177 책임저자:김형식, 609-735, 부산시 금정구 장전동 산 30
부산대학교 약학대학 분자독성학교실 Tel: 051-510-2816, Fax: 051-582-2816 E-mail: [email protected]
접수일:2008년 7월 28일, 게재승인일:2008년 8월 11일
Correspondence to:Hyung Sik Kim
Division of Molecular Toxicology, College of Pharmacy, Pusan National University, San 30, Jangjeon-dong, Geumjeung-gu, Pusan 609-735, Korea
Tel: +82-51-510-2816, Fax: +82-51-582-2816 E-mail: [email protected]
Molecular Mechanism of Progesterone-Induced Apoptosis in Human Breast Cancer T47D Cells
Ji Young Im1, Tae Hyung Kim1, Young Jun Lee1, In Young Kim2, Seung Jun Kwack2, Byung Mu Lee3, Chang Sun Myung4 and Hyung Sik Kim1
1Laboratory of Molecular Toxicology, College of Pharmacy, Pusan National University, Busan 609-735,
2National Institutes of Toxicological Research, Korea Food and Drug Administration, Seoul 122-704,
3College of Pharmacy, Sungkyunkwan University, Suwon 440-746, 4Department of Pharmacy, Chungnam National University, Yuseong, Daejeon 305-764, Korea
Progesterone plays an essential role in the development and differentiation of the mammary gland.
Although it is well established that progesterone regulates cellular proliferation and differentiation, the molecular mechanisms of progesterone-mediated anticancer effects remain unclear. Therefore, the aim of the present study was to determine how progesterone regulates the growth and viability of T47D human breast cancer cells. In this study, the proliferation of T47D cells was inhibited after 48 hours of incubation in the presence of 10μM progesterone. The progesterone-treated T47D cells also exhibited hallmarks of apoptosis including DNA fragmentation and nuclear morphological changes detected with DAPI staining. Also, PR-A and PR-B progesterone receptor expression was decreased in a dose-dependent manner. Moreover, progesterone significantly decreased the expression of p53, cyclin D1, and CDK4, while it increased the expression of p27. These results clearly demonstrate high doses of progesterone induce T47D human breast cancer cell apoptosis and suppress proliferation by inhibiting the expression of key cell cycle regulators. (Cancer Prev Res 13, 177-183, 2008)
Key Words: Progesterone, Apoptosis, T47D cells, p53, Cyclin D1
INTRODUCTION
Progesterone is an important regulator of the growth and differentiation of normal and malignant breast epithelial cells, and is used for hormone-related breast tumor therapy.1,2) However, how progesterone elicits these antiproliferative effects is poorly understood. The mechanism of progesterone-mediated inhibition of breast cancer cell growth is controversial. Several studies have indicated that the inhibition of cancer cell growth by progesterone is closely associated with genes that regulate cell cycle arrest and stimulate apoptosis.3∼5) However, these studies present conflicting data. For example the suppression of in vitro breast cancer cell proliferation by progesterone has
been linked to an upregulation of p53 expression.6∼9) In contrast, Moore et al. reported that the R5020 synthetic progestin significantly decreased p53 expression levels.7) The evidence discussed here supports the conclusion that the role of cell cycle and apoptosis regulatory proteins in progesterone-mediated inhibition of breast cancer cell growth is controversial. It is likely that multiple factors regulate the relative expression levels of these cell cycle and apoptosis regulators and that the balance between their activity levels determines tumor cell growth rate.
Progesterone also stimulates proliferation of breast tissue, and this effect is dependent upon the progesterone receptor (PR). The antiprogestin, RU486, blocks PR signaling and the proliferative effect of synthetic progestin.10) Generally, proges-
terone acts by direct binding to two PR isoforms, PR-A and PR-B, which have different transcriptional activities.11,12) Various model systems have been employed to study the mechanisms of progesterone signaling. The T47D cell line is a widely used model system because it has high PR expression and high progesterone sensitivity.13) This model has been used to identify many progesterone target genes that are upregulated by progesterone treatment and contribute to growth inhibition including p21, p27, and p53.14) Progestin treatment of T47D breast cancer cells also results in the inhibition of cyclin D1-Cdk4, cyclin D3-Cdk4, and cyclin E-Cdk2 kinase activities, and reduced phosphorylation of pRB.15) The T47D model shows a biphasic change in the rate of cell cycle progression in response to treatment with synthetic progestins. Thus, the opposing effects of progesterone on cell cycle regulation can be observed in this model, demonstrating the complexity of progestin effects on breast cancer cell proliferation.
