257
대장균으로부터 생산된 Bacillus licheniformis WL-12의 Cellulase 특성
박종덕·김연아·윤기홍*
우송대학교 식품생물과학과
Bacillus licheniformis WL-12
의carboxymethyl cellulase (cellulase)
유전자를함유한대장균균체파쇄상등액으로 부터DEAE-Sepharose
와Q-Sepharose
컬럼크로마토그래피를통해cellulase
를정제하였다.
정제된효소의비활 성은163 U/mg
이었으며, SDS-PAGE
에의해측정된분자량은약49.5 kDa
으로나타났다. pH 5.5
와55
oC
에서최대 반응활성을보였으며, SDS (5 mM)
에의해서는cellulase
의활성이완전히저해되었고Cu
2+(5 mM)
에의해서는약 간 증진되었다.
정제된cellulase
는CMC, konjac, barley
β-glucan
과lichenan
을가수분해하였으나xylan, locust bean gum
및p -nitrophenyl-
β-glucopyranoside
를 분해하지 못하였다. Cellooligosaccharides
를 정제된WL-12 cellulase
로분해하였을때cellobiose
와cellotriose
가주된최종반응산물로관찰되었으며cellobiose
보다는중합도 가큰cellotriose, cellotetrasoe
와cellopentaose
는분해하였으나cellobiose
는분해하지못하는것으로확인되었다. Key words
□B. licheniformis , cellulase, E. coli , properties, purification
Endo-
β-1,4-glucanase (carboxymethyl cellulase : cellulase)
는exo-
β-1,4-glucanase,
β-glucosidase
와더불어cellulase
계구성효소 로서재생자원인농·임산폐자원의당화,
과일음료와맥주의청징
,
사료첨가물및면직물가공등다양한용도로이용되는산업용효소이다
. Cellulase
는세균,
곰팡이와 더불어식물과 동물에서도생산되고있으며
glycosyl hydrolase (GH) family
를구성하 는 가장 큰 집단이다. Cellulase
의 산업적 생산은Trichoderma
reesei
와같은곰팡이로이루어지고 있으나,
용도에따라서는곰팡이효소가갖는단점을보완하기위해다양한특성을지닌세
균의
cellulase
계효소에 대한관심이 지속되고있다.
청바지 염료의탈색공정에는 곰팡이 효소가산성
pH
에서주로 반응하게 되어 얼룩을 발생시키는 문제가있어 중성pH
에서 작용할수있는세균성
cellulase
의개발이진행되고있으며(4, 9),
리그노셀 룰로즈 바이오매스를cellulase
로 처리하면서Saccharomyces cerevisiae
와Zymomonas mobilis
로동시당화발효시키는데있어 서경제적인문제점으로지적되고있는cellulase
효소의가격을낮추기 위해
Bacillus coagulans
의 중온성cellulase
를 곰팡이cellulase
와 동시에 시용함으로써 바이오매스 당화에 필요한cellulase
효소량을절감하였다(17).
Bacillus licheniformis
는B. subtilis
와더불어산업용 효소의생 산에사용되고있으며,
유전자재조합숙주균으로서산업적이용 이가능한유용GRAS
균주로B. licheniformis DSM3
의총염색 체염기서열이 결정되었다(21). B. licheniformis
는고분자물질의 가수분해효소생산균으로여러종류가다양한장소에서탐색되 어그들이생산하는효소와그유전자에대한특성이다수보고되었다
. B. licheniformis
에서 가장많이연구된 효소로는 내열성 α-amylase
가있으며,
호알칼리성protease (18), cellulase-chitosanase (8). xylanase (5),
β-1,3-1,4-glucanase (15, 20), cellulase (2, 3, 14),
mannanase (7)
와같은섬유물질분해에 관련된효소의 특성이나그유전자가분석되었다
.
또한chitosanase
를생산하는MB-2
가인도네시아온천에서분리된바있으며
(6), 14A
의펙틴분해에관련된효소
(1)
에대한연구도이루어졌다.
한편국내가정에서제조된된장에서고온성 미생물로분리된
B. licheniformis WL-12
의cellulase
는B. licheniformis ATCC 14580
의 효소와 동일한 아미 노산잔기서열을갖는것으로확인되었는데(23),
현재까지그특 성에대한보고가없어본연구에서는대장균에서cellulase
유전 자를발현하여효소를정제하고효소반응특성을검토하였다.
