사용하였다. 출현 하는 저서동물의 특성을 파악하기 위해 단변량분석인 종 다양도 지수(Shannon and Wiener 1963)를 구하였으며, 정점별 종조성과 그에 따른 시·공간적 유사도를 측정하기 위하여 군집분석을 실시하였다. 대형저서동물의 군집구조 는 발전소 4개호기 가동 2년 후(2001)와 6개호기 가동 (2005) 후로 구분하여 분석하였다. 발전소 가동에 따른 대 형저서동물 군집구조는 2001년 KEPRI에서 조사된 연구 보고서 및 한국해양연구원의 자료를 재분석하였다(KEPRI 2001; KORDI 2008). 발전소 6개호기 가동 후의 대형저서 동물 군집구조는 가동 1년 후의 자료와 본 연구에서 분석 Fig. 1. Map showing the sampling stations and bathymetry
in this study. The drainage port is located in near st. 9
Table 1. Sampling location and depth
Site Longituge Latitude Depth(m)
1 129.35600 37.17280 17.0
2 129.37250 37.18530 46.8
3 129.37080 37.15570 20.7
4 129.38660 37.16610 53.5
5 129.38580 37.12970 28.0
6 129.40360 37.13250 55.2
7 129.38660 37.10320 15.6
8 129.39895 37.10740 34.7
9 129.39758 37.09160 15.3
10 129.40550 37.09920 38.1
11 129.41277 37.10547 47.0
12 129.40950 37.07970 10.0
13 129.42578 37.08953 46.7
14 129.43430 37.06160 23.8
15 129.45028 37.07382 61.7
16 129.42223 37.04298 12.6
17 129.44610 37.05130 44.6
18 129.42320 37.02330 23.1
19 129.44863 37.02667 51.0
20 129.43238 37.00035 30.0
21 129.45330 37.00713 65.0
22 129.42537 36.97945 19.0
23 129.44420 36.98672 56.0
된 자료를 통합하여 분석하였다(KEPRI 2002; Yu et al.
2011). 대형저서동물의 군집분석에서는 생물자료간의 편 중을 피하기 위해 모든 자료는 4th root로 변환하였으며, Bray-Curtis 유사도를 이용한 집괴분석(Cluster analysis)을 통해 대형저서동물군집의 분포특성을 파악하였다(Clarke and Ainsworth 1993). 집괴분석 시 파악된 조사 정점 간의 유사도는 SIMPROF (Similarity Profile Test) 분석을 통해 통계적 유의수준 5% 내에서의 최대 유사 정점을 선별하 였으며, 수지도(dendrogram) 혹은 MDS(non-Metric multi- Dimensional Scaling) 배열법을 통해 표기하였다. 또한 구 분된 저서동물군집의 유의성 검증과 각 정점군내의 정점 간 유사도 및 정점군간의 비유사도에 영향을 미치는 기 여종을 파악하기 위해 군집분석 통계패키지인 PRIMER (ver. 6)를 이용한 SIMPER(Similarity Percentages Procedure) 분석을 실시하였으며, 분석 시 기여도가 높은 상위 5종을 기여종으로 선정하였다. 대형저서동물의 공간 분포에 영향을 끼치는 주요 환경요인을 파악하기 위해서 는 다변량분석에서 사용된 Bray-Curtis 유사도지수와 환 경변수와의 순위상관계수를 통한 BIO-ENV 분석을 실시 하였으며, 분석 시 Draftsman plot 분석을 통해 95% 이상 의 상관관계를 가지는 중복 환경변수는 제외하였다 (Clarke and Ainsworth 1993). 또한 조사된 환경 요인 및 생물요인 간의 상관관계를 파악하기 위해 Spearman rank correlation 분석을 실시하였다.
3. 결 과
저서해양환경
조사 지역의 저층 염분도는 평균 33.51 psu로 정점 간 큰 차이가 나지 않았으며, 조사 기간 동안에도 차이가 없 었다(Fig. 2). 반면에, 저층 수온은 2012년 8월에 평균 12.0
oC 였으나, 수심이 낮은 연안에 위치한 정점에서의 수 온은 13.42−23.08
oC의 분포를 보였고, 배수구 아래쪽에 위치한 정점 12에서 23.08
oC로 가장 높았다. 그러나 2013 년 2월의 저층 수온은 7.45−9.61
oC( 평균 9.14
oC) 로 대부분의 정점에서의 수온은 일정하였다. 수심이 가장 낮 은 정점은 배수구 아래쪽에 위치한 정점 12의 10 m였으 며, 가장 깊은 수심은 정점 21의 65 m였다.
조사 기간 동안 퇴적물의 입도 분포는 −0.09−6.24 ø사 이였으며, 평균 입도는 3.35 ø였다(Fig. 2). 퇴적물의 입도 는 수심이 30 m 보다 낮은 지역에서 대부분 3 ø 이하의 조립한 입도 분포를 보였으나, 수심이 30 m 보다 깊은 지 역에서는 3 ø 이상의 세립한 입도 분포를 보였다(KrusKal- Wallis test H = 18.2, p < 0.001). 각 정점 별 퇴적물 입도 분포는 조사기간 동안 대부분의 정점에서 큰 차이가 없었 지만 배수구 앞에 위치한 정점 9와 정점 13에서의 퇴적물
입도가 2012년 8월에 각각 2.73 ø와 2.54 ø를 보였지만 2013년 2월에는 각각 −0.09 ø와 0.08 ø로 자갈의 함량이 급격히 증가하였다. 이는 조사 기간 동안 신울진원자력발 전소 심층 배수로 공사 작업과 관련이 높은 것으로 여겨 진다.
조사 기간 동안 퇴적물에 존재하는 총유기탄소의 양은 0.05−1.79% 범위로 대부분의 정점에서 비슷하였으며, 평 균 0.69%를 보였다. 총유기탄소의 양은 수심이 깊은 정점 에서 비교적 1% 이상의 값을 보였으나, 수심이 낮은 정점 에서는 1% 이하의 값을 보였다. 특히 퇴적물의 분급도가 높은 지역에서 유기물의 함량이 높았으나 정점 13의 경 우, 분급도가 2.49로 높음에도 유기물의 함량은 0.48%로 낮았다(Fig. 2).
대형저서동물
분류군 별 종수와 개체수와 생체량
조사 기간 동안 출현한 대형저서동물의 총 종수는 319 종이었으며, 평균 출현 개체수는 3,330 개체/m
2였고, 평균 생체량은 131.96 g/m
2이었다. 대형저서동물의 총 출현 종 수는 2012년 8월에 253종이었으나, 2013년 2월에는 229 종으로 감소하였다. 대형저서동물의 주요 분류군 가운데 다모류 종 수가 106종으로 가장 많이 출현하였으며, 갑각 류, 연체동물 순으로 감소하였다. 다모류의 출현 종수는 대부분의 정점에서 50% 이상을 차지하였다. 갑각류와 연 체동물의 출현종수는 2012년 8월에서 2013년 2월로 갈수 록 차지하는 비율이 증가하였으며, 특히 배수구 바로 앞의 연체동물의 출현 종수는 두 기간 동안 매우 낮았다.