Dinda et al6) reported that T47D cells that were cultured in serum-containing medium maintained high levels of p53 expression, whereas T47D cells that were cultured in charcoal- treated serum had minimal levels of p53.16) It is now widely recognized that p53 is the most frequently mutated protein in human cancers and controls tumor cell growth by regulating cell cycle progression and apoptosis.17) Progesterone-induced apoptosis is closely related to induction of p53 tumor suppressor protein expression in T47D cells.24,25) Therefore, we explored the possibility that the progesterone-induced antiproliferative effect on T47D human breast cancer cells is due to its ability to induce apoptosis. To identify the molecular mechanism linking progestin to the inhibition of cell cycle progression, we have examined the effects of progestin treatment of T47D human breast cancer cells on the expression of p53, cyclin D1, and CDK4. The results of this study demonstrate that proges- terone not only regulates cyclin abundance, but also decreases the expression of p53, providing a mechanism for the growth- inhibitory effect of progesterone in breast cancer cells.
MATERIALS AND METHODS 1. Chemicals
Dimethylsulfoxide (DMSO) and progesterone were purchased from Sigma Chemical Co. (St. Louis, MO). Dulbecco’s Modified Eagle Medium (DMEM), fetal bovine serum (FBS), penicillin- streptomycin, and 0.25% trypsin-EDTA were purchased from
Gibco BRL (Grand Island, NY). Non-fat dry milk was purchased from Bio-Rad Co. (Hercules, CA). Progesterone receptor (PR) antibody (polyclonal) was purchased from Zymed Laboratories Inc. (San Francisco, CA). The Enhanced Chemilu- minescence System (ECL plus) was purchased from Amersham Pharmacia Biotech (Buckinghamshire, UK). All stock solutions were stored at 4°C and diluted to the desired concentration in phenol red-free medium for testing. The final DMSO concentration in the medium did not exceed 0.2%.
2. Cell Culture Conditions
The human breast cancer T47D cells were purchased from ATCC (Bethesda, MD). The cells were maintained at 37°C in a humidified atmosphere with 95% air and 5% CO2 in DMEM, 10% FBS, and 1% penicillin/ streptomycin. The culture medium was changed routinely every two days. For experiments, cell were plated at 80% confluence in phenol red-free DMEM containing 5% charcoal-dextran (CD) stripped- FBS for 24 h to minimize estrogenic activity [18].
3. Cell Proliferation Assay
This assay was carried out as previously described.10) About 1,500 cells per well in 100μl of media were plated into a 96-well microtiter plate. Two days later, the growth media was removed and replaced with experimental medium containing the test chemicals. The experimental medium consisted of phenol red-free DMEM supplemented with CD-treated FBS.
The culture was continued for two days and then the 3-(4, 5- dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay was conducted to measure cell proliferation. Cells were treated with MTT (Sigma) solution (5 mg/ml) and further incubated for 4 h. After 100μl of the supernatant was replaced with the same volume of DMSO, the absorbance was read at 570 nm with a micro-ELISA reader (DI Biotech, Winooski, VT). ‘Percent of cell survival’ was defined as the relative absorbance of treated versus untreated cells.
4. DAPI Staining
The cells (5×104) were seeded into 35 mm diameter six-well plates containing 5% CD-FBS for 48 h. The cells were then treated with progesterone (1 and 10μM) in the presence of 5% FBS. The cell morphology was examined. Then the cells were washed with PBS, fixed with 100% methanol, and stained with 4', 6-diamidino-2-phenylindole (DAPI, Sigma, 1μg/ml).
Fig. 1. Dose-dependent effect of progesterone on the prolife- ration of T47D cells. T47D cell were incubated with various concentrations of progesterone in Charcoal-dextran stripped 5% FBS for 2 and 4 days, respectively. Values are mean±SEM of duplicates from three independent experiments.
