185
청국장 발효 균주인 Bacillus subtilis 028-1의 항생물질 생산과 특성
안경준 서원대학교 과학교육과
Bacillus subtilis 028-1 균주는 청국장 발효에 사용하는 균주로 Staphylococcus sp. LS2 뿐만 아니라 여러 yeast 균주 의 생장을 억제하는 항생물질을 생산하며 , soybean meal 2% 와 maltose 와 같은 이당류를 1% 첨가하여 15~18 시간 진탕 배양하였을 때 최대의 항생물질 생산을 보였으며 배지의 pH 는 6.5 이하였다 . 항생물질의 활성은 약염기성 조건에서 극대화되었으며 , 100
oC 에서 20 분간 가열하여도 활성은 크게 감소하지 않았고 , 실온 보관 시 한 달 이상
효과가 지속되며 chymotrypsin 과 papain 같은 단백질 분해효소 처리에 의해 서서히 활성이 줄어들었다 . 투석에
의해 항생물질의 분자량을 측정한 결과 1,000 에서 500 dalton 사이인 것으로 나타났으며 항미생물 효과는 있으나
fibrin 분해 능력이 없으므로 surfactin 이 아닌 iturin 계열의 peptide 성 항생물질로 보인다 .
Key words □ antimicrobial activity, Bacillus subtilis , chungkookjang, peptide antibiotics, soybean meal medium
내생포자를 생산하는 그람양성 토양세균에서 대표적인
Bacillus 속 세균들은 각종 발효식품 생산을 비롯한 농업 및 공
업적으로 유용한 세균으로서 구조상으로 놀라운 다양성을 보이 는 여러 가지 항생물질을 생산한다 . Bacillus 속 세균을 배양하면 한 가지 이상의 필수영양소 성분이 고갈되고 , 이는 균 생장을 정 체기로 들어가게 한다 . 이 시기에 포자 형성 , competence 화 , 세
포 외부로의 분해효소 생산 , 그리고 항생물질 생산에 관련된 유 전자 무리들의 발현이 유도된다 . 이러한 다양한 반응은 영양
stress 에 대한 생존 전략이며 항생물질의 생산은 생산균주로 하여
금 제한된 영양분이 존재하는 환경에서의 경쟁력을 강화하기 위 함으로 보여진다 . 여러 종류의 항생물질 생합성에는 350 kb 에 달 하는 유전자들이 동원되며 이는 전체 genome 의 4~5% 에 해당한
다 (30). 항 미생물 활성을 보이는 Bacillus 속이 생산하는 항생물
질들은 주로 peptide 류로서 ribosome 에 의해 합성되고 차후에
thioether 결합을 포함하는 특이한 lanthionine 구조를 갖게 되는 lantibiotics 종류 (23) 와 ribosome 에 의하지 아니하는 lipopeptide 를 비롯한 각종 peptide 들 , 그리고 peptide 가 아닌 polyketide, aminosugar, phospholipid 들이 있다 (42). 이들은 1 차적으로 항 미 생물 활성을 보임과 동시에 , ribosome 에 의하지 않는 surfactin
같은 lipopeptide 는 biofilm 형성이나 swarming 발달에 기여하며 , lantibiotics 는 quorum-sensing 과정에서 pheromone 역할을 담당 하기도 한다 (20).
Bacillus subtilis 가 생산하는 항생물질의 주종인 peptide 들은 구 조적으로 매우 단단하며 소수성이고 환상구조로 되어있으며 ,
D형 아미노산을 포함하고 있어서 peptidase 나 protease 에 저항성을 나타낸다 .
또한 ribosome 에 의하지 않는 lipopeptide 항생물질들은
peptide 에 덧붙여서 β -hydroxyl 또는 β -amino fatty acid 가 모여진
구조를 가지며 (17), non-ribosomal peptide synthetase (NRPS) 라고 불리는 큰 효소에 의해 독특하게 합성되는데 , 이들이 세포 내의
특정 목표에 정확하게 작용하는 것이 알려지면서 새로운 약품의
제조에 기대를 모으고 있다 (41).