재료 및 방법
사용 균주와 플라스미드 및 배양B. licheniformis WL-12
의cellulase
유전자를과잉발현하기위 한 플라스미드와 숙주균으로pET23a(+)
와E. coli BL21(F
-ompT hadS
B(r
B-m
B-) , gal dcm (DE3))
를각각사용하였으며,
배지 로는ampicillin (100
µg/ml)
을첨가한LB
액체배지(yeast extract, 5 g; tryptone, 10 g; NaCl, 5 g; water, 1 L)
를사용하였다.
DNA 분리와 조작
Cellulase
유전자를증폭하기위해WL-12
의cellulase
유전자를 함유한 재조합플라스미드를주형으로하고유전자의염기서열 에근거하여 제조한12CelF (5 ’ -GGAAAACATATGTCATATATG AAACGTTC-3 ’ : Nde I sites; underlined)
와12CelR (5 ’ -CGTC AAACTCGAGTTATTTAGGTTCAGTG-3 ’ : Xho I sites; underlined)
*To whom correspondence should be addressed.
Tel: 82-42-630-9742, Fax: 82-42-636-2676
E-mail: [email protected]
primers
를사용하여pfu DNA polymerase
로PCR
반응을실시하 였다. PCR
반응은95
oC
에서3
분간열처리한후95
oC
에서30
초, 62
oC
에서40
초, 72
oC
에서3
분간반응을30
회반복하고최종적으 로72
oC
에서10
분간방치하여수행하였다.
Cellulase 정제
Cellulase
유전자를 함유한E. coli BL21(DE3)/pPES3
를ampicillin
이첨가된LB
배지에접종하여37
oC
에서진탕배양하면 서600 nm
에서 흡광도가0.7
정도에 이르렀을때IPTG
를최종농도가
0.5 mM
이되도록첨가한 후25
oC
에서5
시간진탕 배양 하였다.
배양액을원심분리하여회수한 균체를50 mM Tris-HCl
buffer (pH 8.0)
에 현탁하고 초음파로 균체를 파쇄한 후15,000
×g, 4
oC
에서20
분간원심분리함으로써균체파쇄상등액을 제조하였다.
균체 파쇄상등액에ammonium sulfate
를 첨가하여30~70% (w/v)
로 분획한 침전물을50 mM Tris-HCl buffer (pH 8.0)
에 현탁하여 동일한 완충용액으로 투석을 한 후, DEAE- Sepharose
컬럼 크로마토그래피를 수행하였다. NaCl (0~0.8 M)
로추출하여얻은활성분획을모아
ultrafiltration
으로 농축한후 이를50 mM Tris-HCl buffer (pH 8.0)
로 투석하고Q-Sepharose
컬럼크로마토그래피를수행하였다
.
결합된효소를추출하기위해서는 동일완충용액으로
NaCl (0~0.6 M)
농도구배를주어분 획하였으며효소활성을 보이는 분획들을모아서ultrafiltration
으로농축하고이를정제된효소로사용하였다
.
Cellulase 활성 측정
Cellulase
활성은CMC
를기질로 하여효소반응후에 유리된환원당을
3,5-dinitrosalicylic acid (DNS)
방법으로다음과같이정 량함으로써측정하였다.
증류수에현탁시킨1.0% (w/v) CMC
용 액0.5 ml, 200 mM sodium citrate buffer (pH 5.5) 0.25 ml
와효소 용액0.25 ml
를혼합하여55
oC
에서15
분동안 반응시켰다. DNS
시약
3 ml
를첨가하여반응을정지시키고끓는물에서5
분동안방치하여 발색시킨 후
540 nm
에서 흡광도를 측정하고 이를glucose
를표준시료로사용하여동일조건하에서발색시켜조사한흡광도와비교함으로써 유리된환원당의양을결정하였다
.
효소 활성도1.0 unit
는위의조건하에서1
분동안CMC
로부터1 µmol
의
glucose
에상응하는환원당을생성하는효소의양으로정의하였다
. p -Nitrophenyl-
β-glycosides
의분해활성을조사하기위해서는기질을
1 mM
사용하였으며10
분간반응시킨후반응액의2
배부피의
1 M Na
2CO
3 용액을첨가하여반응을종결시키고405 nm
에 서흡광도를측정하였다.