Fig. 2. Time-course effect of progesterone on the proliferation of T47D cells. T47D cell were incubated with various concent- rations of progesterone in Charcoal-dextran stripped 5% FBS for 2 and 4 days, respectively. Values are mean±SEM of duplicates from three independent experiments.
The stained nuclei were visualized with a Zeiss Axioplan fluo- rescence microscope equipped with a DAPI filter.
5. Flow Cytometry Analysis
The T47D cells were exposed to apicidin for 48 h. The suspension and adherent cells, were collected, washed, and resuspended in cold PBS. The cells were fixed in chilled 75%
methanol and stained with a propidium iodine (PI) solution (100μg/ml RNase, 10μg/ml PI in phosphate buffered saline (PBS)). Data acquisition and analysis was performed using a flow cytometry system (Becton Dickinson, San Jose, CA).
6. Western Blot Analysis
The T47D cells were plated at a density of 2.5×105 cells per 60 mm dish and incubated for 24 h. The medium was then changed to phenol red-free DMEM supplemented with 5% CD FBS. After a 48 h incubation, the cells were treated with 1 and 10μM of progesterone and incubated for an additional 48 h. They were then washed twice with cold PBS and lysed in 250μl of lysis buffer (0.5% Triton X-100, 0.15 M NaCl, 50 mM Tris-HCl, pH 7.4, 25 mM NaF, 20 mM EGTA, 1 mM DTT, 1 mM Na3VO4) containing protease inhibitor cocktail. Equal amounts of total protein were resolved on a 10% SDS-polyacrylamide gel and transferred to nitrocell- ulose membranes. The membranes were blocked with 5%
non-fat dry milk in PBS containing 0.1% Tween-20 (PBST).
After briefly washing with PBST, the levels of PR and p53 were detected using a 1:1500 dilution of HRP-conjugated secondary antibody. Immunoreactive bands were visualized using ECL plus. Relative band intensities were determined using an Image Analyzer.
7. Statistics
Values are presented as the mean ± standard deviation (SD).
Statistical analysis was performed using the one-way analysis of variance (ANOVA) and the Dunnett’s tests. p values of <
0.05 were considered to be statistically significant.
RESULTS AND DISCUSSION
Progesterone is a key regulator of female reproductive functions including embryonic implantation, the maintenance of pregnancy, and the proliferation of breast tissue.19) A number of studies have demonstrated that progesterone acts by
binding to the PR to mediate its antagonist effects on cellular proliferation and regulate the expression of the progesterone- responsive proteins.20,21) To determine if progesterone enhanced T47D breast cancer cell proliferation, the cells were cultured in stripped serum with or without progesterone and the proliferation rate was monitored. The dose dependent response of T47D cells to progesterone is shown in Fig. 1. The proliferation of T47D cells was estimated after two or four days of treatment with progesterone (10−12 to 10−4 M). Proges- terone dose-dependently stimulated the cell proliferation by
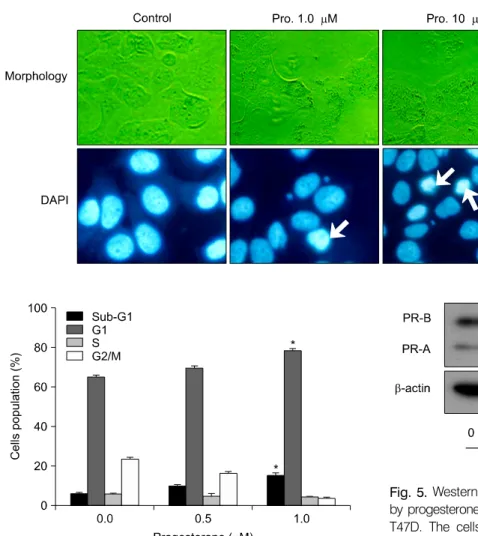
Fig. 3. Characterization of proges- terone-induced apoptotic cell death. T47D cells were cultured without (control) or with proges- terone (1 and 10μM) for 48 hr.
Yellow arrows indicate condensed nuclei by DAPI staining.
Fig. 4. Effect of progesterone on cell cycle analysis. The T47D cells were treated with the indicated concentrations of progesterone for 48 h. The cells stained with propidium iodine (PI) were subjected to flow cytometry analysis to determine the cell distributions at sub-G1 phase. The data is expressed as the mean±SD of two independent experiments.
