바실러스 프로바이오틱스를 이용한 발효대두의 제조 및 생리활성
박홍진1․오현화1․정도연2․김영수1
1전북대학교 식품공학과
2(재)발효미생물산업진흥원
Physiological Properties of Fermented-Soybean Manufactured with Selected Bacillus Probiotics
Hong-Jin Park1, Hyeon Hwa Oh1, Do Youn Jeong2, and Young-Soo Kim1
1Department of Food Science and Technology, Chonbuk National University
2Microbial Institute for Fermentation Industry (MIFI)
ABSTRACT Bacillus subtilis strains that have probiotic properties, such as acid resistance, bile resistance, intestinal adhesion, and antimicrobial activity, were isolated from Korean traditional fermented soybean products. The in vitro anti-obesity and anti-cancer effects of extracts from fermented-soybeans produced with the selected strain (B. subtilis JBS17166) were investigated. B. subtilis JBS17166 had a high survival rate of 70.52% in acidic environments and 80.22% in bile environments. This strain had high adhesion rates for Caco-2 cells and mucin with 78.59% and 74.32%, respectively. The antimicrobial activity of this strain on Bacillus cereus KACC 13064 and Staphylococcus aureus KACC 1927 was also confirmed. The extract from fermented-soybean, which was developed using B. subtilis JBS17166, had a high anti-obesity effect (59.95%) on 3T3-L1 preadipocytes at 24 h. In addition, this extract had anti-cancer activity (IC50), showing 1.80 mg/mL and 1.09 mg/mL for HepG2 and HT-29 cells, respectively. Based on these results, it is expected that fermented-soybeans manufactured using B. subtilis JBS17166 as a probiotic could be beneficial as a synbiotic with anti-obesity and anti-cancer effects.
Key words: Bacillus probiotics, fermented-soybean, anti-obesity, anticancer, synbiotic
Received 21 October 2019; Accepted 16 November 2019 Corresponding author: Young-Soo Kim, Department of Food Sci- ence and Technology, Chonbuk National University, Jeonju, Jeon- buk 54896, Korea
E-mail: [email protected], Phone: +82-63-270-2569
Author information: Hong-Jin Park (Graduate student), Hyeon Hwa Oh (Researcher), Young-Soo Kim (Professor)
서 론
인간의 위장관은 미생물의 복잡한 생태계에 의해 숙주인 인간과 미생물총(microbiota)의 공생관계가 이루어진다. 장 내미생물 문(phylum)은 주로 4개의 문(Bacteroidetes, Fir- micutes, Actinobacteria, Proteobacteria)이 있으며, 이 중에서 Bacteroidetes와 Firmicutes가 장내세균의 70~90
% 이상을 차지한다(Eckburg 등, 2005). 정상 성인의 경우 장내세균 중 Firmicutes(Bacillus, Clostridium, Lactoba- cillus, Streptococcus, Staphylococcus 등)가 65~70%를 차지하는 반면, Bacteroidetes(Bacteroides thetaiotaomi- cron, Bacteroides acidifaciens, B. distasonis, Prevote- lla 등)는 15~20%를 유지하는데, Firmicutes의 비율이 증 가할수록 비만을 촉진하는 것으로 보고되었다(Tennyson과
Friedman, 2008). 이와 같은 기작은 숙주에 의해 소화되지 않은 다당류가 Firmicutes에 속하는 미생물에 의해 발효되 어 단당류나 단쇄지방산으로 분해되어 더 많은 칼로리를 체 내로 흡수시킬 수 있기 때문으로 추정된다(Turnbaugh 등, 2006). 이에 비해 유익한 장내세균은 인간에게 비타민과 단 쇄지방산을 생산하여 다양한 영양분을 제공하고 병원성 미 생물의 감염을 억제하며, 정상 면역반응을 조절하는 등의 중 요한 기능을 하는 것으로 보고되었다(Markowiak과 Sliz- ewska, 2017).