대형저서동물의 평균 개체수는 2012년 8월에 4,895 개 체/m
2이었으나, 2013년 2월에는 1,768 개체/m
2로 반 이하 로 감소하였다. 대형저서동물의 평균 생체량은 2012년 8월에 178.7 g/m
2이었으나, 2013년 2월에는 85.2 g/m
2로 반 이하로 감소하였다. 대형저서동물의 주요 분류군의 출현 생체량은 성구동물이 2012년 8월과 2013년 2월에 각각 76.3 g/m
2(55%) 와 30.2 g/m
2(35%) 로 전체 출현 생체량에 서 가장 높았다. 조사 지역의 대부분에서 극피동물과 연체 동물에 의한 생체량 비율이 높았으나, 수심이 깊은 곳에서 는 극피동물에 의한 생체량 비율이 증가하였다. 2012년 8월 배수구 앞에 위치한 정점 9에서의 생체량은 2012년 8 월에는 연체동물이 90% 이상을 차지하였으며, 2013년 2월에는 갑각류가 60% 이상을 차지하였고, 다모류의 생 체량 비율은 매우 낮았다.
시·공간 분포
2012년 8월 정점 간 출현하는 대형저서동물의 출현 종
수는 23−78종 사이였으며, 평균 48종이 출현하였다. 배수
구 인근에 위치한 정점 9와 정점 12에서 각각 24종과 23
종으로 가장 낮았으며, 발전소에서 멀어질수록 출현 종수 는 증가하였다(Fig. 3). 또한 대형저서동물의 출현 종수는 수심이 30 m 이하에 위치한 연안 정점에서는 평균 42종 이 출현하였으나, 30 m 이상의 깊은 수심에 위치한 정점
에서는 평균 55종으로 종수는 증가하였다.
대형저서동물의 출현 개체수는 1,115−16,465 개체/m
2분포를 보였으며, 배수구 인근에 위치한 정점 12에서
16,465 개체/m
2로 가장 높았으며, 배수구 앞인 정점 9에서
Fig. 2. Environmental conditions at each station in the period of 2012 − 2013
13,410 개체/m
2로 두 번째로 높았다. 그러나 배수구에서 동쪽으로 가장 수심이 깊은 곳에 위치한 정점 11에서의 개체수는 1,115 개체/m
2로 가장 낮았다. 대형저서동물의 개체수는 수심이 낮은 정점에서 높았으며, 수심이 깊어질 수록 감소하는 경향을 보였다. 평균 수심이 30 m 이상인 깊은 지역에 위치한 정점들의 평균 개체수는 2,180 개체/
m
2이상을 차지하였으나, 평균 수심이 30 m 이하인 낮은 지역에 위치한 정점들의 평균 개체수는 7,851 개체/m
2이 상으로 수심이 깊은 지역보다 약 3배 이상 높았다.
대형저서동물의 생체량은 정점 간 8.8−629.7 g/m
2사 이를 보였으며, 정점 14에서 가장 낮았고, 정점 10에서 가 장 높았다. 특히 출현 개체수가 10,000 개체/m
2이상 출현 하였던 정점 가운데 정점 1과 정점 12에서의 생체량은 각 각 24.6 g/m
2과 38.1 g/m
2로 낮았다. 대형저서동물의 생체 량은 극피동물이 높게 출현한 수심이 깊은 정점에서 높았 다. 수심 30 m를 기준으로, 낮은 수심에서의 생체량은 104 g/m
2이었으나, 깊은 수심에서는 247 g/m
2로 2배 이상 높았다.
대형저서동물의 종 다양도 지수는 평균 2.12이였으며, 정점 간 0.27−3.45 분포를 보였다. 종 다양도 지수 값이 1 이하를 나타내는 지역은 발전소 주변 연안에 위치한 정점 이었다. 배수구 앞에 위치한 정점 9에서 대형저서동물의 종 다양도 지수는 0.27로 가장 낮았으며, 배수구 인근 남
측에 위치한 정점 12에서 0.36으로 두 번째로 낮았다. 또 한, 취수구 앞에 위치한 정점 7에서의 종 다양도 지수는 0.61 이었다. 반면에 발전소에서 가장 동쪽에 위치한 정점 11에서 종 다양도 지수는 3.45로 가장 높았다. 종 다양도 지수는 수심이 깊은 정점에서 비교적 높았다. 수심 30 m 를 기준으로, 수심이 낮은 지역에 비해 수심이 깊은 지역 에서의 종 다양도 지수는 약 3배 이상 높았다(Fig. 3).
2013 년 2월 대형저서동물의 출현 종수는 정점 간 16−
60 종 사이였으며, 평균 42종이 출현하였다. 배수구 인근에 위치한 정점 9와 정점 12에서 각각 17종과 16종으로 가장 낮았으며, 수심이 낮은 곳에 위치한 정점 가운데 발전소에 서 멀어질수록 출현 종수는 증가하는 경향을 보였다(Fig.
3). 대형저서동물의 출현 종수는 수심이 30 m 이하에 위 치한 연안 정점에서는 평균 30종이 출현하였으나, 30 m 이상의 깊은 수심에 위치한 정점에서는 평균 52종으로 2012년 8월과 비슷한 경향을 보였다.
2013년 2월의 대형저서동물의 출현 밀도는 730−3,510 개체/m
2분포를 보였으며, 배수구 앞에 위치한 정점 9에서 730 개체/m
2로 가장 낮았고, 배수구 인근 동남측에 위치 한 정점 13에서의 개체수는 890 개체/m
2로 주변 지역에 비해 낮았다. 대형저서동물의 개체수는 평균 수심이 30 m 이상인 깊은 지역에 위치한 정점들의 평균 개체수는 1,738 개체/m
2이상을 차지하였으나, 평균 수심이 30 m
Fig. 3. Number of species, density, biomass and diversity of macrobenthos at each station in the study period of 2012 −
2013
이하인 낮은 지역에 위치한 정점들의 평균 개체수는 1,802 개체/m
2이상으로, 수심간 큰 차이를 보이지 않았다 (Fig. 3). 그러나 발전소를 기준으로 북측에 위치한 정점들 은 수심이 낮은 정점에서 개체수가 높았으나, 발전소 남측 에 위치한 정점에서는 수심이 깊은 정점들에서의 개체수 가 높았다.
2013년 2월의 대형저서동물의 생체량은 정점 간 4.7−
377.5 g/m
2사이를 보였으며, 정점 18에서 가장 낮았으 며, 정점 2에서 가장 높았다. 대형저서동물의 생체량은 극 피동물이 높게 출현한 수심이 깊은 정점에서 높았다. 수심 30 m 를 기준으로, 낮은 수심에서의 생체량은 26.1 g/m
2이 었으나, 깊은 수심에서는 139 g/m
2로 약 4배 이상 높았다.
2013년 2월의 대형저서동물의 종 다양도 지수는 평균 2.41이였으며, 정점 간 0.63−3.46 분포를 보였다. 종 다양 도 지수 값이 1 이하를 나타내는 지역은 발전소 취수구 앞에 위치한 정점 7(0.63)이었다. 또한 발전소 배수구 인 근에 위치한 정점 9와 정점 12에서 각각 1.60과 1.03으로 다른 정점에 비해 상대적으로 낮은 종 다양도 값을 보였 다. 종 다양도 지수는 수심이 깊은 정점에서 비교적 높았 다. 수심 30 m를 기준으로, 수심이 낮은 지역은 1.76이었 으나, 수심이 깊은 지역에서의 종 다양도 지수는 3.02로 약 2배 이상 높았다(Fig. 3).