Fig. 5. Western blot analysis of progesterone receptors (PRs) by progesterone treatment in the human breast cancer cell line T47D. The cells were incubated with 1 and 10μM proges- terone for 2 days without (control) or with 5% charcoal-dextran (CD)-treated FBS. The total cell lysates were prepared and the protein was subjected to SDS-PAGE followed by Western blot analysis and chemiluminescent detection. Representative bands from three independent experiments are shown.
approximately 1.49 fold, with an optimal concentration of 10−10 M progesterone. Concentrations above 10−8 M resulted in a decreased cell growth. The 50% inhibitory concentration (IC50) for cell proliferation was estimated to be 5×10−8 M. Therefore, the optimal working range of progesterone for studying apoptotic mechanisms is between 1 and 10μM. The time- dependent effect of progesterone on T47D cell proliferation was determined at 1 or 10μM progesterone for five days. In the presence 1μM progesterone, there was no significant inhibition of T47D proliferation throughout the monitored growth period. In contrast, T47D cell growth was significantly inhibited by 10μM progesterone (Fig. 2).
The morphological changes induced by progesterone were
examined by phase-contrast microscopy. T47D cells showed irregular shapes and cytoplasmic blebbing in the presence of 10μM progesterone. Cells were observed via fluorescence microscopy following treatment with DAPI, which specifically stains the nuclei. Progesterone (10μM) significantly increased nuclear condensation and the appearance of apoptotic bodies (Fig. 3). To further confirm the induction of apoptosis by progesterone in T47D cells, flow cytometry analysis was performed progesterone treatment. As shown in Fig. 4, a dose dependent increase in the sub-G1 population resulting from DNA fragmentation was observed. In order to determine the mechanism of inhibition of T47D cell proliferation by proges- terone involved PRs, PR protein levels were measured by Western blot analysis. Cytosolic lysate was prepared from T47D cells treated with progesterone. The baseline PR protein
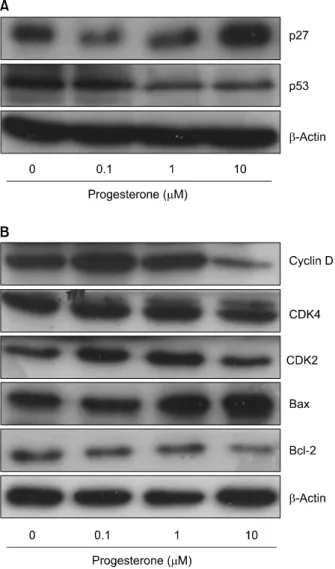
Fig. 6. Western blot analysis of cell cycle regulators cyclin D1, CDK4, CDK2, Bcl-2, and Bax proteins levels by progesterone in T47D cells. The cells were incubated with 0.1, 1.0 or 10 μM progesterone for 48 h without (control) or with 5%
charcoal-dextran (CD)-treated FBS. The total cell lysates were prepared and the protein was subjected to SDS-PAGE followed by Western blot analysis and chemiluminescent detection. The control cells were treated with the vehicle alone.
Representative bands from three independent experiments are shown.
expression in untreated control T47D cells was defined as 100%. Progesterone dose-dependently decreased the cytosolic level of PR protein expression to undetectable levels with the highest dose of 10−5 M (Fig. 5).
Previous studies demonstrated that progesterone-induced apoptosis is closely related to induction of p53 tumor suppressor protein expression in T47D cells.24,25) Based on these results, we examined the effects of progesterone on the expression levels of p53 in T47D cells. Fig. 6A shows that progesterone caused
a down-regulation of p53 protein expression in a dose-depen- dent manner. To show that down-regulation of p53 was not due to a general effect on protein expression, the same membrane was re-probed with β-actin antibody. There was no altered expression of β-actin levels by progesterone in T47D cells. These results suggest that the growth inhibition induced by progesterone is p53-independent in T47D cells.