한편 장내 유해균(병원균)을 제어하기 위한 방법으로 항 생제와 항균제를 투여하면 장내 유익균의 생육에도 영향을 주기 때문에 숙주에 유익한 영향을 주는 살아있는 미생물인 프로바이오틱스가 항생제 사용의 대안으로 제시되고 있다 (Maftei, 2019). 생균제인 프로바이오틱스는 위장관 환경에 서 생존 및 증식이 가능해야 하므로 내산성, 내담즙성, 장부 착능과 잠재적 병원균에 대한 항균 활성이 우수해야 한다 (FAO/WHO, 2002). 프로바이오틱스로는 Bifidobacterium, Lactobacillus, Enterococcus, Bacillus 속 균주가 있으나 대부분의 연구가 유산균에 집중되어 있다(Yoon 등, 2013).
Bacillus 속 균주는 내성포자를 형성하기 때문에 열과 pH 변화에 대해 높은 생존율을 유지하는 것으로 보고되었다
(Barbosa 등, 2005). Bacillus 속 균주 중 Bacillus subtilis 는 Firmicutes 문에 속해있는 GRAS(Generally Recognized as Safe Substance) 균주로 안전성이 확보된 식품원료이며 프로바이오틱스로서 유용성에 대한 관심이 증대되고 있다 (Cui 등, 2013).
프로바이오틱스가 장내에서 서식하기 위해서는 적절한 영양분의 공급이 필요하다. 이들 영양분인 프리바이오틱스 는 대장에서 몇몇 세균의 성장과 활동을 선택적으로 자극하 여 건강을 개선함으로써 숙주에게 유익을 줄 수 있는 비소화 성 식품성분으로 정의된다(Gibson과 Roberfroid, 1995). 주 요 프리바이오틱스로 inulin, oligofructose, galacto-oli- gosaccharide, galactofructose 등이 있으며, 이들은 미생 물 성장과 활동의 기질로 작용하여 숙주의 건강에 도움을 준다(Yoon과 Shin, 2017). 특히 대두(soybean seed)는 난 소화성 galacto-oligosaccharide가 풍부하여 프리바이오 틱스로써 사용 가능한 유용한 식품원료이다(Gourineni 등, 2011).
최근에는 프로바이오틱스와 프리바이오틱스를 함께 식이 하면 각각의 생리활성이 동시에 나타나게 되어 시너지가 발 생한다는 장점이 있어(Kim, 2016) 신바이오틱스에 대한 새 로운 영역이 확장되고 있다. 신바이오틱스는 장 건강(Choura- qui 등, 2008; O’Bryan 등, 2013), 면역(Kanamori 등, 2006), 항암 효과(Van Loo 등, 2005)와 같은 기능성을 갖는다고 알려져 있다. 따라서 본 연구에서는 전통발효식품으로부터 분리된 Bacillus 균주 중 B. subtilis 6종에 대하여 이들의 프로바이오틱스 특성(내산성, 내담즙성, 장부착능 및 항균 활성)을 비교한 후 가장 우수한 균주를 선발하였다. 또한 선발된 균주로 제조한 대두발효물의 기능성(항비만, 항암)을 평가하여 신바이오틱스로서의 가능성을 확인하였다.
재료 및 방법
실험 재료 및 기기
실험 균주는 선행 연구 결과 분리 선발된 B. subtilis로 동 정된 6개의 균주(JBD17091, JBS17166, JBG18023, JBC 18075, JBC18087, JBS18101)와 비교 균주인 B. subtilis KACC 10854(Korea Agricultural Culture Collection (KACC), Wanju, Korea)를 사용하였다.
Nutrient agar(NA)와 nutrient broth(NB)는 Becton, Dickinson & Co.(Franklin Lakes, NJ, USA) 제품을 사용 하였고 bile bovine, mucin, TritonTM X-100, sodium phosphate monobasic, sodium phosphate dibasic, Oil Red O solution, 3-isobutyl-1-methylxanthine(IBMX), insulin from bovine pancreas, dexamethasone, rosigli- tazone, epigallocatechin gallate(EGCG), formaldehyde solution, 2-propanol은 Sigma-Aldrich Co.(St. Louis, MO, USA) 제품을 사용하였다. Dulbecco’s modified Eagle’s medium(DMEM), bovine calf serum(BCS), fetal bovine
serum(FBS), antibiotics(penicillin/streptomycin), non- essential amino acid(NEAA), 4-(2-hydroxyethyl)-1- piperazineethanesulfonic acid(HEPES), 0.5% trypsin- EDTA, Dulbecco’s phosphate-buffered saline(DPBS)은 Gibco BRL(Grand Island, NY, USA) 제품을 사용하였으 며, cell counting kit-8(CCK-8)은 Dojindo Molecular Technologies(Rockville, MD, USA) 제품을 사용하였다.
