Optimization of Medium Composition for Lipopeptide Production from Bacillus subtilis N7 using Response Surface Methodology
Luo, Yi, Guoyi Zhang, Zhen Zhu, Xiaohui Wang, Wei Ran*, and Qirong Shen
Jiangsu Key Lab for Organic Solid Waste Utilization, Nanjing Agricultural University, Nanjing 210095, China
Received : July 23, 2012 / Revised : October 31, 2012 / Accepted : November 1, 2012
Introduction
Bacillus subtilis lipopeptides (iturin, surfactin and fen- gycin) synthesized non-ribosomally via muti-enzymes are considered some of the most promising biosurfactants [6, 17]. Iturin A is a cyclic lipopeptide containing heptapep- tide (L-Asn-D-Tyr-D-Asn-L-Gln-L-Pro-D-Asn-L-Ser) cycled with a β-amino fatty acid. The surfactin family encom- passes structural variants, but all the members of this family are heptapeptides interlinked with a β-hydroxy fatty acid to form a cyclic lactone ring structure. Fengycin are lipode- capeptides with an internal lactone ring in the peptide moi- ety and with a β-hydroxy fatty acid chain that can be saturated or unsaturated [12]. Because of their amphiphilic
nature, lipopeptides readily associate with and tightly anchor into lipid layers; in addition they exhibit antagonistic activities against several plant pathogens by interfering with biological membrane integrity in a dose-dependent manner [7]. Along with their surface activity and wide spectrum of antibiotic activity, biologically-derived lipopeptides have advantages over synthetic chemical fungicides, specifically low toxicity, low allergic effect on humans and animals, and high biodegradability [14].
To meet the need for lipopeptides, most previous work emphasized the isolation and screening of hyper-producing microorganisms [2] that were cultured via submerged fer- mentation (SMF) [16] or solid-state fermentation (SSF) [11].
Fermentation processes mostly used organic nitrogen, car- bon, and potassium salt. However, few studies have been devoted to investigating trace elements and precursors such as Fe2+, Mn2+ and fatty acids, which are related to synthetase activity in Bacillus [8]. In industrial-scale produc- The nutritional requirements for the maximum production of lipopeptides by Bacillus subtilis N7 (B. subtilis N7) were investi- gated and optimized using response surface methodology (RSM) under shake flask fermentation. A one-factor-at-a-time experimental setup was used to screen carbon and nitrogen sources. A Plackett–Burman design (PBD) was employed to screen the most critical variables for lipopeptides production amongst ten nutritional elements. The central composite experi- mental design (CCD) was finally adopted to elucidate the composition of the fermentation medium. Statistical analyses (analy- sis of variance, ANOVA) of the results showed that KCl, MnSO4 andFeSO4·6H2O were important components and that their interactions were strong. Lipopeptide production was predicted to reach 709.87 mg/L after a 60 h incubation using an optimum fermentation medium composed of glucose 7.5 g/L, peanut oil 1.25 g/L, MgSO4 0.37 g/L, KH2PO4 0.75 g/L, monosodium glutamate 6.75 g/L, yeast extract and NH4Cl (5:3 w/w) 10 g/L, KCl 0.16 g/L, FeSO4·6H2O 0.24 mg/L, MnSO4 0.76 mg/L, and an initial pH of 7.0. Lipopeptide production (706.57 ± 3.70 mg/L) in the optimized medium confirmed the validity of the predicted model.
Keywords: Lipopeptides, Bacillus subtilis, optimization, response surface methodology
*Corresponding author
Tel: +86-025-84396212, Fax: +86-025-84396212 E-mail: [email protected]
tion of lipopeptides, a lack of knowledge regarding the sophisticated interactions among various factors leads to complexities and uncertainties in the fermentation process.
Response surface methodology (RSM) is a powerful technique for simultaneously testing multiple variables by statistical experimental design and identifying and quantify- ing interactions between such variables [4]. Therefore, it has been increasingly used for various phases of optimiz- ing the fermentation process [4, 13, 20].
In the present study, we adopted a one-factor-at-a-time experiment to screen carbon and nitrogen sources for lipopeptides production by Bacillus subtilis N7. Further- more, the Plackett–Burman design technique was applied to screen ten factors for the ones that most inuence the pro- duction of lipopeptides. Subsequently, important factors were optimized using the central composite design tech- nique.