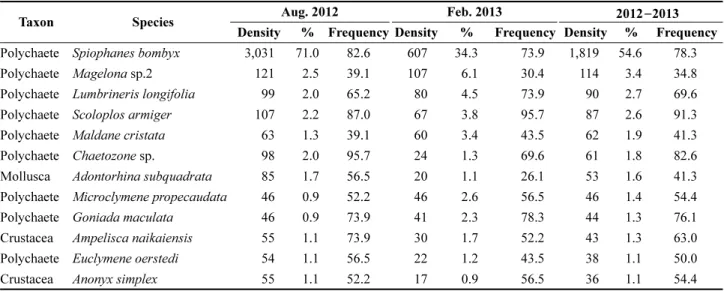
우점종
조사 기간 동안 총 출현 개체수의 1% 이상을 차지한 우점종은 총 12종이었다(Table 2). 가장 우점한 종은 다모 류 Spiophanes bombyx(민얼굴갯지렁이)로 평균 1,819 개 체/m
2가 출현하였으며, 총 출현 개체수의 54.6%를 차지하 였고, 78.3%의 출현율을 보였다. 두 번째로 우점한 종은 다모류인 양손갯지렁이류(Magelona sp.2)로 평균 114 개
체/m
2(3.4%) 가 출현하였으나, 출현율은 34.8%로 일부 정 점에서만 출현하였다. 세 번째로 우점한 종은 다모류 Lumbrineris longifolia(긴자락송곳갯지렁이, = Scolotoma longifolia) 로 평균 90 개체/m
2가 출현하였으며, 총 출현량 의 2.7%를 차지하였다. 네 번째로 우점한 종은 Scoloplos armiger(삼각모자갯지렁이)로 평균 87 개체/m
2(2.6%)가 출현하였으며, 출현율은 91.3%로 대부분의 정점에서 출 현하였다. 나머지 주요 우점종들 가운데 다모류에는 Maldane cristiata(민숭대나무갯지렁이), Chaetozone sp.
(실타래갯지렁이류), Microclymene propecaudata(꼬마대 나무갯지렁이), Euclymene oerstedi, Goniada maculata(작 은갈매기고리갯지렁이) 등이 있으며, 이매패류에 속하는 Adontorhina subquadrata( 꼬마말발조개)와 단각류에 속하 는 Ampelisca naikaiensis(나이카안경옆새우), Anonyx simplex( 긴팔옆새우류) 등이 우점종이었다. 가장 우점한 다모류 민얼굴갯지렁이는 2012년 8월 평균 3,031 개체/
m
2가 출현하였으며, 총 출현 개체수의 62.0%를 차지하였 고, 82.6%의 출현율을 보였다. 그러나 2013년 2월의 경우, 평균 607 개체/m
2가 출현하여, 여름철에 비해 5배 이상 감소하였다. 반면에 두 번째로 우점한 다모류 양손갯지렁 이류(Magelona sp.2)로 2012년 8월에 평균 121 개체/m
2(2.5%) 가 출현하였고, 2013년 2월에 107 개체/m
2(6.0%) 가 출현하여, 두 기간 동안 개체수의 차이는 거의 없었다. 다 모류 삼각모자갯지렁이는 2012년 8월에 평균 107 개체/
m
2(2.2%) 가 출현하였으나, 2013년 2월에 67 개체/m
2(3.8%) 로 개체수는 감소하였으나, 출현 빈도율은 87.0%에 서 95.7%로 증가하였다. 나머지 우점종들 또한, 2012년 8월에서 2013년 2월로 갈수록 개체수가 감소하는 경향을 보였다(Table 2).
조사기간 동안 우점종들의 정점 별 출현양상을 보면,
Table 2. Dominant species of macrobenthic communities in the study area during 2012 −2013
Taxon Species Aug. 2012 Feb. 2013 2012−2013
Density % Frequency Density % Frequency Density % Frequency
Polychaete Spiophanes bombyx 3,031 71.0 82.6 607 34.3 73.9 1,819 54.6 78.3
Polychaete Magelona sp.2 121 2.5 39.1 107 6.1 30.4 114 3.4 34.8
Polychaete Lumbrineris longifolia 99 2.0 65.2 80 4.5 73.9 90 2.7 69.6
Polychaete Scoloplos armiger 107 2.2 87.0 67 3.8 95.7 87 2.6 91.3
Polychaete Maldane cristata 63 1.3 39.1 60 3.4 43.5 62 1.9 41.3
Polychaete Chaetozone sp. 98 2.0 95.7 24 1.3 69.6 61 1.8 82.6
Mollusca Adontorhina subquadrata 85 1.7 56.5 20 1.1 26.1 53 1.6 41.3
Polychaete Microclymene propecaudata 46 0.9 52.2 46 2.6 56.5 46 1.4 54.4
Polychaete Goniada maculata 46 0.9 73.9 41 2.3 78.3 44 1.3 76.1
Crustacea Ampelisca naikaiensis 55 1.1 73.9 30 1.7 52.2 43 1.3 63.0
Polychaete Euclymene oerstedi 54 1.1 56.5 22 1.2 43.5 38 1.1 50.0
Crustacea Anonyx simplex 55 1.1 52.2 17 0.9 56.5 36 1.1 54.4
최우점종인 민얼굴갯지렁이는 주로 수심이 낮은 연안에서 출현 개체수가 높았으며, 특히 2012년 8월 배수구 앞에 위치한 정점 9와 정점 12에서 각각 12,860 개체/m
2과 15,405 개체/m
2로 극우점하였다(Fig. 4). 2013년 2월 정점 9와 정점 12에서의 민얼굴갯지렁이 개체수는 각각 430과 780 으로 급격히 감소하였지만, 정점 1과 정점 5, 정점 7에 서의 개체수는 1,120−3,100으로 높은 개체수를 유지하였 다. 두 번째로 우점한 양손갯지렁이류는 수심이 깊은 정점
2, 4, 6, 15, 23에 주로 출현하였으며, 출현 개체수는 조사
시기별 차이를 보였다, 양손갯지렁이는 정점 6에서
2012 년 8월(905)에서 2013년 2월(25)로 갈수록 개체수가
급격히 감소하였으나, 정점 2와 4에서는 3배 이상 증가하
였다. 세 번째로 우점한 긴자락송곳갯지렁이는 발전소 인
근의 수심이 낮은 정점을 제외한 대부분의 정점에서 개체
수가 출현하였다. 2012년 8월에 비해 2013년 2월에는 수
심이 낮은 연안 정점에서 개체수가 증가하였지만, 발전소
Fig. 4. Dominant species of macrobenthos at each station in the study period of 2012 − 2013
취수구 앞에 위치한 정점 7과 배수구 앞과 남측에 위치한 정점 9, 정점 12, 정점 14에서는 2012년 8월과 마찬가지 로 전혀 출현하지 않았다. 다모류 삼각모자갯지렁이와 실 타래갯지렁이류(Chaetozone sp.)는 수심과 상관없이 대부 분의 정점에서 출현하였으며, 삼각모자갯지렁이의 개체수 는 수심이 깊은 정점에서 증가하였으나, 실타래갯지렁이 류는 수심이 낮은 곳에서 증가하는 경향을 보였다. 다모류 민숭대나무갯지렁이는 수심이 깊은 정점에서 출현하였으 며, 수심이 낮은 정점에서는 전혀 출현하지 않았다. 다모 류에 속하는 꼬마대나무갯지렁이, E. oerstedi, 작은갈매기 고리갯지렁이 등은 대부분 수심이 깊은 정점에서 주로 출 현하였으나, 수심이 낮은 정점에서는 거의 출현하지 않았 다. 조사 시기별 개체수 차이는 크지 않았다. 꼬마말발조
개는 발전소 북측의 수심이 깊은 정점에서 2012년 8월에 높은 개체수가 출현하였으나, 2013년 2월에는 거의 출현 하지 않았다. 단각류에 속하는 나이카안경옆새우는 수심 에 상관없이 대부분의 정점에서 출현하였으나, 긴팔옆새 우류(A. simplex)는 수심이 낮은 지역에서만 출현하였다.