식품유해균(Bacillus cereus KACC 13064, Listeria mon- ocytogenes KACC 10764, Staphylococcus aureus KACC 1927, Escherichia coli KACC 10115)은 한국농업미생물 자원센터(Korea Agricultural Culture Collection(KACC)) 에서 분양받아 사용하였다. 미생물의 배양 및 보관용으로 원심분리기(CT15RE, Hitachi Koki Co., Ltd., Hitachinaka, Japan), 정치배양기(HB-101M, Hanbaek Scientific Co., Ltd., Bucheon, Korea), 진탕배양기(SI-2S, Universal Scientific Industrial Co., Ltd., Shanghai, China) 및 초저 온 냉동기(DF8517, Ilshin Lab Co., Ltd., Dongducheon, Korea)를 사용하였다. 세포 배양을 위하여 CO2 incubator (MCO-18AC, Panasonic Healthcare Co., Ltd., Osaka, Japan)와 도립 삼안 위상차 현미경(TCM400, Labo Amer- ica Inc., Fret, CA, USA)을 사용하였다.
균주 배양
실험 균주는 NA에 활성화시켜 NB에 단일콜로니를 접종 한 후 37°C에서 180 rpm으로 18시간 동안 배양하였다. 균 주 배양액은 원심분리(4°C, 10,000×g, 5분) 한 다음, 상층 액과 pellet을 분리하여 사용하였다. 상층액은 항균 활성 평 가에 사용하였고, pellet은 내산성, 내담즙성 및 장부착능 평가에 사용하였다. Pellet은 내산성, 내담즙성 평가에 NB 를 첨가하여 재현탁시켰고, 장부착능 평가를 위하여 DPBS 를 이용하였다. 현탁액에 NB(또는 DPBS)를 첨가하여 현탁 액의 농도를 조정하였고, OD(Abs. 600 nm)를 측정하여 생 균수와 비교했을 때 OD=0.5가 생균수 7~8 log CFU/mL임 을 확인하였다.
내산성 및 내담즙성
내산성과 내담즙성 측정은 Seo 등(2011)의 방법을 일부 변형하여 실시하였다. 즉 내산성 측정을 위해 pH 2.5로 조정 한 NB에 균주 배양액(7~8 log CFU/mL)을 10% 접종한 후 37°C에서 180 rpm으로 2시간 배양하였다. 배양 후 0.1 M sodium phosphate buffer(pH 6.2)로 연속 희석한 후에 NA에 도말, 배양하여 생균수 변화를 측정하였다. 내담즙성 측정을 위하여 bile bovine이 0.3%(w/v) 포함된 NB에 균주 배양액(7~8 log CFU/mL)을 접종하고 37°C에서 180 rpm 으로 24시간 배양하였다. 배양 후 0.1 M sodium phosphate buffer(pH 6.2)로 연속 희석하고 NA에 도말, 배양하여 생균 수 변화를 측정하였다. 각 실험에서 처리 전 생균수에 대한 처리 후 생균수의 백분율을 생존율로 평가하여 비교하였다.
Caco-2 cell을 이용한 장부착능
Human의 colon에서 유래한 Caco-2 cell의 배양을 위하 여 DMEM에 10% FBS(v/v), 1% NEAA(v/v)와 1% anti- biotics(penicillin/streptomycin)(v/v)가 함유된 배지를 사 용하였다. CO2 배양기에서 37°C와 5% CO2를 유지하여 실 험을 진행하였으며, 현미경으로 배양상태를 관찰하면서 단 층을 형성하였을 때 0.05% trypsin-EDTA를 처리하여 계 대배양 후 본 실험에 사용하였다.