효소의활성도는1
분동안1
µmol
의p - nitrophenol
을유리시키는효소양을1 unit
로정의하였다.
반응산물 분석
Cellooligosaccharides
를반응기질로하여효소반응을 수행하고반응액을
95
oC
에서3
분동안열처리한후원심분리하여단백질침전물을 제거하고 상등액을 적정량 취해
n-propanol,
nitromethane
과증류수[7:1:2, (v/v)]
혼합용액을전개용액으로하 여silica gel-precoated thin layer plate (Merck Kiesegel, No.
5748)
에서박층크로마토그래피를수행하였다.
전개된물질을발색시키기 위해서는
9 ml ethanol, 0.5 ml p -anisaldehyde, 0.5 ml sulfuric acid
와glacial acetate
몇 방울을 혼합한발색제 용액을 뿌린후, 120
oC
에서10
분간방치하였다.
결과 및 고찰
Cellulase의 과잉발현B. licheniformis WL-12
의cellulase
유전자를대장균에서 과잉발현시키기위해
signal peptide
를포함하는cellulase
구조유전자 가 증폭되도록 설계된12CelF
와12CelR primers
를 사용하여PCR
을 수행하였다. Primers
의 서열에는pET23a(+)
에cellulase
유전자를 크로닝하기 위해
Nde I
과Xho I
을 도입하였다.
따라서 증폭한cellulase
유전자를Nde I
과Xho I
으로절단한 후구조유전자부분을추출하고이를동일한효소로절단된
pET23a(+)
에도입함으로써 재조합 플라스미드
pPES3
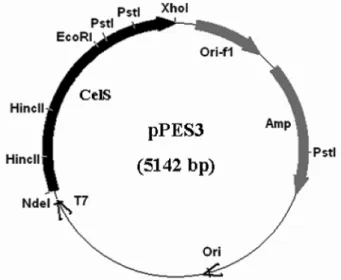
을 제조하였다(Fig. 1).
pPES3
에 삽입된cellulase
유전자의 염기서열을 결정하여PCR
반응중변이가일어나지않은것을확인하였다
.
재조합 플라스미드
pPES3
을E. coli BL21(DE3)
에 도입하여 얻은형질전환주를LB
배지에 접종하여37
oC
에서진탕배양하면서
IPTG
를처리하였다.
발현된cellulase
의수용화정도를높이 기위해IPTG
처리후에는배양온도를25
oC
로낮추어5
시간배양하였다
.
균체를회수하고초음파로파쇄한후원심분리하여얻 은 균체파쇄상등액과 배양상등액에 존재하는cellulase
활성을비교한결과배양부피기준으로균체파쇄상등액은
5.28 U/ml
의 효소활성을 보였으나,
배양상등액에서는균체 파쇄상등액의약1%
정도에해당하는효소활성을보였다.
이로보아재조합대장균에서 생산된
cellulase
는대부분이균체내에존재하는것으로Fig. 1. Structure of recombinant plasmid pPES3 containing the cellulase gene. Black bar indicates the cellulase gene. The arrows indicate the direction of transcription of genes. Abbreviations are as follows: T7, T7 RNA polymerase promoter; CelS, cellulase gene;
Amp, ampicillin resistance gene; Ori-f1, f1 origin; Ori, replication
origin of plasmid pBR322.
확인되었는데
,
대장균에도입된cellulase
유전자가signal peptide
를부분을포함하고있음에도불구하고균체외로분비되지않았 음을알수있다
.
대장균에서cellulase
또는이와유사한균체외 분비효소의외래유전자가발현될 경우일정량의효소가균체외 부로 분비되는경우가 다수보고된 바가 있다. B. licheniformis
의내열성 α
-amylase
유전자는T7 promoter
를이용하여대장균 에서과잉발현되었을때생산된효소가균체외로분비되며대 장균분비계가 α-amylase
의signal peptide
를인지할 수있는것 으로알려졌고(19), B. licheniformis
의endo-
β-1,3-1,4-glucanase
도대장균에서생상될때약
40%
이상이균체외로분리된다고보 고되었다(15).