단각류 2종은 배수구 앞에 위치한 정점 9와 정점 12에서 는 거의 출현하지 않았다(Fig. 4). 오염지시종으로 알려진 다모류 긴얼굴갯지렁이류(Polydora sp.)는 정점 13번에서 주로 출현하였으며, 2012년 8월(65 개체/m
2) 보다 2013년 2월(105 개체/m
2)에 약 2배 이상 증가하였다.
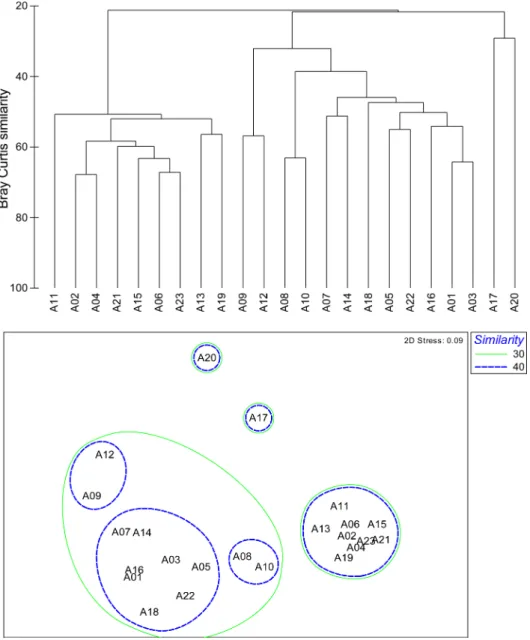
군집구조
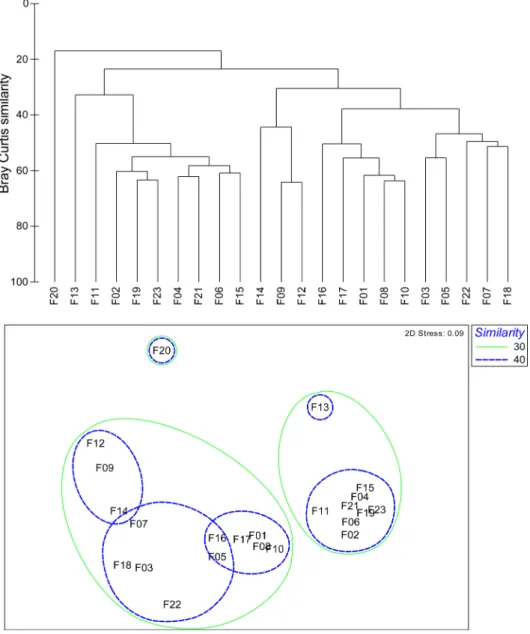
2012년 8월 조사 정점 간 종별 개체수 자료의 유사성에
Fig. 5. Dendrogram and 2-dimensional plot of stations result from the cluster analysis and MDS ordination using the
macrobenthic faunal abundance data, respectively in August 2012
근거한 집괴분석과 MDS 분석 결과, 총 23개의 정점은 유 사도 약 30%를 기준으로 크게 2개의 정점군 I과 II로 구 분되었다(Fig. 5). 수심 30 m를 기준으로 연안에 위치한 정점군 I 정점과 외해의 정점군 II로 구성되었다. 정점군 I 그룹은 유사도 약 40%를 기준으로, 정점군 I-1(정점 9와 정점 12), I-2(정점 8과 정점 10) 그리고 나머지 정점인 I- 3으로 구분되었다. 대형저서동물의 군집구조에 영향을 미 치는 주요 환경요인은 평균입도, 수심, 저층 수온으로 분 석되었으며(Bio-Env; Rho = 0.845, p < 0.01), 연안정점들 로 구성된 정점군 I는 정점군 II에 비해 높은 퇴적물온도 와 모래함량 그리고 낮은 총유기탄소량의 저서환경특성을 보였고, 생물학적특성으로는 상대적으로 낮은 종수와 종 다양도와 생체량을 보였으나, 개체수는 상대적으로 높았
다(Table 3). 정점군 I 가운데, 정점군 I-1은 발전소 배수구 근처에 위치한 정점으로, 평균 수심은 12.7 m, 수온은 18.2
oC 를 보여, 정점군 I보다 평균 0.7이상 높았다. 반면 에 정점군 I-2는 평균 수심이 36.4 m로 수심이 30 m 이 상인 정점들로, 정점군 I과 환경특성의 차이를 보였다.
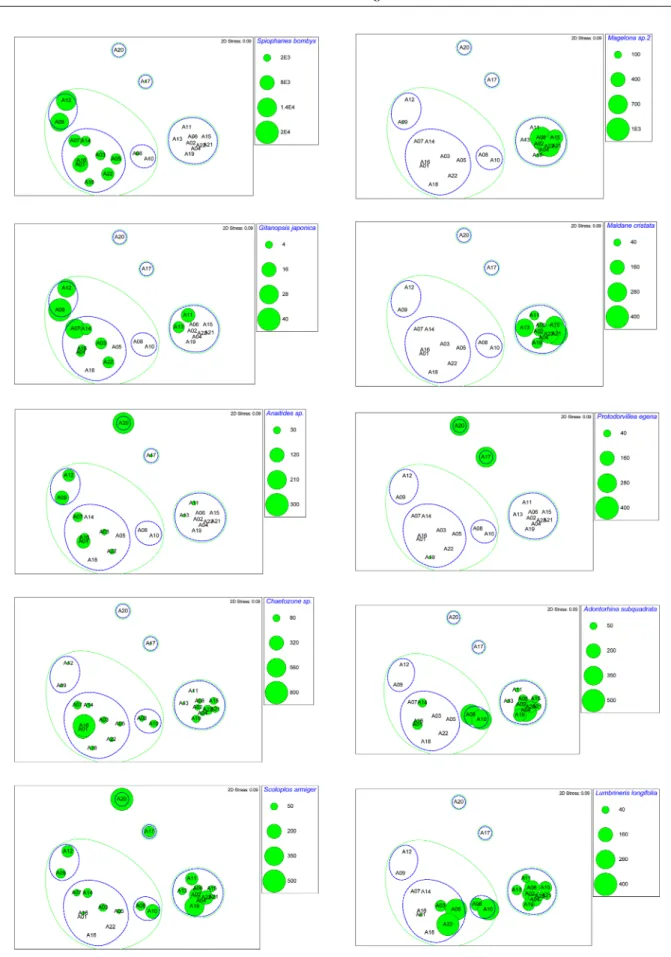
SIMPER 분석결과 정점군 I에서는 민얼굴갯지렁이와 실타 래갯지렁이류(Chaetozone sp.) 그리고 단각류 Synchelidium lenorostralum( 납작뿔붙은눈옆새우), 나이카안경옆새우, 긴팔옆새우류(A. simplex)가 정점군에 속한 정점간의 유사 도에 영향을 주는 주요 기여종이었으며, 정점군 II에서는 긴자락송곳갯지렁이(L. longifolia), 민숭대나무갯지렁이, 작은갈매기고리갯지렁이, 삼각모자갯지렁이와 꼬마말발조 개(A. subquadrata)가 기여종이였다(Fig. 6). 한편, 정점군 Table 3. Ecological character of same macrobenthic structure in August 2012
I
(st. 1, 3, 5, 7, 8, 9, 10, 12, 14, 16, 18, 22)
I-1 (st. 9, 12)
I-2 (st. 8, 10)
I-3 (st. 1, 3, 5, 7, 14, 16, 18, 22)
II (st. 2, 4, 6, 11, 13,
15, 19, 21, 23)
III (st. 17, 20)
Depth 19.6 12.7 36.4 20.0 53.7 37.3
SAND 95.6 94.5 85.6 95.7 54.3 97.7
SILT 2.4 3.5 8.6 2.4 29.0 0.7
CLAY 1.8 1.9 5.8 1.9 16.5 0.8
MEAN 2.5 2.3 3.3 2.8 4.8 0.7
SORTING 0.6 0.9 1.3 0.6 2.7 0.6
TN 0.0 0.0 0.0 0.0 0.1 0.1
TOC 0.3 0.1 0.3 0.2 1.1 1.5
Temp sediment 17.5 18.2 8.7 17.2 7.8 15.4
Salinity in bottom 33.4 33.3 33.6 33.4 33.3 33.5
Temp in bottom 17.8 19.0 8.9 18.1 5.9 11.0
Oxygen in bottom 7.9 8.0 8.9 7.9 8.6 7.9
S 41.6 23.5 54.5 47.4 56.1 38.0
N 7851.4 14937.5 2567.5 6695.0 2170.6 2207.5
H'(loge) 1.2 0.3 3.0 1.3 3.1 2.5
biomass 103.8 117.7 326.4 67.2 229.8 308.1
Depth 19.6 12.7 36.4 20.0 53.7 37.3
Spiophanes bombyx 6273.6 14132.5 265.0 5091.9 7.2 60.0
Magelona sp.2 0.5 2.5 0.0 0.0 308.3 0.0