Caco-2 cell을 이용한 장부착능 측정은 Azcarate-Peril 등(2009)의 방법을 일부 변형하여 실시하였다. 배양 중인 Caco-2 cell을 12-well plate(Corning Inc., Corning, NY, USA)에 6.5×104 cells/mL로 seeding 한 후, 2일 간격으로 배지를 교체하며 21일간 배양하였다. 배양 후 DPBS를 이용 하여 세척한 다음, 아무것도 첨가하지 않은 DMEM 배지를 각 well에 주입하여 1시간 배양하였다. 그리고 균주 현탁액 1 mL를 주입하고 37°C에서 1시간 동안 배양하여 Caco-2 cell에 균주를 부착시켰다. 그 후 DPBS로 세척하여 cell에 부착되어있지 않은 균주를 제거하고 부착된 균주는 0.05%
triton X-100을 이용하여 탈착시킨 뒤 회수하였다. 회수된 액을 PBS로 연속 희석한 후 NA에 도말하여 37°C에서 18시 간 배양한 후 생균수를 측정하였다. 장부착능은 처리 전 측 정된 생균수(log CFU/mL)에 대한 처리 후 회수된 생균수 (log CFU/mL)의 백분율로 나타내었다.
Mucin을 이용한 장부착능
Mucin을 이용한 장부착능은 Bengoa 등(2018)의 방법을 일부 변형하여 측정하였다. Mucin을 DPBS에 3 mg/mL 농 도로 제조한 다음, 100 μL씩 96-well plate(Corning Inc.) 에 주입하여 4°C에서 overnight 하였다. 그리고 DPBS를 이용하여 두 차례 세척하여 plate를 준비하였다. 각 well에 균주 현탁액(6~8 log CFU/mL) 100 μL를 주입하고 37°C 에서 1시간 동안 incubation 하여 mucin에 균주를 부착시켰 다. 그 후 DPBS로 세척하여 mucin에 부착되어 있지 않은 균주를 제거하고 부착된 균주는 0.05% triton X-100을 처 리하여 탈착시킨 후 회수하였다. 회수된 액은 PBS로 연속 희석한 후 NA에 도말하여 배양(37°C, 18시간)한 다음 생균 수를 측정하였다. 장부착능은 처리 전 측정된 생균수에 대한 처리 후 회수된 생균수의 백분율로 나타내었다.
항균 활성
항균 활성은 paper disc diffusion assay를 사용하여 측 정하였다. 즉 식품 유해균(B. cereus KACC 13064, L. mono- cytogenes KACC 10764, S. aureus KACC 1927, E. coli KACC 10115)을 NB에 접종하여 진탕배양(37°C, 180 rpm, 18시간)한 후, 멸균된 면봉을 사용하여 NA에 도말하고 6 mm paper disc를 부착시켜 plate를 준비하였다. 그 위에 균주배양 상층액을 20 μL 분주하여 37°C에서 18시간 배양 하였다. 배양 후에 paper disc 주변으로 유해균이 자라지
못해 투명하게 보이는 환의 지름을 측정하여 균 집락의 지름 에 대한 투명환의 지름을 상대활성도로 나타내었다.
대두고형발효 및 추출물 제조
대원콩(soybean seed, Gunsan, Korea)을 ddH2O에 overnight 침지한 후 물기를 제거하여 30 g씩 플라스크에 담아 121°C에서 30분간 증자하였다. 증자 후 상온으로 식 힌 콩에 균주 배양액을 1%(v/w)로 접종하고 37°C에서 24 시간 및 48시간 동안 발효시켰다. 대조구는 균주가 접종되 지 않은 시료를 control로 하였고, 선발균주(Bacillussub- tilis JBS17166)와 비교균주(Bacillussubtilis KACC 10854) 를 접종하여 발효하였다. Control과 발효물은 동결건조 하여 분말화한 다음 70% 에탄올로 상온에서 2시간 교반 추출하 였으며, 감압농축 하여 추출물의 에탄올을 제거하고 동결건 조 하여 기능성 평가(항비만, 항암)의 시료로 사용하였다. 동 결건조 분말로부터 추출된 대두발효물의 추출수율은 11.78
±0.23%로 추출수율에 유의차가 보이지 않아 기능성 평가 에 동일농도로 처리하여 분석하였다.