또한pUC19
에크로닝된WL-12 cellulase
유전자로형질전환시킨대장균에서도생산된
cellulase
일부가균체외 로분비되었고(
결과 미제시), B. subtilis
형질전환주는배양상등 액에WL-12 cellulase
를7 U/ml
로생산하였다(23).
이와는 달리E. coli BL21(DE3)/pPES3
에서생산된cellulase
가분비되지않은 것은WL-12
의cellulase
특성이기보다는IPTG
를첨가한후배 양온도를25
oC
로낮추어signal peptide
가효과적으로 제거되지 못한 때문으로 여겨진다. B. subtilis
의mannanase
유전자를T7 promoter
에도입하여BL21(DE3)
에서IPTG
를첨가하여37
oC
에 서과잉발현을유도하였을때signal peptide
를포함하지않는유전자는 균체 내에 과량의
mannanse
를 생산하였으나, signal
peptide
를포함한 유전자는거의mannanase
를생산하지않은것으로확인된바있다
(11).
한편최근에B. subtilis 1N
의2
개cel- lulases
유전자를T7 promoter
에도입하여BL21(DE3)
에서 발현하였을 때생산성이 배양액 기준으로
0.52 U/ml
과0.22 U/ml
로 확인된것에 비하면(13)
대장균에서WL-12
의celllulase
생산성 은훨씬높은것을알수있다.
Cellulase의 정제
BL21(DE3)/pPES3
로부터 생산된cellulase
를SDS-PAGE
에서 활성염색을실시한결과3
종류이상의활성bands
가확인되었다(
결과미제시). WL-12
의cellulase
가한개의활성영역과한개의cellulose binding domain (CBD)
으로 구성되어 있고대장균에서생산된
cellulase
의C-
말단에 존재하는CBD
가분해되어도 활성영역은
cellulase
활성을 유지할 수 있으므로 여러 개의 활성bands
가관찰된것으로여겨진다.
이러한현상은다른균주유래의
cellulases
를대장균에서 발현하였을 때도보고된 바있고(3,
9),
이미WL-12 cellulase
유전자를B. subtilis
에 발현시켰을때 도2
개 이상의 활성bands
가 관찰된 바 있다(23).
그런데B.
licheniformis B-41361
의cellulase
를 대장균에서 과잉 발현하여불용성단백질로존재할 경우
proteolysis
를받지않는다는사실이밝혀졌다
(3).
Cellulase
를 정제하기 위해서 대장균 균체 파쇄상등액을ammonium sulfate (30%~70%)
로 분획하여 얻은 단백질을DEAE-Sepharose
컬럼 크로마토그래피로 정제하였는데resin
에결합되지 않고 그대로 용출되는
cellulase
와resin
에 결합되어NaCl
농도구배를 주었을 때용출되는cellluase
로구별되었다.
SDS-PAGE
로활성염색을하여resin
에결합되지않은효소와결합된효소를비교하였을때
resin
에결합되지않은cellulase
는크 기가 작은 것으로 확인되었다.
본 연구에서는 크기가 큰cellulase
를정제하기위해NaCl
로용출한분획중cellulase
활성 을보이는 분획을ultrafiltration
으로농축하고 이를Q-Sepharose
컬럼크로마토그래피로정제하였다
. Q-Sepharose
에결합된단백질을
NaCl
로 용출한 후활성분획을 모아 농축하여SDS-PAGE
로분석한결과
Fig. 2
에보인바와 같이분자량이약49.5 kDa
인
cellulase
가 확인되었는데 이는 대장균에서 정제된B.
licheniformis NBL420
의cellulase-chitosanase
의분자량과 동일하였다
(8).
유전자의 염기서열에서 유추되는cellulase
의 분자량은56.8 kDa
과비교해 보았을때정제된 단백질은 그크기가 약7
kDa
작은 것으로 보아signal peptide
지역이나C-
말단 지역의 일부가 절단되었을 것으로 추정된다.
대장균에서 정제된B- 41361 cellulase
는 분자량이42 kDa
으로 유전자로부터 예측되는 단백질의 분자량보다 약15 kDa
이작은 것으로보고되었다(3).