Lumbrineris longifolia 77.3 0.0 240.0 106.3 105.0 0.0
Scoloplos armiger 84.5 115.0 135.0 26.9 127.2 302.5
Maldane cristata 0.0 0.0 0.0 0.0 161.7 0.0
Chaetozone sp. 145.0 10.0 72.5 196.9 57.2 2.5
Adontorhina subquadrata 14.5 0.0 395.0 20.0 111.1 0.0
Microclymene propecaudata 2.7 0.0 7.5 3.8 112.2 0.0
Goniada maculata 3.2 0.0 52.5 3.1 100.6 10.0
Ampelisca naikaiensis 73.2 0.0 195.0 100.6 5.6 5.0
Euclymene oerstedi 41.4 0.0 30.0 56.9 81.1 0.0
Anonyx simplex 106.8 7.5 47.5 138.8 0.0 25.0
Fig. 6. MDS ordination plots and the spatial distribution of 8 characteristic species emerged by the SIMPER analysis
in August 2012
I 와 정점군 II간의 비유사도에 기여하는 종은 민얼굴갯지 렁이, 꼬마대나무갯지렁이(M. propecaudata), 양손갯지렁 이류(Magelona sp.2)였다. 정점군 I 내의 I-1과 I-3간의 비 유사도에 기여하는 종은 갑각류들로, I-1에는 거의 출현하 지 않았다.
2013년 2월 조사 정점 간 종별 개체수 자료의 유사성에 근거한 집괴분석과 MDS 분석 결과, 총 23개의 정점은 유 사도 약 30%를 기준으로 크게 2개의 정점군 I과 II로 구 분되었다(Fig. 7). 수심 30 m를 기준으로 연안에 위치한 정점군 I 정점과 외해의 정점군 II로 구성되었다. 정점군 I 그룹은 유사도 약 40%를 기준으로, 정점군 I-1(정점 9, 12, 14), I-2( 정점 1, 8, 10, 16, 17) 그리고 나머지 정점인
I-3 으로 구분되었다. 대형저서동물의 군집구조에 영향을 미치는 주요 환경요인은 평균 입도, 수심으로 분석되었으 며(Bio-Env; Rho = 0.8, p < 0.01), 연안정점들로 구성된 정점군 I는 정점군 II에 비해 낮은 수심과 평균입도가 조 립하였으며, 생물학적 특성으로는 상대적으로 낮은 종수 와 종 다양도와 생체량을 보였으나, 개체수는 상대적으로 높았다(Table 4). 정점군 I 가운데, 정점군 I-1은 발전소 배 수구 근처에 위치한 정점으로, 평균 수심은 16.4 m로, 2012년 8월에 비해 배수구에서부터 남쪽으로 정점 14까지 확장된 곳까지 동일 그룹으로 묶였으며, 이 지역에서의 출 현 종수와 개체수는 정점군 I 가운데 가장 낮은 값을 보였 다. 반면에 정점군 I-2는 평균 수심이 29.6 m로 수심이
Fig. 7. Dendrogram and 2-dimensional plot of stations result from the cluster analysis and MDS ordination using the
macrobenthic faunal abundance data, respectively in February 2013
상대적으로 깊었으며, 종수와 출현 개체수가 다른 정점 군 I보다 높은 특징을 보였다. SIMPER 분석결과 정점군 I에서는 민얼굴갯지렁이와 삼각모자갯지렁이, 단각류 Grandifoxus bangpoensis( 민가시긴뿔옆새우)가 정점군에 속한 정점간의 유사도에 영향을 주는 주요 기여종이었으 며, 정점군 II에 기여한 종은 긴자락송곳갯지렁이, 민숭대 나무갯지렁이, 작은갈매기고리갯지렁이, 삼각모자갯지렁 이였다. 한편, 정점군 I와 정점군 II간의 비유사도에 기여 하는 종은 민얼굴갯지렁이, 민숭대나무갯지렁이, 양손갯 지렁이류(Magelona sp.2) 등이였다(Fig. 8). 정점군 I 내의 I-1 과 I-2 또는 I-3간의 비유사도에 기여하는 종은 주요 우 점종으로, I-1에는 거의 출현하지 않았다.
2012년 8월과 2013년 2월 기간 동안 수심이 30 m 이하 인 연안 정점만을 가지고 집괴분석과 MDS 분석을 실시 한 결과, 발전소 배수구에 인근한 정점(정점 7, 9, 12, 14) 에서 여름철(군집 I; 정점 7, 9, 12, 14)과 겨울철(군집 II;
정점 3, 8, 9, 12, 14) 동안 군집 범위 차이가 있었으나, 나 머지 정점들(군집 III)은 두 기간 동안 차이가 없었다(Fig.
9). 여름철의 경우, 발전소를 중심으로 취수구에 위치한 정 점 7에서 죽변항 이전까지의 정점 14가 동일 군집(군집 I) 으로 묶여졌으나, 겨울철에는 취수구 정점 7을 제외한 정 점 3, 8, 9, 12, 14가 동일 군집(군집 II)으로 나타났다.