Materials and Methods
Microorganism and culture conditions
The lipopeptide producing strain B. subtilis N7 was iso- lated from the cucumber rhizosphere, whose 16s rRNA gene sequence shows 99% identity to Bacillus subtilis DSM 10 based on a BLAST search against all nucleotide sequences in the NCBI database. This strain was stored in nutrient agar (NA) medium at 4oC, and the sequence of the N7 strain was deposited into GenBank with the accession number JQ317780. To prepare the seed culture, a loop of N7 cells from an NA slant culture of fresh NA was inocu- lated into a 250 ml flask containing 50 ml Luria–Bertani (LB) broth; the flask was shaken for 60 h at 240 rpm and 24 h at 200 rpm on a rotary shaker at 30oC and used as a seed culture for subsequent submerged cultivations. Batch fer- mentation was carried out with 170 rpm orbital agitation for 60 h at 30oC in 250 ml asks containing 50 ml of fermenta- tion medium inoculated with 1% v/v of seed culture. Basal medium (glucose 5 g/L, peptone 10 g/L, NaCl 5 g/L, pH 7.0) was used for carbon and nitrogen source selection. The original fermentation medium (pH 7.0) consisted of 1% car- bon source (glucose, sucrose, starch or potato), 0.5% nitro- gen source (yeast extract, peptone, beef extract, trypsin, (NH4)2SO4, NH4Cl and KNO3), sodium glutamate 5 g/L, peanut oil 1 g/L, KH2PO4 1 g/L, MgSO4 0.5 g/L, KCl 0.5 g/L, MnSO4 0.5 mg/L, FeSO4·7H2O 0.15 mg/L, and CuSO4
·5H2O 0.15 mg/L.
All the experiments were carried out independently in triplicate, and the results were the average of three repli- cate experiments.
Analysis of samples
The production of the lipopeptides was measured using the a previously-reported method [16] with slight modica- tion. High-performance liquid chromatography (HPLC) for quantitative determination of lipopeptides was performed on an Agilent Technologies 1200 series system (Agilent Co., Santa Clara, CA, USA) composed of an Eclipse XDB- C18 column (4.6 mm × 250 mm, 5 µm). The injection vol- ume was 20 µl. The sample was eluted with a mobile phase of 3.8 mM triuoroacetate/acetonitrile (3:2, v/v) at a flow rate of 0.8 ml/min. The chromatogram was monitored at 280 nm. Iturin A and surfactin standards obtained from Sigma-Aldrich Chemical (st. Louis, MO, USA) to construct calibration curves from which lipopeptide concentration in the fermentation media was determined.
Experimental design and data analysis
A one-factor-at-a-time approach and a two-way classifi- cation design [9] were used to screen suitable carbon and nitrogen sources. Experiments were conducted in triplicate, and the data were analyzed using SPSS Version 16.0 soft- ware (SPSS Inc., Chicago, IL, USA).
To identify the important variables for lipopeptides pro- duction, different medium components were evaluated using PBD The total number of trials to be carried out according to PBD is k+1, where k is the number of vari- ables. Each variable was studied at two levels, high and low, denoted by (+1) and (-1) signs, respectively (Table 1).
One dummy variable was introduced into the PBD matrix, which was used to calculate the standard error (SE) as fol- lows:
(1)
Where Ed is the effect of the dummy variable and n is the SE
∑
( )Ed 2---n
=
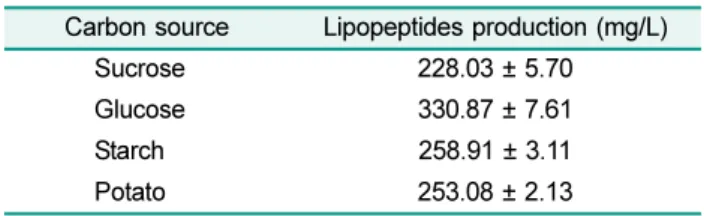
Table 1. Carbon source selection for lipopeptide production.