이러한 lipopeptide 는 B. subtilis 가 생산하는 가장 보편적인 항 생물질로서 amphiphilic 한 특성은 계면에서의 물리적 , 화학적 성
질을 변화시킬 수 있어서 소수성 기질의 이용 능력을 향상시키 며 , 미생물이 표면에 부착하거나 탈락하는데 영향을 주기도 한다 .
이런 역할은 B. subtilis 의 자연계에서의 생존에 강력한 영향을
줄 것이며 , 가장 잘 연구된 것은 anionic lipoheptapeptide 인
surfactin 이다 (37). Surfactin 은 β -hydroxy fatty acid 의 carboxyl group 과 heptapeptide 의 hydroxyl group 이 결합한 구조로 되어 있으 며 , 이의 생산은 B. subtilis 외에도 B. pumilus , B. amyloliquifaciens 들 에 널리 분포하고 있으며 많은 종류의 idiotype 이 존재한다 .
한편 surfactin 에 부가하여 fengycin, iturin 같은 항 미생물 및
항균작용이 있는 lipopeptide 는 B. subtilis 로 하여금 경쟁자를 제 거하는 이점을 제공한다 (30). Iturin family 는 밀접하게 연관된 환
상 lipoheptapeptide 로서 B. subtilis 를 포함한 Bacillus 속 여러 균 주들에서의 합성이 보고된 iturin (5), mycosubtilin (45), bacillomycin (6), bacillopeptin (16) 등을 포함하는데 , 이들은 강 력한 항 곰팡이 효과 (1, 3, 10, 12, 18, 21, 24, 35, 40, 44) 와 용
혈성을 보이지만 항 세균 활성 (36) 은 낮은 편이다 . Iturin 종류는
peptide 의 1~3 번 자리가
L-Asx,
D-Tyr,
D-Asx 로 구성되고 , 6 번 자리가
D-amino acid 로 되어 있으며 β -amino fatty acid 를 가지
고 있어서 (5) 세포막의 인지질 이중층에 수월하게 끼어들게 되고 이어서 환상의 peptide 는 일종의 ion conducting pore 를 형성하여 (27, 28, 29) 세포막의 투과성을 변화시켜 proton gradient 를 파괴
*To whom correspondence should be addressed.
Tel: 82-43-299-8403, Fax: 82-43-299-8400
E-mail: [email protected]
함으로써 세균의 편모 운동성을 저해하거나 특히 곰팡이와 같은 진핵생물의 세포막 손상을 유발하는데 이때 cholesterol 의 존재는 이 과정을 가속화시키는 것으로 알려져 있다 (26).
한편 , B. subtilis 말고도 Bacillus 속에 속하는 여러 균주들로부 터 다양한 종류의 lipopeptide 의 생산이 보고되어 있다 (8, 11, 14, 15, 19, 31, 38).
식품을 장기간 안전하게 보존하기 위하여 방부제를 비롯한 여 러 가지 화학물질을 첨가하고 있으나 장기간의 섭취로 인하여 체내에서 발암물질로 작용하기도 하고 크고 작은 여러 가지 질 환의 원인이 되고 있는 실정이다 . 현재 미생물에 의해서 생산된 물질을 천연방부제로 식품에 사용하고 있는 것은 유산균인
Lactococcus lactis 가 생산하는 nisin 등이 있으나 적용 대상이 제
한되는 문제점이 있다 (18).
대두를 이용한 발효식품인 청국장은 발효 숙성 과정 중에 B.
subtilis 나 B. natto 등이 생산하는 효소 작용으로 인해 콩 단백질 이 분해되어 그 특유의 맛과 냄새를 내는 동시에 끈적끈적한 점 질물이 다량 생성된다 . 청국장은 다른 장류와 함께 단백질 섭취 량이 비교적 적었던 한국인에게는 단백질의 중요한 공급원이었 으며 영양면에서도 된장이나 고추장보다 단백질과 지방함량이 높은 영양식품이며 생리활성물질인 식이섬유와 isoflavone 을 비롯
한 다양한 건강증진 성분을 포함하고 있다 .