SIMPER 분석결과 군집 I은 민얼굴갯지렁이와 삼각모자 갯지렁이, 실타래갯지렁이류(Chaetozone sp.), Glycinde Table 4. Ecological character of same macrobenthic structure in February 2013
I
(st. 1, 3, 5, 7, 8, 9, 10, 12, 14, 16, 17,
18, 20, 22)
I-1 (st. 9, 12, 14)
I-2 (st. 1, 8, 10,
16, 17)
I-3 (st. 3, 5, 7,
18, 22)
II (st. 2, 4, 6, 11,
13, 15, 19, 21,23)
III (st. 20)
Depth 23.3 16.4 29.4 21.3 52.2 30.0
Gravel 3.6 15.5 0.0 0.0 2.5 0.0
SAND 90.0 83.1 89.3 94.9 51.0 99.8
SILT 3.3 0.6 5.9 2.5 28.8 0.1
CLAY 3.1 0.9 4.8 2.7 17.7 0.1
MEAN 2.5 1.2 2.9 2.7 4.8 0.6
SORTING 1.0 0.9 1.2 0.8 2.6 0.4
TN TOC
Temp sediment 10.5 11.1 10.2 10.3 9.3 10.2
Salinity in bottom 33.8 33.9 33.7 33.8 33.4 33.7
Temp in bottom 9.5 9.5 9.4 9.5 8.7 9.3
Oxygen in bottom 10.9 10.6 11.1 10.8 11.8 10.8
S 35.8 21.0 51.8 28.6 51.1 30.0
N 1946.2 1078.3 2107.0 2306.0 1580.6 980.0
H'(loge) 1.9 1.5 2.6 1.6 3.1 2.5
biomass 40.1 16.6 79.6 14.8 162.0 39.6
Spiophanes bombyx 1064.2 660.0 748.0 1623.0 11.3 0.0
Magelona sp.2 0.0 0.0 0.0 0.0 298.1 0.0
Lumbrineris longifolia 85.4 1.7 159.0 62.0 85.6 0.0
Scoloplos armiger 55.0 78.3 79.0 17.0 75.6 65.0
Maldane cristata 0.4 0.0 1.0 0.0 126.9 0.0
Chaetozone sp. 16.9 0.0 18.0 26.0 36.3 0.0
Adontorhina subquadrata 16.2 0.0 42.0 0.0 27.5 0.0
Microclymene propecaudata 13.8 0.0 34.0 2.0 80.6 0.0
Goniada maculata 19.2 1.7 36.0 13.0 78.1 0.0
Ampelisca naikaiensis 49.6 0.0 105.0 24.0 5.0 10.0
Euclymene oerstedi 8.1 0.0 9.0 12.0 46.9 0.0
Anonyx simplex 27.3 10.0 2.0 63.0 1.9 10.0
Fig. 8. MDS ordination plots and the spatial distribution of 8 characteristic species emerged by the SIMPER analysis
in February 2013
gurjanova, 납작뿔붙은눈옆새우가 군집의 유사도에 50%
이상 영향을 주는 주요 기여종이었으며, 군집 II에 기여한 종은 민얼굴갯지렁이와 삼각모자갯지렁이, 민가시긴뿔옆 새우, 유형동물인 Emplectonema mitsuii 등이었다(Fig.
10). 나머지 군집 III에 기여한 종은 민얼굴갯지렁이와 실 타래갯지렁이류(Chaetozone sp.), 납작뿔붙은옆새우, 유형 동물인 E. mitsuii 등을 포함하여 총 11종이 유사도 50%
이상 영향을 주었다. 한편, 군집 I과 군집 II간의 비유사 도에 기여하는 종은 다모류에 속하는 민얼굴갯지렁이, 실타래갯지렁이류(Chaetozone sp.), 큰부채발갯지렁이류 (Anaitides sp.), 송곳갯지렁이류(Lumbrineriopsis sp.) 등이 었다(Fig. 10). 또한, 그룹 I 또는 II와 그룹 III의 비유사도
에 기여하는 종은 각각 다모류 긴자락송곳갯지렁이, 실 타래갯지렁이류(Chaetozone sp.), 나이카안경옆새우 등 이었다.
4. 고 찰
대형저서동물의 종 수·개체수·생체량
발전소가 가동 전인 1987년부터 가동이 시작된 1988년 부터 2012년까지 발전소 주변해역의 연성기질에 서식하 는 대형저서동물에 대한 문헌 자료를 비교하였다(Table 5). 2012−2013년 조사 기간 동안 대형저서동물의 총 종수 는 319종이었으며, 평균 출현 개체수는 3,330 개체/m
2였 Fig. 9. Dendrogram and 2-dimensional plot of stations result from the cluster analysis and MDS ordination using the
macrobenthic faunal abundance data, respectively in the lower depth of 30 m, 2012 −2013 (A, Ausgust; F,
February)
고, 평균 생체량은 131.96 g/m
2이었다. 이와 같은 결과는 2006−2007년 한국해양연구원이 울진원자력발전소 6개호 기 가동 1년 후에 조사한 자료와 유사한 경향을 보였다 (Yu et al. 2011). 대형저서동물은 온배수의 영향을 많이 받는 지역일수록 종수와 출현 밀도가 감소하는 것으로 알 려져 있고(Bamber and Spencer 1984), 지속적인 온배수 공급으로 인해 주변의 저서동물의 종수와 개체수가 감소 해 생태계 교란을 유발시키는 것으로 알려져 있다 (Kailasam and Sivakami 2004). 또한 저서동물은 해수 유
동이 증가하면 스트레스로 인해 개체수가 감소한다고 알 려져 있다(Lardicci et al. 1999). 그러므로 본 연구 지역에 서 발전소 가동 후 10년 동안 대형저서동물의 종수와 개 체수의 큰 차이가 나타나지 않는 이유는 배출되어지는 온 배수의 양을 제외한 다른 인위적인 교란이 거의 나타나지 않기 때문인 것으로 여겨진다. 본 연구에서 발전소 배수구 를 중심으로 반경 5 km 이내의 정점을 분석한 대형저서 동물의 종수와 개체수를 비교했을 때, 2012−2013년의 대 형저서동물의 종수는 약간 감소하는 경향을 보였지만, 개 Fig. 10. MDS ordination plots and the spatial distribution of 8 characteristic species emerged by the SIMPER analysis
in the lower depth of 30 m
체수는 급격히 증가하였다(Table 5).
대형저서동물 종 조성 및 우점종
두 기간 동안에 주요 출현한 우점종의 조성은 큰 차이 를 보이지는 않았지만, 차지하는 비율은 차이가 컸다. 일 반적으로 대형저서동물의 종 조성은 퇴적물의 안정성에 따라 변동이 있으며, 퇴적물의 변동에 따라 종 조성과 개 체수의 변동이 나타나는 것으로 알려져 있고(Abe et al.
2005), 발전소 주변 온배수 배출 지역은 일반적인 해양 환 경을 가진 타 지역보다 기회종과 온수종의 개체수가 높게 나타나는 것으로 알려져 있다(Bamber and Spencer 1984).
그러므로 과거 조사 결과와 비교했을 때, 본 조사에서 대 형저서동물의 종 조성에 큰 차이를 보이는 것은 발전소 주변의 환경 변화에 의한 영향으로 여겨진다. 또한 2006 년 이전까지 수행된 울진원자력발전소 주변해역 저서동물 조사결과와 비교하면 월등히 많은 종 및 밀도가 출현하였 다(Table 5). 동일한 채집기와 채집면적을 통한 조사임에 도 이러한 차이를 보이는 것은 본 조사가 과거조사에 비 해 상대적으로 많은 정점 및 넓은 범위에 걸쳐 이루어졌 으며, 과거 조사에서는 우점종으로 출현하는 다모류, 단각 류, 거미불가사리류 등에 대한 종 수준까지의 동정이 이루 어지지 않았기 때문으로 여겨진다. 과거에 기록된 우점
종 중 일부 단각류 및 이매패류에 대한 명확한 종 정보 를 알 수는 없으나, 본 조사에 우점한 민얼굴갯지렁이(S.