Carbon source Lipopeptides production (mg/L)
Sucrose 228.03 ± 5.70
Glucose 330.87 ± 7.61
Starch 258.91 ± 3.11
Potato 253.08 ± 2.13
number of dummy variables. In our experiments, the vari- ables with confidence levels above 95% were considered as significantly influencing lipopeptides production. All the experiments were conducted in triplicate. Experimental design and statistical analyses were performed using Minitab 15 Statistical Software® (Minitab Inc., PA, USA).
RSM was used to optimize the most signicant variables identied by PBD. The three independent variables were studied at ve different levels (-α, -1, 0, +1, +α, where α = (2k)1/4 and k is the number of variables). The role of each variable, their interactions, and statistical analysis to obtain predicted lipopeptides production is explained by applying the following quadratic equation:
(2) where Y is the predicted response, β0 is an offset term, βi is the linear effect, βii is the squared effect, βij is the interaction effect, and Xi is the dimensionless coded value of xi. The statistical software package “Design Expert Version 7.0”
(Stat-Ease Inc., Minneapolis, MN, USA) was used for ana- lyzing the experimental data.
Shake-flask Fermentation of B. subtilis N7 using opti- mum medium
The standard seed culture was inoculated into a 500 ml Erlenmeyer flasks with 100 ml optimum medium (glucose
7.5 g/L, peanut oil 1.25 g/L, MgSO4 0.37 g/L, KH2PO4 0.75 g/
L, monosodium glutamate 6.75 g/L, yeast extract and NH4Cl (5:3, w/w) 10 g/L, KCl 0.16 g/L, FeSO4·6H2O 0.24 mg/L, MnSO4 0.76 mg/L, and initial pH 7.0). Batch fer- mentation was carried out with 170 rpm orbital agitation for 60 h at 30oC for 84 h. The profile of cell growth of B. subtilis N7 and lipopeptieds production were detected by regular time interval.
All the experiments were carried out independently in triplicate, and the results were the average of three repli- cate experiments.
Results
Carbon and nitrogen source selection
Pre-experiments were done to investigate the influence of various carbon and nitrogen sources on lipopeptide produc- tion by B. subtilis N7. In the carbon selection experiment, each trial used basal medium, except that glucose was replaced by carbon the sources listed in Table 1. The results showed glucose was superior to other carbon sources. Both the inorganic nitrogen source (A) and organic nitrogen source (B) (Table 2) were evaluated with two-way classification design (Marks 1968). Table 3 pre- sents the ANOVA results of the pre-experiments. The results showed that both inorganic and organic nitrogen Y β= 0+
∑
βiXi+∑
βijXiXj+∑
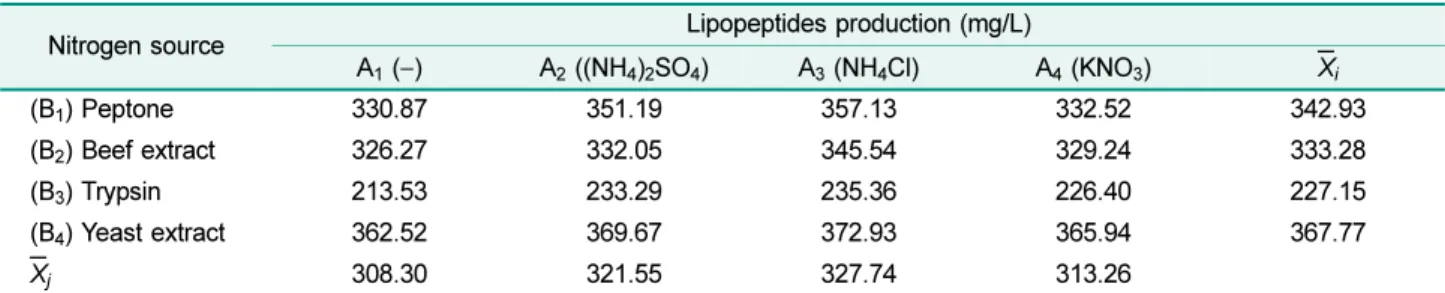
βijXi2Table 2. Selection of nitrogen source and results in lipopeptide production.