본 연구는 우리가 오랜 동안 안전하게 섭취해 오고 있는 청국 장 제조 균주를 배양하여 천연방부제 역할을 할 수 있는 성분을 효율적으로 생산하는 배양 방법을 확립하고 그 성분을 분리하여 특성을 조사함으로써 화학적 식품 보존제 및 방부제 , 항 곰팡이 제 제를 대체하는 물질 (22) 을 개발하는 연구의 일환으로 시도하였다 .
재료 및 방법
균주, 배지 및 배양조건본 실험에서는 호기적 조건에서 빠른 생장을 보이고 내생포자 를 생산하며 점질이 양호한 생청국장을 만드는 균주들을 충북 음성군 생극면에서 수확한 볏짚 등에서 분리하였고 , 이 중에서
청국장 음료를 생산하기 위하여 특허 출원한 균주인 Bacillus
subtilis 028-1 (KCTC 11099BP) 균을 항생물질 생산균주로 사용
하였다 . 균주의 동정을 위해서 API 50 CHB kit (France,
Biomerieux) 를 사용하여 탄수화물 이용 범위를 알아보았으며 생
산된 항생물질의 생장 억제 효과를 보기 위해 그람양성세균인
Staphylococcus sp. LS2, Bacillus subtilis ATCC 6633, Bacillus cereus 1-1 과 그람음성세균인 Escherichia coli K12, Klebsiella pneumonieae M5a1, Pseudomonas putida mt2, 그리고 효모균인
Candida albicans KCTC 7965, Saccharomyces cerevisiae ATCC
26603 을 시험균주로 사용하였다 .
사용배지로는 볏짚으로부터 균주를 분리하거나 계대배양하기 위하여 LB 배지 (tryptone 1%, yeast extract 0.5%, NaCl 0.5%, agar 1.5%) 와 soybean meal 배지 (soybean meal 2%, agar 1.5%)
를 사용하였고 균주들이 생산하는 항생물질의 효과를 cylinder method 로 알아보기 위하여 PCA 평판배지 (tryptone 0.5%, yeast
extract 0.25%, agar 1.5%) 를 제조하여 하층배지로 사용하였고 ,
0.5% 의 한천을 첨가한 PCA 배지를 상층배지로 사용하였다 .
효모에 대한 항생물질의 효과를 보기 위하여 YM 배지 (yeast extract 0.3%, malt extract 0.3%, peptone 0.5%, glucose 1%, agar 1.5%) 를 PCA 배지의 경우처럼 사용하였다 .
청국장균의 항생물질 생산을 위해 액체 soybean meal 배지
(soybean meal 1~2%) 를 사용하였으며 , 당을 첨가하여 진탕배양
하면서 최적의 배양 조건을 찾았다 .
항생물질에 대한 감수성 조사
선발된 청국장 제조 균들을 LB, PCA, YM, soybean meal 평
판배지 중앙에 한 줄로 접종하여 1 일 배양하여 항생물질을 생산 하게 한 다음 대상균주들의 감수성 여부를 알아보기 위해 좌우 로 접종하여 30
oC 에서 키우면서 생장 억제 정도를 알아보았다 .
항생물질의 생장 억제효과
B. subtilis 028-1 균주가 생산하는 항생물질의 대상균에 대한 생장 억제 효과를 알아보기 위하여 그람양성균인 Staphylococcus sp. LS2, 효모균 Candida albicans KCTC 7965, Saccharomyces cerevisiae ATCC 26603 을 대상으로 하였다 .
항생물질의 활성은 대상균주의 배양액을 상층배지에 섞어 하
층배지 위에 얇게 깐 다음 , 8 mm 의 cylinder 를 설치하여 항생물
질을 50 µ l 씩 가하고 1 일간 배양한 후 나타나는 투명환의 크기를 세 번의 반복 실험을 통한 평균값으로 구하였다 .
항생물질 생산 및 효율성 조사
500 ml 진탕 플라스크에 1% soybean meal 배지 100 ml 를 만 들고 B. subtilis 028-1 균을 접종하여 32
oC 에서 280 rpm 으로 진탕 배양하였다 .
실험하고자 하는 조건 별로 배양을 한 후 배양액은 10,000 × g
로 10 분간 원심 분리하여 상층액만을 모았으며 이 과정을 두 번
반복하였다 .