그런데
B. licheniformis B-41361
의배양상등액에서도여러개의cellulase
활성단백질bands
가관찰되었으나,
분자량이35 kDa
에해당하는단백질이
cellulase
활성이가장높은것으로보고되었다
(2).
유전자 서열에서 분자량이27.4 kDa
으로 유추된B.
licheniformis
의cellulase
나(15)
많은Bacillus
유래의cellulases
의 분자량(<42 kDa)
보다는 재조합 대장균에서 생산된WL-12
의cellulase
는분자량이큰것을 알수있다.
한편BL21(DE3)
에서 과잉발현된B. subtilis 1N(13)
의cellulase
는분자량이54 kDa
로 확인되었으며B. stearothermophilus No. 236
의배양상등액에서 정제된cellulase
는분자량이95 kDa
에이르는것으로보고되었다(10).
Fig. 2. SDS-PAGE of the cellulase purified from recombinant E. coli . Lane 1, the molecular weight markers; 2, the purified enzyme.
Molecular size is shown in kilodaltons to the left side of the gel.
Cellulase의 반응특성
대장균이생산한
WL-12
의cellulase
활성에미치는반응온도와 반응pH
의 영향을 분석하였다.
그 결과WL-12
의cellulase
는55
oC
와pH 5.5
에서 최대의 활성을 보였다(Fig. 3). B.
licheniformis B-41361
배양상등액에서 정제된 분자량35 kDa
의cellulase
와대장균에서정제된42 kDa
의cellulase
는모두65
oC
와pH 6.0
에서 최대활성을 보여WL-12
의cellulase
와는 다른 것으 로보고되었다(2, 3).
그런데Bacillus
유래의cellulases
는반응온도가
50~70
oC,
반응pH
는5.0~7.0
범위에서 최대활성을보이는 효소가다수인 것을 보면WL-12
의cellulase
는Bacillus
유래의일반적인
cellulase
특성을보이는것으로판단된다.
또한열안정성을분석하기위해온도를달리하여효소액을
3
시간방치하면 서시간별로잔존활성을측정한 결과50
oC
에서는3
시간까지안 정하였으나55
oC
이상에서는1
시간방치후에도급격히효소활 성이실활되었으며3
시간방치후에실활된 정도와유사하였다(Fig. 4). WL-12 cellulase
는 열안정성은B-41351
과B.
amyloliquefaciens DL-3
의 효소에 비해 낮은 편이며(3, 12), NBL420
이나Bacillus sp. CH43
과HR68
의효소와는유사하였다(8, 16).
최적반응조건에서CMC
를기질로 하여정제된WL-12 cellulase
를반응하였을때효소비활성은163 U/mg
으로확인되었 으며 이는 대장균에서 생산된B-41351
의cellulase
비활성(60.7 U/mg)
보다는 높고B-41351
의 배양상등액에서 정제된35 kDa cellulase
의비활성(183 U/mg)
과는유사하며,
최근에정제 된DL-3
의cellulase
비활성(6,070 U/mg)
보다는매우낮았다.
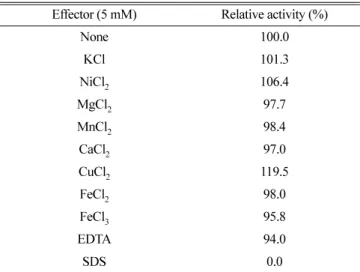
금속이온이나화합물이
cellulase
활성에미치는영향을조사하기위해반응액에
5 mM
의농도로첨가하여정제된효소로반응을실시하였다
. Table 1
에보인바와같이SDS
의존재하에서는 효소활성이거의대부분저해되었으며,
대부분의다른화합물은적은범위에서효소활성을저해또는증가시키는것으로나타났 다
. Cu
2+은B. licheniformis B-41361
의cellulase
와는 달리WL- 12
의cellulase
활성을 증가시켰고, WL-12
의cellulase
활성에거 의영향을 미치지않는Mn
2+은cellulase
종류에따라그활성에 미치는 영향이 다양한것으로 보고되었다(3, 10, 12, 16).
또한EDTA
는 대부분의Bacillus
유래의 효소와 같이WL-12
의cellulase
활성을저해하지않았지만, DL-3
의cellulase
활성을상당히 저해하였고
B. subtilis AH18
의cellulase
의 활성도저해하 였다(12, 22).