Nitrogen source Lipopeptides production (mg/L)
A1 (−) A2 ((NH4)2SO4) A3 (NH4Cl) A4 (KNO3) Xi
(B1) Peptone 330.87 351.19 357.13 332.52 342.93
(B2) Beef extract 326.27 332.05 345.54 329.24 333.28
(B3) Trypsin 213.53 233.29 235.36 226.40 227.15
(B4) Yeast extract 362.52 369.67 372.93 365.94 367.77
Xj 308.30 321.55 327.74 313.26
The concentration of each nitrogen source was 5 g/L organic nitrogen and 3 g/L inorganic nitrogen, except for the first group that had no inorganic nitrogen source.
A: Inorganic nitrogen source, B: organic nitrogen source, xi: mean value of row, xj: mean value of column.
Table 3. ANOVA of nitrogen source selection in lipopeptides production.
Source Type III Sum of
Squares df Mean Square F Sig.
A 12780.405 3 926.802 80.805 p<0.05
B 139108.22 3 46369.407 4.043E3 p<0.05
A * B 524.641 9 58.293 5.082 p<0.05
Error 367.026 32 11.470
Table 4. Range of different variables for the Plackett- Burman design.
Code Variables Level
-1 +1
X1 Glucose (g/L) 7.5 12.5
X2 Peanut oil (g/L) 0.75 1.25
X3 MgSO4 (g/L) 0.37 0.62
X4 KCl (g/L) 0.37 0.62
X5 KH2PO4 (g/L) 0.75 1.25
X6 FeSO4·6H2O (mg/L) 0 0.02
X7 MnSO4 (mg/L) 0.37 0.62
X8 Sodium glutamate (g/L) 3.75 6.75 X9 Yeast extract and NH4Cl (g/L) 6 10
X10 CuSO4·5H2O (mg/L) 0 0.02
X11 Dummy - -
Variable X9 is a combination of yeast extract and NH4Cl in the ratio of 5:3.
Table 5. Plackett-Burman experimental design matrix for screening of important variables for lipopeptides production.
Experiment
no. X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 X11 Lipopeptides
production (mg/L)
1 1 -1 1 -1 -1 -1 1 1 1 -1 1 481.92
2 1 1 -1 1 -1 -1 -1 1 1 1 -1 181.47
3 -1 1 1 -1 1 -1 -1 -1 1 1 -1 228.81
4 1 -1 1 1 -1 1 -1 -1 -1 1 1 244.59
5 1 1 -1 1 1 -1 1 -1 -1 -1 -1 307.71
6 1 1 1 -1 1 1 -1 1 -1 -1 1 394.50
7 -1 1 1 1 -1 1 1 -1 1 -1 -1 670.65
8 -1 -1 1 1 1 -1 1 1 -1 1 1 276.15
9 -1 -1 -1 1 1 1 -1 1 1 -1 1 347.16
10 1 -1 -1 -1 1 1 1 -1 1 1 -1 631.20
11 -1 1 -1 -1 -1 1 1 1 -1 1 1 689.55
12 -1 -1 -1 -1 -1 -1 -1 -1 -1 -1 -1 165.69
Table 6. Regression analysis of Plackett-Burman design.
Variables T value P> t Variables T value P> t intercept 116.75 0.005 X6 34.83 0.018
X1 -4.91 0.128 X7 38.80 0.016
X2 9.63 0.066 X8 4.54 0.138
X3 -2.15 0.277 X9 10.06 0.063
X4 -15.57 0.041 X10 -1.39 0.396
X5 -7.69 0.082 sources had very significant effects on lipopeptide produc-
tion; however, NH4Cl was more effective at increasing lipopeptide production than were other inorganic nitrogen sources. Furthermore, lipopeptide production of the B4 group was higher than that of other groups. We also found that the addition of both NH4Cl and yeast extract to the medium had a synergistic effect on lipopeptide production above their independent individual contributions.