bombyx), 양손갯지렁이류(Magelona sp.2), 긴자락송곳갯 지렁이(L. longifolia), 솜털바퀴실타래갯지렁이(C. setosa) 등은 발전소 가동후인 1995년 이후부터 우점종한 종으로 계절 별로 큰 차이가 없었다(Table 6). 본 조사에서 최 우 점한 민얼굴갯지렁이(S. bombyx)는 수심이 30 m 이하인 정점에서 주로 출현하였으며, 민얼굴갯지렁이의 개체수는 2000년 이후 감소하는 경향을 보여 2001년에는 9 개체/
m
2로 매우 낮아졌고, 2006−2007년에는 95 개체/m
2였지 만, 본 조사에서는 평균 1,819 개체/m
2로 급격히 증가하였 다(Fig. 11). 특히 1994년 봄철 기록된 이 종의 최대 밀도 2,856 개체/m
2(KEPCO 1995)와 본 조사결과(8월, 3,031 개체/m
2) 의 비교에서 큰 차이를 보이지 않았다. 본 조사에 서 극우점한 민얼굴갯지렁이의 경우, 발전소 배수구 아래 에 위치한 정점 9와 정점 12에서 극우점하였지만, 2006−
2007년 조사의 경우에는 이 지역에서는 거의 출현하지 않 았다. 그러나 발전소 배수구 주변 연안 해역에서의 민얼굴 갯지렁이의 개체수는 증가하였다(Yu et al. 2011). 본 조사 에서는 여름철에 개체수가 매우 높았지만, 겨울철에는 거 의 출현하지 않았다. 본 조사에서 여름철 이후, 발전소 배 수구 인근 연안 정점에서의 퇴적물이 조립화되고, 자갈의 Table 5. Annual variation in the number of species and density of macrobenthos around the study area (modified the
data from Yu et al. 2011) Item
Year No. sampling Season No. species Density
(ind/m
2) Reference 1987
(before operating) 16 season 53 318 KEPCO (1987)
1990* 13 season 103 1,362 KEPCO (1992)
1991 13 season 120 1,080 KEPCO (1992)
1992 5 season 121 997 KEPCO (1993)
1993 5 season 103 843 KEPCO (1994)
1994 5 season 88 1,455 KEPCO (1995)
1995 5 season − 832 KEPCO (1996)
1996 10 season 107 1,470 KEPRI (1997)
1997 10 Summer
Winter 68 748 KEPRI (1997)
KEPRI (1998)
1998** 10 season 143 739 KEPRI (1999)
1999*** 10 season 114 805 KEPRI (2000)
2000 10 season 169 430 KEPRI (2001)
2001 10 season 153 225 KEPRI (2002)
2002 10 season 190 317 KEPRI (2003)
2003 10 season 170 335 KEPRI (2004)
2006−2007**** 23 season 334 3.221 Yu et al. (2011)
2012−2013 23 Summer
Winter 319 3,330 In this study
*, 2 years after 1 unit operating; **, 3 units operating; ***, 4 units operating; ****, 1 year after 6 units operating
함량이 증가하였다(Fig. 2). 민얼굴갯지렁이는 자갈의 함 량이 매우 낮은 가는 모래 퇴적물에서 관을 만들어 서식 하며, 표층 퇴적물에서 먹이를 섭식하는 표층퇴적물식자 이다(Macdonald et al. 2010). 민얼굴갯지렁이는 발전소 배수구 주변 사질 퇴적물에서 주로 출현하며, 환경 변화에 민감하게 반응하는 기회주의 특성을 가지는 종으로 알려 지고 있다(Yu et al. 2013). 특히, 해수의 흐름이 증가하여 퇴적상이 조립하게 변화하면 개체수가 급격히 감소한다 (Marlin 2005). 그러므로 본 조사에서 민얼굴갯지렁이의
개체수는 조사 지역 내의 퇴적물의 퇴적상이 자갈이 거의 없는 세립한 사질 퇴적상에서 자갈이 포함된 조립한 사질 퇴적상으로 변화함에 따라 크게 감소한 것으로 여겨진다.
반면에 긴자락송곳갯지렁이는 2001년에는 같은 속 (Genus)에 속하는 긴자락송곳갯지렁이류(L. nipponica)가 평균 24 개체/m
2출현하였으나, 2006−2007년에는 개체수 가 3배 이상 유의하게 증가하였고, 2012−2013의 경우에 는 95 개체/m
2로 큰 차이를 보이지 않았다. 2006−2007년 자료와 군집분석을 실시한 결과 긴자락송곳갯지렁이는 발 Table 6. Temporal change in the dominant species and their percentage composition of the macrobenthic communities
in the study area (modified the data from Yu et al. 2011)
Year Spring Summer Fall Winter
1987 Spiophanes bombyx Amphipoda spp.
Nephtys caeca
33 23 11
Amphipoda spp.
Magelona japonica Maldane sarsi
43 21 11
Amphipoda spp.
Prionospio pinnata
49 12
Amphipoda spp.
Prionospio pinnata
46 17
1995 Spiophanes bombyx Nephtys ciliata
27 15
Spiophanes bombyx Nephtys ciliata
70 11
Spiophanes bombyx Chaetozone sp.
66 8
Spiophanes bombyx Nephtys ciliata
80 6 1996 Spiophanes bombyx
Gammeridae spp.
37 35
Spiophanes bombyx Gammeridae spp.
63 18
Spiophanes bombyx Gammeridae spp..
55 25
Spiophanes bombyx Chone teres
28 20
1997 Gammeridae spp.
Praxillella affinis Ophiuroidea spp.
Spiophanes bombyx
19 9 9 8
Prionospio sp.
Spiophanes bombyx Gammeridae spp.
Lumbrineis spp.
31 24 9 8 1999 Spiophanes bombyx
Gammeridae spp.
Pillucina pisidium
53 19 6
Spiophanes bombyx Pillucina pisidium Lumbrineris spp.
Gammeridae spp.
48 9 6 6
Spiophanes bombyx Lumbrineris spp.
Gammeridae spp.
Pillucina pisidium
51 8 6 5
Spiophanes bombyx Cirolanidae unid.
Lumbrineris longifolia 57 14 6
2000 Spiophanes bombyx Lumbrineris longifolia Ophiura sp.
45 7 4
Spiophanes bombyx Euzonus ezoensis Lumbrineris longifolia Ophiura sp.
20 10 7 7
Spiophanes bombyx Ophiura sp.
Bivalvia unid.
Lumbrineris longifolia 27 13 8 6
Spiophanes bombyx Bivalvia unid.
Glycera chirori Lumbrineris longifolia
34 15 6 5
2001 Euzonus sp.
Urothe sp.
Magelona japonica Goniada maculata Lumbrineris nipponica Spiophanes bombyx Praxillela affinis
10 9 6 5 4 4 3
Lumbrineris nipponica Praxillela affinis Urothe sp.
Melinna cristata Goniada maculata Ophiactis brancygenys Armandia lanceolata
11 8 8 5 4 4 3
Bivalvia juvenile Magelona japonica Urothe sp.
Amphelisca sp.
Eohaustorius sp.
Goniada maculata Lumbrineris nipponica
19 18 10 4 4 4 3 2002 Spiophanes bombyx
Lumbrineridae unid.
Ophiura sp.
Gammeridae unid
12 8 6 5
Euzonus ezoensis Lumbrineris longifolia Melinna elisabethae
15 5 5
Chaetozone setosa Spiophanes bombyx Euzonus ezoensis
32 6 6
Magelona japonica Spiophanes bombyx Gammeridae unid
20 11 6
2003 Ophiura sp.
Rynchospio glutaea Lumbrineris longifolia Chaetozone setosa Armandia lanceolata
28 9 5 6 6
Lumbrineris longifolia Spiophanes bombyx Chaetozone setosa
14 11 6
Chaetozone setosa Spiophanes bombyx Lumbrineris longifolia Magelona japonica
19 10 7 7
Ammotrypane aulogaster Chaetozone setosa Lumbrineris longifolia Magelona japonica Ophiura sp.