배지 종류에 따른 영향을 알아보기 위해서 LB, PCA, YM,
soybean meal 액체배지를 만들어 028-1 균주를 접종하고 진탕
배양하여 생산된 항생물질의 활성을 cylinder method 에 의한 투
명환의 크기로 비교하였다 .
Soybean meal 농도에 의한 영향을 알아보기 위해 농도를 1%,
2%, 3% 로 달리하여 028-1 균주를 배양 후 항생물질을 획득하여
비교하였다 .
당류 첨가에 따른 항생물질 생산의 변화를 알아보기 위하여 단당류로 glucose, 이당류로 lactose, maltose, 그리고 sucrose 를 첨가하여 비교하였다 .
Maltose 농도에 의한 영향을 알아보기 위해 2.0% 의 soybean meal 배지에 maltose 농도를 0%, 0.5%, 1.0%, 1.5%, 2.0% 로 달
리하여 028-1 균주를 접종하여 280 rpm, 32
oC 에서 진탕 배양하 면서 12 시간부터 3 시간 단위로 상등액을 획득하여 생장 억제 효
과를 조사하였으며 , maltose 를 첨가하지 않은 경우와 0.5% 첨가
한 배지에서 028-1 균주를 배양하며 3 시간 간격으로 pH 변화를
측정하였다 .
항생물질의 농축 및 특성 조사
B. subtilis 028 - 1 균주가 항생물질을 활발히 생산하는 최적배
지인 soybean meal 배양액을 원심분리하여 균을 제거하고 농축
하여 항생물질로 사용하였다 . 농축은 동결건조와 n-butanol
extraction 을 거친 다음 하층의 수용액 부분을 회전농축하는 두
가지 방법으로 진행하였으며 농축 정도는 약 10 배로 하였다 .
단백질로 추측되는 항생물질 성분을 침전시키기 위하여
ammonium sulfate 와 polyethylene glycol 2000, 4000, 6000, 8000, 10000 (Sigma, USA), polyvinylpyrrolidine (PVP-40T, Sigma) CaCl
2, HCl 을 첨가하거나 , 용매추출법으로 chloroform, ethyl alcohol, methyl alcohol 등을 첨가하면서 침전을 유발하였다 .
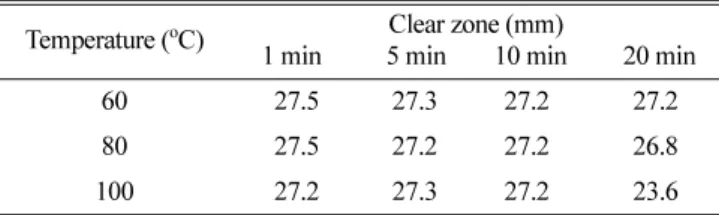
10 배 농축한 시료를 60
oC, 80
oC, 100
oC 의 각 조건에서 1 분 , 5
분 , 10 분 , 20 분간 열처리를 한 후 대상균에 처리하여 나타나는 투명환의 직경을 측정하였으며 , 시료를 20
oC, 4
oC, -20
oC 조건에 보관하면서 보관기간에 따른 활성 변화를 알아보았다 .
대상균으로 LS2 균을 이용하여 PCA 액체배지의 초기 pH 를
pH 5~8 로 조절한 다음 , 전 배양한 대상균을 1% 농도로 첨가하
여 배양하면서 600 nm 에서의 흡광도가 0.2 정도 되었을 때 농축
한 028-1 균 배양액을 1% 첨가하였고 OD 를 측정하여 대상균의
생장에 미치는 억제 효과를 비교하였으며 , 대조군으로 028-1 균
배양액을 첨가하지 않은 대상균의 생장도 같이 조사하였다 .
028-1 균 배양액에 포함되어 있는 항생물질의 성질을 알기 위
하여 단백질 분해효소를 처리하여 항생물질의 효과가 감소하는 지 알아 보았다 .