Plackett-Burman experimental design
PBD was used to identify which variables had a signifi- cant inuence on lipopeptide production. Eleven variables,
including 10 medium components in the original fermenta- tion medium and 1 dummy variable, were screened in PB experiments (Table 4). Table 5 shows the PB experimental design for 20 trials with two levels of each variable and their corresponding effects on lipopeptide production. Confi- dence levels were accepted only when above 95%
(p<0.05). Based on statistical analysis, three variables were signicant in terms of lipopeptide production: X4 (KCl), X6 (FeSO4·6H2O), andX7 (MnSO4). Among them, FeSO4· 6H2O and MnSO4 had positive effects, while KCl had a negative one. Although the independent effects of X2 (pea- nut oil) and X9 (yeast extract and NH4Cl) were insignificant, they were chosen for use at their high levels based on their positive effects and secondary role in lipopeptides produc- tion. In our PB experiments, the effects of X3 (MgSO4) and X10 (CuSO4·5H2O) were neither positive nor significant to lipopeptide production by B. subtilis N7. Hence, these two variables were not considered for use as medium compo- nents. Concentrations of the variables (excluding the three significant variables) in the fermentation medium were as follows: glucose 7.5 g/L, peanut oil 1.25 g/L, KH2PO4
0.75 g/L, sodium glutamate 3.75 g/L, yeast extract and NH4Cl (5:3, w/w) 10 g/L.
Central composite design
X4 (KCl), X6 (FeSO4·6H2O) andX7 (MnSO4) were se- lected and further optimized using CCD. Each variable was studied at five coded levels (-1.68, -1, 0, 1, and 1.68), and all variables were taken at a central coded value of zero.
Table 7 shows the experimental design matrix for optimiza- tion of lipopeptide production. By applying quadratic regres- sion analyses (ANOVA) on the experimental data, results of the CCD were tted to a second-order polynomial equation as follows:
Y = 699.27− 46.33A + 4.23B + 39.43C - 94.68AB + 48.78AC − 25.66BC − 113.98A2- 43.63B2
− 107.97C2 (3)
where Y represents lipopeptides production (mg/L), and A, B and C represent KCl, FeSO4·6H2O and MnSO4·H2O respectively.
ANOVA results for the model are summarized in Table 8.
The results of the ANOVA showed that the model is signifi- cant. Furthermore, the lack of fit value of 0.0927 implies that the lack of fit is not significant relative to the pure error.
The coefcient of determination (R2) for lipopeptide produc- tion was calculated to be 0.9946, indicating that 99.46% of the total variation was explained by the model. The plot of predicted values vs. experimental values in Fig. 1 also shows that all the predicted values of the RSM model were close to the experimental values. The adequate precision ratio, which measures the signal-to-noise ratio, was calcu-
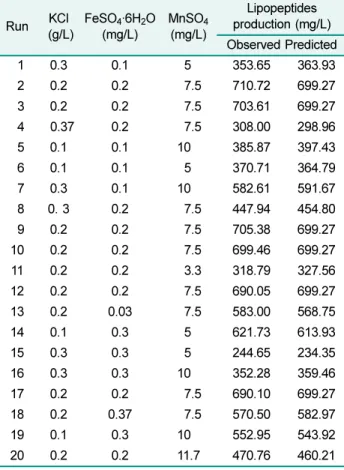
Table 7. Experimental design matrix for optimization of lipopeptide production using CCD.
Run KCl (g/L)
FeSO4·6H2O (mg/L)
MnSO4 (mg/L)
Lipopeptides production (mg/L) Observed Predicted
1 0.3 0.1 5 353.65 363.93
2 0.2 0.2 7.5 710.72 699.27
3 0.2 0.2 7.5 703.61 699.27
4 0.37 0.2 7.5 308.00 298.96
5 0.1 0.1 10 385.87 397.43
6 0.1 0.1 5 370.71 364.79
7 0.3 0.1 10 582.61 591.67
8 0. 3 0.2 7.5 447.94 454.80
9 0.2 0.2 7.5 705.38 699.27
10 0.2 0.2 7.5 699.46 699.27
11 0.2 0.2 3.3 318.79 327.56
12 0.2 0.2 7.5 690.05 699.27
13 0.2 0.03 7.5 583.00 568.75
14 0.1 0.3 5 621.73 613.93
15 0.3 0.3 5 244.65 234.35
16 0.3 0.3 10 352.28 359.46
17 0.2 0.2 7.5 690.10 699.27
18 0.2 0.37 7.5 570.50 582.97
19 0.1 0.3 10 552.95 543.92
20 0.2 0.2 11.7 470.76 460.21
Table 8. Variance analysis for the regression equation.