The levels of cyclin D1 and CDK4 were also significantly decreased (Fig. 6B). In contrast, progesterone markedly increased the level of p27 expression. These data suggest that progesterone may cause T47D growth inhibition by upregulating p27 and downregulating cyclin D1/ CDK4. This mechanism is consistent with reports that progesterone regulates cell cycle regulatory proteins in a variety of other tissues.9,26)
There is considerable interest in elucidating the factors that regulate progesterone-responsive tumor cell differentiation, growth, and apoptosis.27,28) This study showed that progesterone exhibits an immediate stimulation of T47D breast cancer cell proliferation. This is consistent with demonstration by Groshong et al. that progesterones have a biphasic action on the cell cycle.29) Acute progesterone treatment stimulated cell growth through a first mitotic cell cycle, whereas chronic treatment with progesterone inhibited growth in the G1 phase of the second cycle. The proliferative effects of progesterone in breast tissue are dependent upon PR signaling. The T47D cell line is a suitable model system to test PR-mediated gene regulation because this cell line contains high levels of PR but low levels of estrogen receptor (ER).13,27) Therefore, we measured PR protein levels in progesterone treated T47D cells to investigate the mode of action for cell cycle regulation by progesterone. The observed inhibition of PR expression in response to progesterone, suggests that progesterone may inhibit T47D cell proliferation by inhibiting PR expression and consequently, PR-dependent gene expression. Progesterone is also known to regulate the expression of cell cycle regulatory proteins in a variety of tissues.9,28) Specifically, cyclin A and B have been shown to be down-regulated by progestins in T47D cells upon prolonged progestin exposure, which have been linked to its growth- inhibitory effects. P53 has been shown to be able to down- regulate both cyclin A and B. Therefore, it will be very interesting to investigate whether these proteins are also regulated through a p53-like activity in T47D cells.
CONCLUSION
Many in vitro cells culture systems have been developed to assess the anti-tumor effects of novel drugs. In comparison, our understanding of the inhibitory effects of naturally occurring progesterone on various cancer cells is comparatively limited.
Based on the results, it was proposed that the ability of progesterone to inhibit T47D cells growth may result from the interaction with other signaling pathways in this cell line. In summary, high doses of progesterone suppress T47D human breast cancer cells proliferation and induce apoptosis via p53 and cyclin D1 independent mechanisms.
ACKNOWLEDGEMENTS
This study was supported by the grants from the two years (2007∼2008) Research Foundation for Pusan National University and the Brain Korea 21 Project (JY Im, TH Kim and YJ Lee).
REFERENCES
1) Clarke CL, Sutherland RL. Progestin regulation of cellular proliferation. Endocr Rev 11, 266-301, 1990.
2) Santen RJ, Manni A, Harvey H, Redmond C. Endocrine treatment of breast cancer in women. Endocr Rev 11, 221-265, 1990.
3) Alkhalaf M, El-Mowafy AM. Overexpression of wild-type p53 gene renders MCF-7 breast cancer cells more sensitive to the antiproliferative effect of progesterone. J Endocrinol 179, 55-62, 2003
4) Horita K, Inase N, Miyake S, Formby B, Toyoda H, Yoshizawa Y. Progesterone induces apoptosis in malignant mesothelioma cells. Anticancer Res 21, 3871-3874, 2001.
5) Yu S, Lee M, Shin S, Park J. Apoptosis induced by proges- terone in human ovarian cancer cell line SNU-840. J Cell Biochem 82, 445-451, 2001.
6) Dinda S, Kodali-Gali S, Sevilla L, Burkley M, Hurd C, Moudgil VK. Inhibition of proliferation of T47D human breast cancer cells: alterations in progesterone receptor and p53 tumor suppressor protein. Mol Cell Biochem 175, 81-89, 1997.
7) Moore MR, Spence JB, Kiningham KK, Dillon JL. Progestin inhibition of cell death in human breast cancer cell lines. J Steroid Biochem Mol Biol 98, 218-227, 2006.
8) Formby B, Wiley TS. Progesterone inhibits growth and induces apoptosis in breast cancer cells: inverse effects on Bcl-2 and p53. Ann Clin Lab Sci 28, 360-369, 1998.
9) Kester HA, Sonneveld E, van der Saag PT, van der Burg B.
Prolonged progestin treatment induces the promoter of CDK inhibitor p21 Cip1,Waf1 through activation of p53 in human breast and endometrial tumor cells. Exp Cell Res 284, 264-273, 2003.
10) Hissom JR, Bowden RT, Moore MR. Effects of progestins, estrogens, and antihormones on growth and lactate dehydrogenase in the human breast cancer cell line T47D.