항비만 활성(in vitro)
3T3-L1 preadipocyte cell의 세포생존율은 10% BCS 와 1% antibiotics(penicillin/streptomycin)가 첨가된 DMEM 배지에 배양시킨 후 5×104 cells/mL로 96-well plate에 seeding 하여 37°C, 5% CO2 배양기에서 24시간 배양하였 다. 각 시료는 세포에 처리했을 때 최종농도가 0.25, 0.5, 0.75, 1 mg/mL가 되도록 처리한 다음 48시간 배양하였다.
배지를 제거한 후 PBS 100 μL와 CCK-8 시약 10 μL를 첨가하여 암실에서 2시간 반응시킨 다음 450 nm에서 흡광 도를 측정하여 세포생존율을 계산하였다.
Cell viability (%)= Absorbancesample ×100 Absorbancecontrol
항비만 활성 확인은 mouse 유래 3T3-L1 preadipocyte cell의 지방분화 시 생성되는 지방축적 억제능을 비교한 Zebisch 등(2012)의 방법을 이용하였다. 10% BCS와 1%
antibiotics(penicillin/streptomycin)가 첨가된 DMEM 배 지에 3T3-L1 지방전구세포를 배양시킨 후 회수하였다.
24-well plate의 각 well에 세포 현탁액을 2×105 cells/mL 씩 seeding 한 후에 37°C, 5% CO2 조건에서 confluency가 될 때까지 배양하였다. 세포에 분화유도제 MDI(0.5 mM IBMX, 1 μg/mL insulin, 1 μM dexamethasone, 2 μM ro- siglitazone)와 10% FBS가 첨가된 DMEM 배지를 처리하 여 2일 동안 분화를 유도하였고, 2일 후 DMEM(10% FBS, 10 μg/mL insulin) 배지로 교체하여 2일간 배양하였다. 시 료는 세포독성 평가 결과 80% 이상 생존율을 유지한 최대농 도의 시료를 배지에 첨가하여 2일 간격으로 교체하여 배양 하였으며, 최종분화는 유도 8일 후에 확인하였다. 양성대조 군으로는 EGCG(0.1 mM)를 이용하였다.
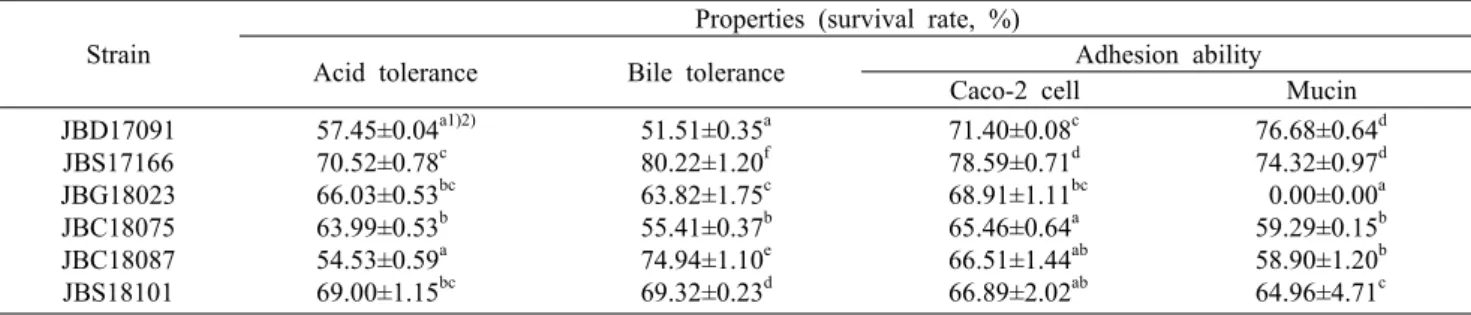
Table 1. Acid tolerance, bile tolerance, and adhesion ability of selected Bacillus subtilis strains Strain
Properties (survival rate, %)
Acid tolerance Bile tolerance Adhesion ability Caco-2 cell Mucin JBD17091
JBS17166 JBG18023 JBC18075 JBC18087 JBS18101
57.45±0.04a1)2) 70.52±0.78c 66.03±0.53bc 63.99±0.53b 54.53±0.59a 69.00±1.15bc
51.51±0.35a 80.22±1.20f 63.82±1.75c 55.41±0.37b 74.94±1.10e 69.32±0.23d
71.40±0.08c 78.59±0.71d 68.91±1.11bc 65.46±0.64a 66.51±1.44ab 66.89±2.02ab
76.68±0.64d 74.32±0.97d 0.00±0.00a 59.29±0.15b 58.90±1.20b 64.96±4.71c
1)Mean±SD.