Source Sum of
Squares df Mean Square
F Value
p-value Prob > F Model 478171 9 53130.11 310.9238 < 0.0001 A-KCl 29316.36 1 29316.36 171.5629 < 0.0001 B-FeSO4·6H2O 244.1839 1 244.1839 1.428994 0.2595 C-MnSO4 21237.56 1 21237.56 124.2848 < 0.0001 AB 71712.53 1 71712.53 419.6703 < 0.0001 AC 19032.98 1 19032.98 111.3833 < 0.0001 BC 5266.972 1 5266.972 30.82295 0.0002 A2 187231.5 1 187231.5 1095.702 < 0.0001 B2 27436.04 1 27436.04 160.559 < 0.0001 C2 168000.8 1 168000.8 983.1612 < 0.0001 Residual 1708.782 10 170.8782
Lack of Fit 1337.914 5 267.5827 3.607514 0.0927 Pure Error 370.8686 5 74.17372
Cor Total 479879.8 19
AB, AC, and BC represent the interaction effect of variables A, B, and C; A2, B2, and C2 are the squared effects of the vari- ables. R2= 0.9964; Adj. R2= 0.9932; CV = 2.52%; adequate precision ratio = 50.298.
Fig. 1. Comparison between the experimental values and the predicted values of the RSM model.
lated as 50.298. Because a ratio greater than 4 is desirable, the ratio of 50.298 indicates an adequate signal. A relatively lower value of the coefcient of variation (CV=2.52%) indi- cated good precision and reliability of the experiment. The results showed KCl and MnSO4 had significant indepen- dent effects and a synergistic effect on protein production.
Although the independent effect of FeSO4·6H2O was insig- nificant, its squared effect and interaction effects with other
variables were significant.
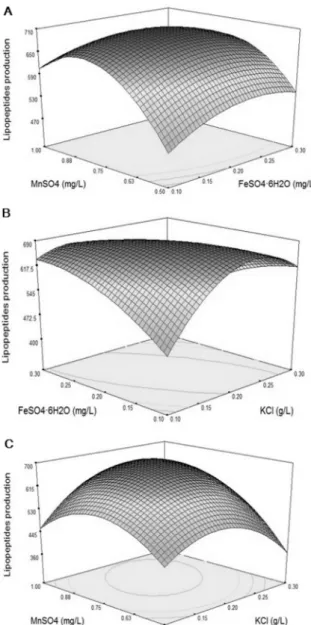
Three-dimensional response plots in Fig. 2 were created to show the interactions of medium components and find the optimum range of the components required for maxi- mum lipopeptide production. The shapes of contour plots indicate the nature and extent of the interactions. The prominent interaction between KCland FeSO4·6H2O (Fig.
2B) is shown by the elliptical nature of the contour plot, while less prominent interactions are shown in Figs. 2A and 2C. It is evident from the response surface plot that maxi- mum lipopeptide production required an increasing concen- tration of MnSO4 and a decreasing concentration of KCl, whereas high concentrations of the MnSO4 variables have a significantly negative influence on the response.
Response surface estimation for maximum production According to Eq. (2), the optimum medium composition was determined to be KCl 0.16 g/L, FeSO4·6H2O 0.24 mg/
L, and MnSO4 0.76 mg/L. The uncoded values of the test variables were as follows: glucose 7.5 g/L, peanut oil 1.25 g/L, MgSO4 0.37 g/L, KH2PO4 0.75 g/L, monosodium glutamate 6.75 g/L, yeast extract and NH4Cl (5:3 w/w) 10 g/
L, initial pH 7.0. The predicted maximum lipopeptide pro- duction was 709.87 mg/L.
Shake-flask fermentation of B. subtilis N7 using opti- mum medium
To confirm the model adequacy for predicting maximum lipopeptide production and suitability for cell growth of B.
subtilis N7 in optimum medium, three replicate experiments using the optimum medium composition were performed.
Fig. 2. Response surface and contour plots of lipopeptide production by B. subtilis N7 showing the effect of two vari- ables (other variables were kept at a fixed coded level).
A: combined effect of MnSO4 and FeSO4·6H2O on the produc- tion of lipopeptides. B: combined effect of FeSO4·6H2O and KCl on the production of lipopeptides. C: combined effect of MnSO4 and KCl on the production of lipopeptides.