20 11 6 7 6 2005 −
2006
Magelona japonica Spiophanes bombyx Lumbrineris longifolia
18 13 10
Spiophanes bombyx Magelona japonica Lumbrineris longifolia
43 15 2.4
Magelona japonica Spiophanes bombyx
11 8
Magelona japonica Spiophanes bombyx Chaetozone setosa
21 6 6 2012 −
2013
Spiophanes bombyx Magelona sp2.
Scoloplos armiger Chaetozone sp.
62 2.5 2.2 2
Spiophanes bombyx Magelona sp2.
Lumbrineris longifolia Scoloplos armiger
34
65
4.5
3.8
Fig. 11. Annual variation in the density of Spiophanes bombyx and Lumbrineris longifolia for 20 years
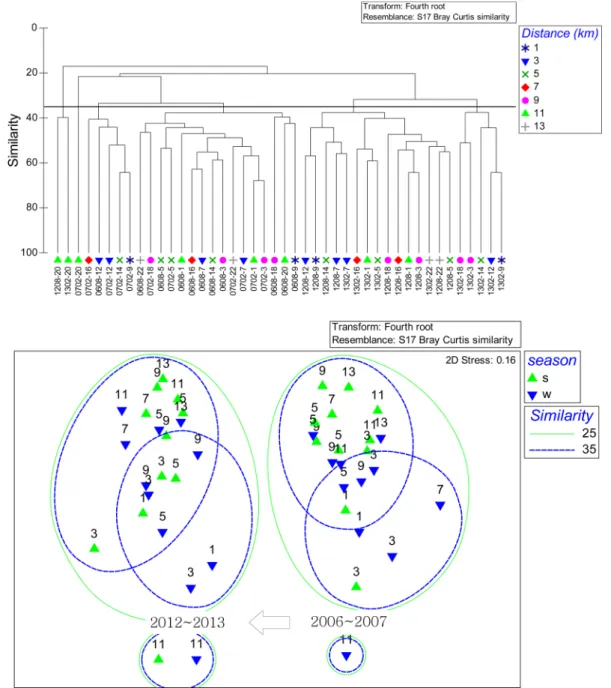
Fig. 12. Cluster analysis and MDS ordination of Macrobenthos between 2006 −2007 and 2012−2013 (The marks in
clustering means the distance from a power plant, km)
전소 취수구 북쪽 외측 지역에서 지속적으로 개체수가 증 가하고 있다(Figs. 12, 13). 또한 양손갯지렁이류(Magelona sp.2) 는 2006−2007년 조사에서 수심이 깊은 외해역에서 개체수가 매우 높았지만(Yu et al. 2011), 본 조사에서는 높지 않았다. 또한, 외해역에서 우점했던 살시거미불가사
리류(Ophiura sp.)는 본 연구에서 개체수가 높지 않았다.
과거 온배수 영향 사질 퇴적물에 주로 출현하였던 온수종
인 단각류 Urothoe convexa(블루손모래무지옆새우사촌),
다모류 Rhychospio sp., 이매패류 Felaniella sowerbyi(꼬
마돌살이조개) 등은 본 연구 기간 동안 발전소 주변 사질
Fig. 13. MDS ordination of contributed species by SIMPER analysis between 2006 −2007 and 2012−2013
지역에 거의 출현하지 않았다. 그러므로 본 조사에서 우점 한 종들의 출현 밀도는 과거 2006−2007년 조사와 많은 차이를 보이고 있어, 이는 온배수 주변 지역의 환경변화에 의한 것으로 여겨지며, 이에 대한 원인과 경향을 파악하기 위한 지속적인 연구가 필요할 것으로 여겨진다.
대형저서동물의 군집 구조
본 연구와 동일한 정점에서 채집이 이루어졌던 2006−
2007년의 여름과 겨울에 출현한 대형저서동물과 함께 군 집구조 분석을 실시한 결과, 두 시기 동안의 군집구조는 유사도 20%로 서로 다른 시간적인 군집구조를 나타냈다.
그러나 각 시기 동안 정점 간 군집구조는 과거 조사와 같 은 정점군으로 나타나 서로 유사한 형태를 보였다(Fig.
12). 이와 같은 시간적인 군집구조의 차이는 본 조사 지역
에서 일부 우점하는 종들이 2006−2007년에 비해 개체수
가 급격히 증가하였기 때문으로 여겨지며, 정점 간 군집구
Fig. 14. The long-term variation in the spatial distribution of macrobenthic fauna in the study area for 10 years
조의 유사성은 2006−2007년 과거 조사와 본 조사간 정점 간 종 조성의 차이는 거의 없었기 때문으로 여겨진다(Fig.
13). 특히, 우점종 가운데 민얼굴갯지렁이는 2006−2007년 보다 2012−2013년에 급격히 증가하였는데, 이는 발전소 주변 기회종의 개체수가 일반적인 환경의 다른 지역에 비 해 상대적으로 높다는 Bamber and Spencer (1984)의 연 구 결과와도 잘 일치한다. 과거 10년 동안 울진원자력발 전소 주변 조하대에 서식하는 대형저서동물 군집구조를 분석한 결과, 대형저서동물의 군집구조는 크게 수심이 30 m 보다 낮은 지역 정점군과 깊은 지역 정점군, 두 개의 군집으로 구분되었다(Fig. 14). 수심이 낮은 지역 내에서 의 군집구조는 조사시기별 그리고 계절별 차이를 보이고 있다. 특히 발전소 4개호기 가동 2년 후의 대형저서동물 군집은 겨울철에는 배수구 주변해역과 차이를 보이지 않 았으나, 6개호기 가동 이후부터는 겨울철을 포함한 모든 계절에 차이를 보였다. 이는 발전소 6개호기 가동에 따른 온배수의 배출량과 연관이 있는 것으로 여겨진다. 발전소 온배수의 지속적인 공급은 발전소 주변의 생태계 교란을 유발시키며(Kailasam and Sivakami 2004), 기회종과 온수 종의 개체수가 일반적인 해양 환경의 타 지역에 비해 높 은 것으로 알려져 있다(Bamber and Spencer 1984). 또한 발전소 배수구 주변에 서식하는 해양 생물들은 발전소 배 수구에서 배출되는 온배수의 영향으로 배수구 주변 수온 에 적응되어 있는 것으로 알려져 있다(Pilati 1976;
Mclennan and Miller 1990). 그리고 대형저서동물의 군집 구조는 계절적 수온 변화와 각 발전소 가동 정지에 따른 온배수 배출량의 차이로 인한 저층수온 변화에 복합적 영 향을 받는다고 했다(Yu et al. 2013). 그러므로 본 연구에 서 발전소 가동 후 장기간의 대형저서동물 군집구조는 자 연적인 환경변화에 의한 것보다 온배수 배출량과 연관이 높은 것으로 여겨지며, 향후 온배수 배출량의 변동이 있을 경우 발전소 주변해역의 대형저서동물 군집 변화가 나타 날 가능성이 높아, 지속적인 모니터링이 필요할 것으로 여 겨진다.
사 사
본 논문을 세심하게 검토해주신 두 분의 심사위원께 깊 은 감사를 드립니다. 본 연구는 한국해양과학기술원의 연 구과제(PI55574, PE99517)의 지원으로 수행되고 분석되 었습니다.
참고문헌