2)Means with the different letters (a-f) within a column are significantly different according to Duncan’s multiple rage test (P<0.05).
Oil Red O의 염색방법은 Hsu와 Yen(2007)의 방법을 변 형하여 진행하였다. 세척(DPBS) → 고정(4% para form al- dehyde, 30분) → 세척(60% isopropanol) 순으로 진행하 여 plate를 완전히 건조시킨 후 60% Oil Red O 염색액을 처리하여 실온에서 20분간 염색하였다. 염색약을 제거한 후 증류수로 여러 번 세척하여 현미경(TCM400, Labo Amer- ica, Inc., Fremont, OH, USA)으로 관찰한 다음 camera (EyeCam, Bimeince Co., Suwon, Korea)로 기록하였다.
100% isopropanol 첨가 후 30분 동안 반응시켜 염색된 지방 구를 용출시킨 다음 microplate reader(US/Eon, BioTek Instruments, Inc., Winooski, VT, USA)를 사용하여 500 nm에서 흡광도를 측정하였다. 항비만 활성은 control(dif- ferential media, DM)의 흡광도에 대한 각 시료첨가구의 흡 광도 백분율을 지방축적률로 환산하여 평가하였다.
항암 활성
항암 활성은 human으로부터 유래한 간암 세포인 HepG2 cell과 결장암 세포인 HT-29 cell을 사용하여 평가하였다.
HepG2 cell의 배양은 DMEM에 10%(v/v) FBS, 2.5%
HEPES, 1% NEAA, 1%(v/v) antibiotics(penicillin/strep- tomycin)가 함유된 배지를 사용하였고, HT-29 cell의 배양 은 DMEM에 10%(v/v) FBS, 1%(v/v) antibiotics(peni- cillin/streptomycin)가 함유된 배지를 사용하였다. CO2 배 양기에서 37°C, 5% CO2를 유지하여 실험을 진행하였으며, 현미경을 통해 배양상태를 관찰하면서 단층을 형성하였을 때 0.05% trypsin-EDTA를 처리하여 계대배양 후 본 실험 에 사용하였다.
항암 평가를 위하여 배양 중인 각 세포를 5×104 cells/
mL로 96-well plate에 seeding 하여 24시간 배양하였다.
각 시료는 세포에 처리했을 때 최종농도가 0.25, 0.5, 1.0, 2.0, 3.0, 4.0, 5.0 mg/mL가 되도록 연속 희석하여 준비하였 고, 희석된 시료를 각 well에 주입한 후 48시간 배양하였다.
배양 후 배지를 제거하고 DPBS 100 μL와 CCK-8 시약 10 μL를 첨가하여 암실, 37°C, 5% CO2 배양기에서 2시간 동안 반응시켰다. 반응 후 microplate reader를 이용하여 450 nm에서 흡광도를 측정하였고, 다음의 식을 이용하여 세포 생존율을 계산하였다.
Cell viability (%)= Absorbancesample ×100 Absorbancecontrol
각 시료의 농도별 세포생존율을 이용하여 세포 생장을 50% 저해하는 시료의 농도(IC50)를 계산하였다.
통계분석
실험 결과는 평균과 표준편차로 나타내었다. 각 시료 간 의 유의성 검사는 SPSS package program(Ver.12.0K, SPSS Inc., Chicago, IL, USA)을 사용하여 실시하였으며, P<0.05 수준에서 one-way ANOVA로 분산분석 한 다음 Duncan’s multiple range test로 비교하였다.