Fig. 3. Time course of cell growth and lipopeptides produc- tion of B. subtilis N7.
The time course of the cultivation is shown in Fig. 3. This study showed that when B. subtilis N7 was cultivated at the aforementioned condition, the cell growth occurred before lipopeptides production accumulation. Lipopeptides pro- duction accumulated rapidly during cell growth stationary phase (36-60 h). The average maximum yield of lipopep- tides (706.57 ± 3.70) was obtained after 60 h cultivation, which was 52.6% higher than the original fermentation medium (463.04 g/L). During cells apoptosis period (72- 84 h), the lipopeptides production dramatically decreased by 30.05%.
Discussion
In our study, RSM was used to optimize the medium composition for lipopeptide production by B. subtilis N7.
RSM has proven to be a valuable tool in exploring all the main nutritional factors and to obtain their optimum levels in a process. This method offers a number of advantages; for example, we can determine the effects of variables and select the most influential factors with less experimentation.
Furthermore, we can develop a system model to predict experimental lipopeptide production. After carbon and nitrogen source selections, glucose, yeast extract and NH4Cl, and eight other variables were tested by PB experi- ments. Among these, KCl, FeSO4·6H2O and MnSO4 were identified as the most significant factors for lipopeptides production. Earlier studies found MgSO4·7H2O, CaC12· 2H2O, Na2HPO4, and KH2PO4 are not related to lipopeptide production, but FeSO4·7H2O and MnSO4·H2O supplied at different moladties during stationary growth phase influ- enced lipopeptide production [5]. Bernheimer and Avigad [3] also reported an increase in Mn2+ from 0.33 g/L to 2.6 g/
L improved surfactin production. However, few studies have reported KCl as being a significant factor for lipopep- tide production. Furthermore, the interactions of KCl and FeSO4·6H2O, KCl and MnSO4, as well as FeSO4·6H2O and MnSO4 were found to be significant based on RSM analysis. The effects of KCl seem to be related to perme- ation pressure by offering a buffer environment in cooper- ation with KH2PO4 for cell growth. In shake flask cultures, permeation pressure and broth pH are hard to control; how- ever, maintaining a suitable environment for B. subtilis N7 growth is very important because changes may inhibit bio- mass formation and promote acidic byproduct production in the fermentation process and ion form inactivation [1]. Ear-
lier studies indicate that Mn2+ can affect metabolites of Bacillus by regulating the metabolic flux of glucose and making the microorganism utilize more glucose for forming metabolites instead of biomass [15, 18]. Here, we found that an increase in the concentration of Mn2+ increased lipopeptide production to a maximum value; then, further increases in Mn2+ concentrations inhibited lipopeptide production based on the response surface plot.
Iron is also considered an essential element for cellular growth and other bioenergetic pathways [19]. An iron- enriched culture of B. subtilis ATCC 21332 exhibited the ability to emulsify kerosene and achieved a maximum emulsion index (E24) of 80% for culture supplemented with 4.0 mM Fe2+. In our study, a high concentration of FeSO4 was one of the key factors for high lipopeptide production.
Because several enzymes in amino acid biosynthesis path- ways are iron-dependent, iron limitation may cause amino acid starvation [10] and subsequently influence the gene expression involved in amino acid biosynthesis associated with pathways essential for lipopeptide production.
Furthermore, the addition of peanut oil and the combina- tion of organic with inorganic nitrogen sources contributed to lipopeptide production improvement by RSM, although they were not considered as key factors.
The results of verification experiments indicated that this optimum medium led to a noticeable improvement in lipopeptide production by B. subtilis N7. However, due to the highly complex nature of biological systems, the actual contribution of the significant factors to the mechanisms for enhancing lipopeptide production may be more complicated than we have discussed in this paper, and lipopeptides pro- duction conditions are under further investigation.
Acknowledgments
This work was supported by the National Basic Research Pro- gram of China (Grant no. 2011CB100503) and the National Depart- ment of Public Benefit Research Foundation of the Ministry of Agriculture of China (Grant no. 201103004).
References
1. Amanullah, A., C. McFarlane, A. Emery, and A. Nienow. 2001.
Scale - down model to simulate spatial pH variations in large - scale bioreactors. Biotechnol. Bioeng. 73: 390-399.