사용한 단백질 분해효소는 chymotrypsin (C-4129, Sigma), trypsin (T-4799, Sigma), papain (P-3125, Sigma), proteinase K (P-8044, Sigma) 를 각각 10 mg/ml 의 농도로 시료와 혼합한 후
37
oC 에 두면서 시간 별로 채취하여 cylinder method 에 의해 나타 나는 투명환의 크기를 비교하였다 .
항생물질 성분의 분자량을 추정하기 위하여 투석을 실시하였 다 . MWCO (molecular weight cut off) 값이 12,000 인 dialysis tube (35 mm, Spectra/Por, Spectrum Laboratories Inc., Canada) 로 분자량 12,000 이하를 얻었으며 , 이어서 MWCO 1,200 정도인 benzoylated dialysis tubing (32 mm, D-7884, Sigma) 을 이용하여
12,000 에서 1,200 사이와 1,200 미만을 각각 얻었고 , 분자량
1,000 미만과 500 미만은 각각 1,000 과 500 MWCO cellulose ester dialysis tube (31 mm, Spectra/Por, Spectrum Laboratories Inc.) 를 이용하여 얻었으며 , 대상균 LS2 에 처리하여 생육 억제 효과가 있는 부분을 찾았다 .
B. subtilis 028 - 1 균주가 생산한 항생물질이 biosurfactant 의 일
종인 surfactin 인지 알아보기 위하여 fibrin 용해 여부를 조사하였 다 . 0.6% 의 fibrinogen (F-3879, Sigma) 에 1.0 unit/ml 의 thrombin (T-6884, Sigma) 을 섞고 , 미리 PBS buffer (0.15 M NaCl, 0.01 M phosphate buffer, pH 7.8) 에 녹인 1% agarose 를 동량 가하여 잘
섞으면서 petri dish 에 부어 굳힌다 . Fibrin 이 생성된 것을 확인하
고 8 mm paper disk 를 놓은 다음 투석을 통해 얻은 분자량
1,000 미만의 시료를 가하여 30
oC 에 보관하면서 fibrin 이 용해되 는지 관찰하였다 .
결과 및 고찰
균주의 동정 결과B. subtilis 028-1 균주는 그람양성이며 내생포자를 생산하고
OF test 에서 oxidation 에 의하여 생장하였으며 청국장 제조 과정
에 첨가하였을 때 점질물질을 다량 만들어서 청국장 음료 생산
을 위하여 미리 특허 출원한 균주 (KCTC 11099BP) 이다 . 028-1
균주를 LB 평판에서 하룻밤 배양하고 API 50 CHB kit 를 이용
하여 2 일간에 걸쳐 028-1 균주의 다양한 탄수화물에 대한 이용
여부를 판독한 결과는 Table 1 과 같다 . 전형적인 B. subtilis 균과 비교해 본 결과 salicin, cellobiose, starch, glycogen 에 대한 이용 능에서 차이가 나는 것으로 나타났으며 , 이 자료를 Biomerieux
회사 (www.biomerieux.com) 에서 제공하는 Apiweb 을 통하여 분석
Table 1. Biochemical characteristics (carbohydrates) of B. subtilis 028-1
Carbohydrate
source Result Carbohydrate
source Result
Glycerol + Salicin -
Erythritol - Cellobiose -
D-Arabinose - Maltose +
L-Arabinose + Lactose -
Ribose + Melibiose -
D-Xylose + Sucrose +
L-Xylose - Trehalose +
Adonitol - Inulin +
β -Methyl-D-xyloside - Melezitose -
Galactose - Raffinose -
Glucose + Starch -
Fructose + Glycogen -
Mannose + Xylitol -
Sorbose - Gentiobiose +
Rhamnose - D-Turanose +
Dulcitol - D-Lyxose -
Inositol + D-Tagatose -
Mannitol + D-Fucose -
Sorbitol + L-Fucose -
α -Methyl-D-mannoside - D-Arabitol -
α -Methyl-D-glucoside + L-Arabitol -
N-Acetyl glucosamine + Gluconate -
Amygdalin - 2-Keto-glu-
conate -
Arbutin + 5-Keto-glu-
conate -
Esculin +
한 결과 B. subtilis 의 가능성은 99.4% 로 나타났다 . 한편 , B.
subtilis 028-1 균주는 최소배지에서 키웠을 때 biotin 을 미량 요 구하였다 .