결과 및 고찰
내산성 및 내담즙성
체내에 유입된 균주가 프로바이오틱스로 작용하기 위해 서는 위산과 담즙 등의 체내 소화액의 극한 환경에서 생존하 여 대장에 도달해야 한다(Lim 등, 2016). 따라서 본 연구에 서는 pH 2.5로 조정된 NB와 0.3% 담즙산이 포함된 NB에 균주를 각각 2시간, 24시간 반응시켜 생존율을 확인하였고, 그 결과를 비교하여 내산성 및 내담즙성을 평가하였다. 내산 성 평가 결과(Table 1), JBS17166 균주가 70.52%로 가장 높은 생존율을 나타내었다. 흔히 프로바이오틱스로 사용되 는 Lactobacillus 속의 생존율은 pH 2.0, 2시간 조건 하에서 35% 이상이라고 보고된 바 있다(Ding과 Shah, 2007). 이는 본 연구에서 확인된 실험균주의 생존율(54.53~70.52%)보 다도 크게 낮은 값으로, Bacillus 속이 내성포자를 보유하기 때문에 산성 조건에서 생존 가능성이 Lactobacillus 속보다 더 유리하다고 판단된다.
내담즙성 평가 결과(Table 1)에서도 JBS17166 균주가 80.22%로 가장 높은 생존율을 나타내었다. 담즙에 대한 내 성은 소장에 도달한 프로바이오틱스의 생존 및 기능성에 중 요한 역할을 하기 때문에, 장내 담즙의 농도(0.3%)에서 생존 율을 평가한다(Seo와 Lee, 2007). 본 연구에 사용된 Bacil- lus 균주는 0.3% oxgall 농도에서 51.51~80.22%로 넓은 범 위의 생존율을 보였으며, 따라서 내담즙성을 보유한 Bacil-
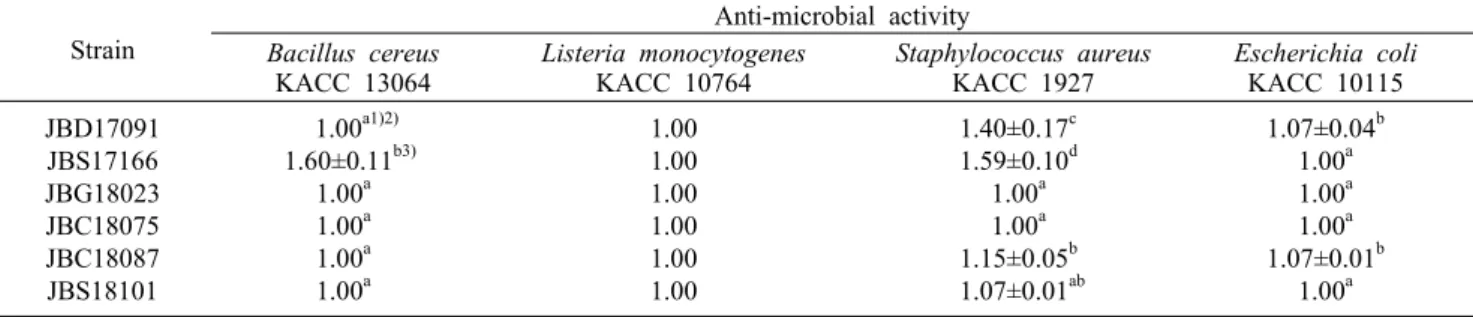
Table 2. Anti-microbial activity of selected Bacillus subtilis strains Strain
Anti-microbial activity Bacillus cereus
KACC 13064 Listeria monocytogenes
KACC 10764 Staphylococcus aureus
KACC 1927 Escherichia coli KACC 10115 JBD17091
JBS17166 JBG18023 JBC18075 JBC18087 JBS18101
1.00a1)2) 1.60±0.11b3)
1.00a 1.00a 1.00a 1.00a
1.00 1.00 1.00 1.00 1.00 1.00
1.40±0.17c 1.59±0.10d
1.00a 1.00a 1.15±0.05b 1.07±0.01ab
1.07±0.04b 1.00a 1.00a 1.00a 1.07±0.01b
1.00a The experiments were carried out in triplicate.
1)1.00 means no activity.
2)Means with the different letters (a-d) within a column are significantly different according to Duncan’s multiple rage test (P<0.05).
3)Mean±SD.
lus 프로바이오틱스로 기대된다.