Endocrinology 125, 418-423, 1989.
11) Vienonen A, Syvälä H, Miettinen S, Tuohimaa P, Ylikomi T. Expression of progesterone receptor isoforms A and B is differentially regulated by estrogen in different breast cancer cell lines. J Steroid Biochem Mol Biol 80, 307-313, 2002.
12) Kastner P, Krust A, Turcotte B, Stropp U, Tora L, Gro- nemeyer H, Chambon P. Two distinct estrogen-regulated promoters generate transcripts encoding the two functionally different human progesterone receptor forms A and B. EMBO J 9, 1603-1614, 1990.
13) Horwitz KB, Mockus MB, Lessey BA. Variant T47D human breast cancer cells with high progesterone-receptor levels despite estrogen and antiestrogen resistance. Cell 28, 633-642, 1982.
14) Richer JK, Jacobsen BM, Manning NG, Abel MG, Wolf DM, Horwitz KB. Differential gene regulation by the two proges- terone receptor isoforms in human breast cancer cells. J Biol Chem 277, 5209-5218, 2002.
15) Musgrove EA, Lee CS, Cornish AL, Swarbrick A, Sutherland RL. Antiprogestin inhibition of cell cycle progression in T-47D breast cancer cells is accompanied by induction of the cyclin- dependent kinase inhibitor p21. Mol Endocrinol 11, 54-66, 1997.
16) Hurd C, Khattree N, Alban P, Nag K, Jhanwar SC, Dinda S, et al. Hormonal regulation of the p53 tumor suppressor protein in T47D human breast carcinoma cell line. J Biol Chem 270, 28507-28510, 1995.
17) Oren M. p53: the ultimate tumor suppressor gene? FASEB J 6, 3169-3176, 1992.
18) Sonnenschein C, Szelei J, Nye TL, Soto AM. Control of cell proliferation of human breast MCF7 cells; serum and estrogen resistant variants. Oncol Res 6, 373-381, 1994.
19) Graham JD, Clarke CL. Physiological action of progesterone in target tissues. Endocr Rev 18, 502-519, 1997.
20) Gronemeyer H. Transcription activation by estrogen and pro- gesterone receptors. Annu Rev Genet 25, 89-123, 1991.
21) Tsai MJ, O'Malley BW. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu Rev Biochem 63, 451-86, 1994.
22) Franke H, Vermes I. Differential effects of progestogens on breast cancer cell lines. Maturitas 46S1, S55-S58, 2003.
23) Moore MR, Conover JL, Franks KM. Progestin effects on long-term growth, death, and Bcl-xL in breast cancer cells.
Biochem Biophys Res Commun 277, 650-654, 2000.
24) Alban P, Hurd C, Dinda S, Khattree N, Moudgil VK.
Differential regulation of retinoblastoma protein by hormonal and antihormonal agents in T47D breast cancer cells. J Steroid Biochem Mol Biol 77, 135-141, 2001.
25) Hurd C, Khattree N, Dinda S, Alban P, Moudgil VK.
Regulation of tumor suppressor proteins, p53 and retino- blastoma, by estrogen and antiestrogens in breast cancer cells.
Oncogene 15, 991-995, 1997.
26) Musgrove EA, Hunter LJ, Lee CS, Swarbrick A, Hui R, Sutherland RL. Cyclin D1 overexpression induces progestin resistance in T-47D breast cancer cells despite p27 (Kip1) association with cyclin E-Cdk2. J Biol Chem 276, 47675- 47683, 2001.
27) DiLorenzo D, Albertini A, Zava D. Progestin regulation of alkaline phosphatase in the human breast cancer cell line T47D. Cancer Res 51, 4470-4475, 1991.
28) Musgrove EA, Swarbrick A, Lee CS, Cornish AL, Sutherland RL. Mechanisms of cyclin-dependent kinase inactivation by progestins. Mol Cell Biol 18, 1812-1825, 1998.
29) Groshong SD, Owen GI, Grimison B, Schauer IE, Todd MC, Langan TA, Sclafani RA, Lange CA, Horwitz KB. Biphasic regulation of breast cancer cell growth by progesterone: role of the cyclin-dependent kinase inhibitors, p21 and p27 (Kip1).
Mol Endocrinol 11, 1593-1607, 1997.