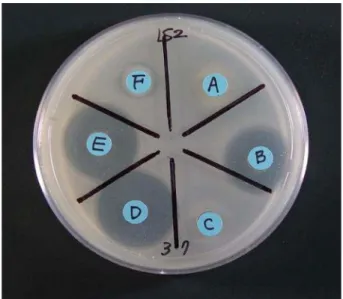
항생물질에 대한 미생물의 감수성
LB, PCA, soybean meal 평판배지에서 B. subtilis 028-1 균주 가 생산한 항생물질에 대한 세균별 감수성 조사를 한 결과 , LB
와 PCA 평판배지에서는 항생물질 효과가 아주 미약하거나 나타
나지 않았으며 soybean meal 평판배지에서는 E. coli K12, K.
pneumonieae M5a1 와 Staphylococcus sp . LS2, B. cereus 1-1 에 대해서 생장 억제 효과를 강하게 보였다 .
대상 균주에 따른 생장 억제 효과를 알아보기 위해 B. subtilis
028-1 균주가 항생물질을 생산하는 1% soybean meal 액체배지
에서 1 일간 배양하여 얻은 시료를 동결건조를 통하여 10 배 농축
하고 , cylinder method 를 통해 생장 억제를 보이는 투명대를 측
정하여 각 균에 대한 효과를 비교하였다 . 그람양성세균인
Staphylococcus sp. LS2 와 효모에 속하는 Candida albicans KCTC 7965, Saccharomyces cerevisiae ATCC 26603 에 대한 생장 억제 효과가 우수하였으며 결과는 Table 2 와 같다 .
항생물질 생산의 효율성 조사
Soybean meal 배지에서 생장 억제 효과가 크게 나타나는 이유
는 다양한 성분을 포함한 soybean meal 배지가 항생물질 생산에
유리하기 때문인 것으로 보인다 . Soybean meal 의 농도를 달리하
여 1 일간 배양하면서 그에 따른 항생물질의 생산 정도를
cylinder method 에 의한 투명환의 크기로 비교하였는데 1%, 2%,
3% 중에서 2% 를 첨가한 경우 투명환의 크기가 가장 크게 나타
났다 (Table 3). Soybean meal 의 농도가 3% 이상이 되면 오히려 투명환의 크기는 감소하였으며 , 배지 제조 및 배양액 처리 과정 에서도 불편하였다 . 그러나 Mizumoto 등 (33) 은 soybean meal 의 농도를 8% 로 하여 8 일간 배양하였을 때 기존에 좋은 성적을 보 였던 Kondoh 등 (24) 의 3S media (3% polypepton S, 1%
glucose, 0.1% KH
2PO
4, 0.05% MgSO
47H
2O, pH 6.8) 에 비해서
적어도 2 배 이상의 lipopeptide 일종인 iturin A 생산을 보고한 바 있다 . 오래 전부터 B. subtilis 를 이용하여 peptide 성 항생물질
을 생산할 때 사용되었던 배지로는 Landy medium (25) 이나 이 를 변형시킨 배지들 (2, 5) 이 있으며 최근에 들어서 soybean meal
을 사용한 항생물질 생산이 여러 번 시도되었다 (32, 33, 34).
Soybean meal 배지에 단당류로 glucose, 이당류로 lactose, maltose, sucrose 를 각각 1% 첨가하여 이에 따른 항생물질 생산
을 비교한 결과 B. subtilis 028-1 균주는 이당류를 첨가한 경우
항생물질의 생산이 증가하였으며 , 특히 maltose 를 첨가한 경우 효과가 좋았다 (Table 4). 이는 Mizumoto 등 (33) 이 B. subtilis RB14-CS 균주를 이용한 iturin A 의 생산을 위하여 soybean
meal 에 maltose 를 첨가한 조합과 같았으나 배지 농도와 배양 시
간에 있어서는 본 실험의 경우가 더 유리하였다 .