기질특이성과 반응산물
정제된 효소를 이용하여
CMC
를비롯한 여러 다당류의가수Fig. 3. Effects of reaction temperature and pH on the cellulase.
Temperature profile (
■) was obtained by measuring the cellulase activities at different temperatures and pH 5.5. The reactions was done at 55
oC and various pHs for determining the pH profile (dotted line).
Buffers used were as follows: sodium citrate (
●), sodium phosphate (
○), Tris (
▼).
Fig. 4. Thermostability of the cellulase. The residual relative activity was determined by measuring after preincubation for 3 h at various temperatures with a fixed pH 6.5.
Table 1. Effects of metal ions and other reagents on the cellulase activity
Effector (5 mM) Relative activity (%)
None 100.0
KCl 101.3
NiCl
2106.4
MgCl
297.7
MnCl
298.4
CaCl
297.0
CuCl
2119.5
FeCl
298.0
FeCl
395.8
EDTA 94.0
SDS 0.0
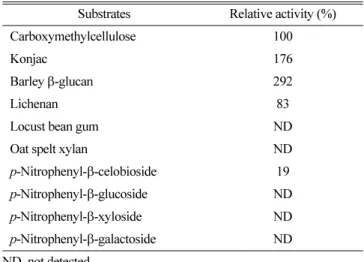
분해활성을비교하였다
.
효소반응에사용된다당류는모두반응액에
0.5%
농도로하여실험을실시하였는데,
정제된cellulase
는glucose
가 β-1,4
결합으로구성된CMC
의가수분해능보다 β-1,3- 1,4
결합으로이루어진barley
β-glucan
에대한가수분해능이2
배이상 높았으며
lichenan
에 대한 가수분해능은CMC
분해능의83%
에 이르렀다(Table 2). Lichenan
과barley
β-glucan
은 모두glucose
가β-1,3-1,4
결합에하고있지만β-1,3
결합과β-1,4
결합의비율이 차이가 있으며
lichenan
은barley
β-glucan lichenan
에 비해 β-1,3
결합의 비율이 높다. Locust bean gum
이나oat spelt xylan
은cellulase
에의해가수분해가일어나지않은것으로보아WL-12
의cellulase
는mannanase
와xylanase
활성은 없는것으로 확인되었다. Konjac
은glucomannan
다당류로 구성되어 있는데cellulase
의경우glucose
잔기간의결합을분해하고mannanase
의경우는
mannose
잔기간의결합을 분해할수있다.
따라서WL-
12 cellulase
가locust bean gum
을 분해하지 못하면서konjac
의분해능을보이는것은
cellulase
활성에의한것임을알수있다.
WL-12
의cellulase
는CMC
보다 β-glucan
의 분해능이 높은Bacillus sp. CH43
과HR68
의cellulases
와유사한 기질특이성을 보였다(16).
그러나B-41351
과DL-3
의cellulase
도β-glucan
을분 해하지만CMC
보다는 분해활성이 낮았으며(3, 12), DL-3
와B.
stearothermophilus No. 236
은WL-12
의cellulase
와는달리xylan
을 분해하는 것으로 보고되었다
(22).
한편p -nitrophenyl-
β- glycosides
를기질로하여분해력을조사한 결과p -nitrophenyl-
β- cellobioside
는분해하였지만 β-xylosidase
와 β-galactosidase
활성 뿐 아니라 β-glucosidase
의 활성은 없었다.
실제 많은cellulases
가
p -nitrophenyl-
β-cellobioside
은 분해하지만p -nitrophenyl-
β- glucoside
는분해하지못하는것으로알려졌다(3, 9, 12, 22).
Cellulase
에 의한 가수분해 산물을 분석하기 위해 기질로glucose
의 중합도가 다른cellobiose, cellotriose, cellotetraose
그리고
cellopentaose
를사용하여과량의효소를 처리한후가수분해산물을
TLC
로조사하였다.
그결과cellobiose
는전혀분해되 지않았으나중합도가3
개이상인올리고당은모두분해되는것 으로 나타났으며cellotriose
의분해정도가 가장 낮았다(Fig. 5).
Cellopentaose, cellotetraose
와cellotriose
의최종 가수분해산물로는