2. Asaka, O. and M. Shoda. 1996. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Appl.
Environ. Microbiol. 62: 4081-4085.
3. Bernheimer, A. and L. S. Avigad. 1970. Nature and properties of a cytolytic agent produced by Bacillus subtilis. J. Gen.
Microbiol. 61: 361.
4. Chen, H. C. 1996. Optimizing the concentrations of carbon, nitrogen and phosphorus in a citric acid fermentation with response surface method. Food Biotechnol. 10: 13-27.
5. Cooper, D. G. and J. D. Sheppard. 1991. The response of Bacillus subtilis ATCC 21332 to manganese during continu- ous-phased growth. Appl. Microbiol. Biotechnol. 35: 72-76.
6. Finking, R. and M. A. Marahiel. 2004. Biosynthesis of nonribo- somal peptides 1. Annu. Rev. Microbiol. 58: 453-488.
7. Heerklotz, H. and J. Seelig. 2007. Leakage and lysis of lipid membranes induced by the lipopeptide surfactin. Eur. Bio- physics J. 36: 305-314.
8. Hutadilok-Towatana, N., A. Painupong, and P. Suntinanalert.
1999. Purification and characterization of an extracellular pro- tease from alkaliphilic and thermophilic Bacillus sp. PS719. J.
Biosci. Bioeng. 87: 581-587.
9. Marks, E. 1968. Profile analysis in a two-way classification problem. Multivar. Behav. Res. 3: 95-106.
10. Miethke, M., H. Westers, E. J. Blom, O. P. Kuipers, and M. A.
Marahiel. 2006. Iron starvation triggers the stringent response and induces amino acid biosynthesis for bacillibactin produc- tion in Bacillus subtilis. J. Bacteriol. 188: 8655-8657.
11. Mizumoto, S., M. Hirai, and M. Shoda. 2006. Production of lipopeptide antibiotic iturin A using soybean curd residue culti- vated with Bacillus subtilis in solid-state fermentation. Appl.
Microbiol. Biotechnol. 72: 869-875.
12. Ongena, M. and P. Jacques. 2008. Bacillus lipopeptides: ver- satile weapons for plant disease biocontrol. Trend. Microbiol.
16: 115-125.
13. Pareek, N., R. P. Singh, and S. Ghosh. 2011. Optimization of medium composition for enhanced chitin deacetylase produc- tion by mutant Penicillium oxalicum SAE (M)-51 using response surface methodology under submerged fermenta- tion. Process Biochem. 46: 1693-1697.
14. Quentin, M., F. Besson, F. Peypoux, and G. Michel. 1982.
Action of peptidolipidic antibiotics of the iturin group on eryth- rocytes: Effect of some lipids on hemolysis. Biochimica et Bio- physica Acta (BBA)-Biomembranes. 684: 207-211.
15. Rado, T. A. and J. A. Hoch. 1973. Phosphotransacetylase from Bacillus subtilis: purification and physiological studies.
Biochimica et Biophysica Acta (BBA)-Enzymology. 321: 114- 125.
16. Shih, I.-L., C.-Y. Lin, J.-Y. Wu, and C. Hsieh. 2009. Production of antifungal lipopeptide from Bacillus subtilis in submerged fermentation using shake flask and fermentor. Korean J.
Chem. Eng. 26: 1652-1661.
17. Stein, T. 2005. Bacillus subtilis antibiotics: structures, synthe- ses and specific functions. Mol. Microbiol. 56: 845-857.
18. Ujita, S. and K. Kimura. 1982. Glucose-6-phosphate dehydro- genase, vegetative and spore Bacillus subtilis. Methods in Enzymol. 89: 258-261.
19. Wei, Y. H., L. F. Wang, and J. S. Chang. 2004. Optimizing iron supplement strategies for enhanced surfactin production with Bacillus subtilis. Biotechnol. Progr. 20: 979-983.
20. Wu, Q.-L., T. Chen, Y. Gan, X. Chen, and X.-M. Zhao. 2007.
Optimization of riboflavin production by recombinant Bacillus subtilis RH44 using statistical designs. Appl. Microbiol. Bio- technol. 76: 783-794.