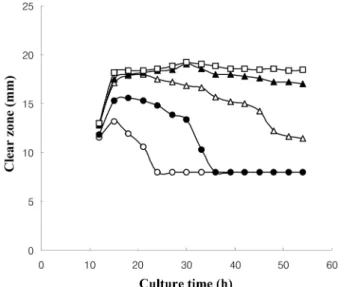
이당류 중 maltose 의 농도를 0%, 0.5%, 1.0%, 1.5%, 2.0% 로 달리하여 배양 시간에 따른 항생물질의 생산 정도를 비교한 결
과 , 1.0% 미만으로 첨가한 경우 항생물질의 생산이 배양 개시
15 시간에 최대치를 보이고 , 배양시간이 지속됨에 따라 maltose
농도가 낮을수록 빨리 감소하며 , maltose 가 1.5% 이상 첨가되면
배양시간이 지속되어도 활성은 크게 줄지 않았으며 최대 생산 시기는 15 시간에서 18 시간 사이였다 (Fig. 1). Rahman 등 (39) 에 의하면 B. subtilis 가 생산하는 lipopeptide 중에서 surfactin 은 대
Table 2. Antibiotic activity of 028-1 culture broth of 1% soybean meal against bacteria and yeast test strains
Test strains Clear zone (mm)
Staphylococcus sp. LS2 13.2
Candida albicans KCTC 7965 13.5
Saccharomyces cerevisiae ATCC 26603 22.2
Table 3. Antibiotic activity of 028-1 culture broth against Stapylococcus sp. LS2 according to the soybean meal concentration
Soybean meal (%) Clear zone (mm)
1 13.8
2 17.8
3 16.2
Table 4. Antibiotic activity of 028-1 culture broth against Stapylococcus sp. LS2 according to the addition of 1% saccharides
Saccharides Clear zone (mm)
Control 13.4
Glucose 16.5
Maltose 25.4
Sucrose 20.6
Lactose 22.0
Fig. 1. Antibiotic activity of 028-1 culture broth against Staphylococcus sp. LS2 according to the addition of maltose 0%( ○ ), 0.5% ( ● ), 1.0%
( △ ), 1.5% ( ▲ ), and 2.0% ( □ ).
수기 말에 최대 생산을 보이지만 iturin 은 정지기에 도달한 이후 에 생산되는 secondary metabolite 라고 하였는데 본 실험에서 생 산된 항생물질은 iturin 계열의 가능성이 크다 . Maltose 의 농도가 높아지면 항생물질 생산의 지속기간이 길어지긴 하였으나 활성 이 크게 증가하지는 않았다 .
Soybean meal 액체배지에 maltose 첨가 여부에 따라 B.
subtilis 028-1 균주를 배양하며 배양과정에서의 pH 변화를 3 시간 단위로 측정한 결과 균이 생장함에 따라 배지의 pH 는 상승하였 으나 0.5% 의 maltose 가 첨가된 배지에서는 pH 가 배양 20 시간
전후로 낮아지다가 24 시간 경과한 후부터 상승하기 시작하여 30
시간이 지나면 빠르게 올라감을 관찰할 수 있었다 . 이를 항생물
질 생산 결과와 비교해 보면 항생물질의 생산이 줄어드는 시간
과 pH 가 6.5 부근에 도달하는 시간이 배양 후 30 시간에 비슷하
게 나타나는 것으로 보아 pH 와 항생물질 생산에 연관관계가 있
으며 pH 6.5 이하의 조건에서 항생물질을 생산한다는 것을 보여
주는 결과이다 (Fig. 2). 그러나 maltose 를 첨가하지 않은 경우에 는 pH 가 지속적으로 상승하였으며 항생물질의 생산은 역시 Fig.
1 에서처럼 pH 6.5 를 넘어서는 시기인 15 시간 이후에 급격히 감
소하였다 .
위의 결과를 종합해보면 B. subtilis 028-1 균주는 배양시작 후
15~18 시간에 최대치의 항생물질을 생산하였고 maltose 가 첨가되
면 항생물질 생산이 지속되며 , soybean meal 의 농도는 2% 로 하
고 maltose 와 같은 이당류를 1% 첨가하여 배양하면서 배지의
pH 는 6.5 이하로 유지하는 것이 유리하다는 것을 알 수 있었다 .
항생물질의 농축 및 특성