Caco-2 cell 및 mucin을 이용한 부착능
대장에 도달한 균주가 체외로 배출되지 않고 프로바이오 틱스로 작용하기 위해서는 대장에서 부착된 상태를 유지해 야 한다. 더불어 인간의 대장 내에는 점막층(mucin)이 상피 세포를 덮고 있기 때문에 프로바이오틱스가 대장에 상재할 때는 상피세포뿐만 아니라 점막층 물질에도 결합하는 것으 로 알려져 있다(Kim 등, 2004). 따라서 대장 상피세포(Caco- 2 cell)를 이용한 부착능과 더불어 mucin에 대한 부착능을 진행하였으며, 그 결과를 Table 1에 나타내었다. 실험균주 의 Caco-2 cell에 대한 부착능은 65.46~78.59%를 나타내 었으며, 그중 JBD17091 균주와 JBS17166 균주가 각각 71.40%와 78.59%로 다른 균주들에 비해 높게 나타났다.
Mucin에 대한 부착능의 경우 실험균주는 0.00~76.68%를 나타내었으며, 그중 JBD17091과 JBS17166 균주가 각각 76.68%와 74.32%로 다른 균주들에 비해 크게 높았다.
Hong 등(2005)은 Bacillus 균이 분비하는 포자는 견고한 생물입자이기 때문에 위산 및 담즙산에서 생존하고 내장을 통과하여 결국에는 배출될 것으로 언급하였다. 하지만 본 연구에서는 위산과 담즙산에서 포자상태로 생존한 균주가 대장에 도달했을 때 재생할 것으로 예상하여 포자가 아닌 균주 자체의 부착 가능성을 Caco-2 cell과 mucin에 대한 부착능 평가로 확인하였다. 또한 Bacillus 종이 장내미생물 의 균형을 유지시키는 특성이 있어(Adami와 Cavazzoni, 1999) 대장에 점착 시 다른 유익미생물과 공생하여 장 건강 을 유지시킬 수 있을 것으로 기대된다. 따라서 본 연구에서 사용된 균주 중 JBS17166 균주는 Caco-2 cell과 mucin에 대한 부착능이 다른 균주보다 우수하였고, 내산성과 내담즙 성을 보유하였기 때문에 위장관을 통과하여 대장에 도달했 을 때 부착하여 생육이 가능하리라 판단된다.
항균 활성
식품 유해미생물인 B. cereus KACC 13064, L. mono- cytogenes KACC 10764, S. aureus KACC 1927 및 E.
coli KACC 10115에 대한 실험균주의 항균 활성을 측정한 결과(Table 2), JBS17166 균주는 B. cereus KACC 13064 및 S. aureus KACC 1927의 Gram positive 유해균에 대한 생육 저해능이 가장 우수하였으나, E. coli에 대한 저해활성 은 없는 것으로 확인되었다. 이는 Park 등(2019)이 보고한 JBC17126 균주의 B. cereus KACC 13064 및 S. aureus KACC 1927에 대한 항균 활성보다도 더 우수하였다. 이들 유해균에 대한 B. subtilis JBS17166 균주의 항균력은 다른 균주들에 비해 더 많은 항균물질(bacteriocin과 subtilin 등) 을 생산하기(Hong 등, 2005) 때문으로 예상된다. 또한 B. subtilis가 생산하는 lipopeptide 이외에 fengycin, iturin, bacillomycin 등 surfactin의 항균작용이 다른 여러 경쟁미 생물의 생육을 저해하고, B. subtilis의 생존력을 유지하는 데 유리하기 때문으로 판단된다(Stein, 2005). 따라서 JBS 17166 균주의 B. cereus 및 S. aureus에 대한 항균 활성은 장내 유해균의 증식을 억제하고 유익균의 증가를 유도할 수 있을 것으로 기대된다.
항비만 활성
미생물을 이용한 대두의 발효는 isoflavonoid aglycone 의 증가 및 small peptide의 생성에 의해 항비만과 항당뇨 등의 생리활성을 나타내는 것으로 알려졌다(Murphy 등, 1999). 또한 발효대두 추출물의 식이가 mouse의 지방 축적 률을 감소시킨다는 연구 결과가 보고되었다(Nam 등, 2012).
이와 같은 연구내용을 기반으로 프로바이오틱스로 선발된 B. subtilis JBS17166 균주를 사용하여 제조한 발효대두 추출물의 항비만 효과를 분석하고자 하였다.
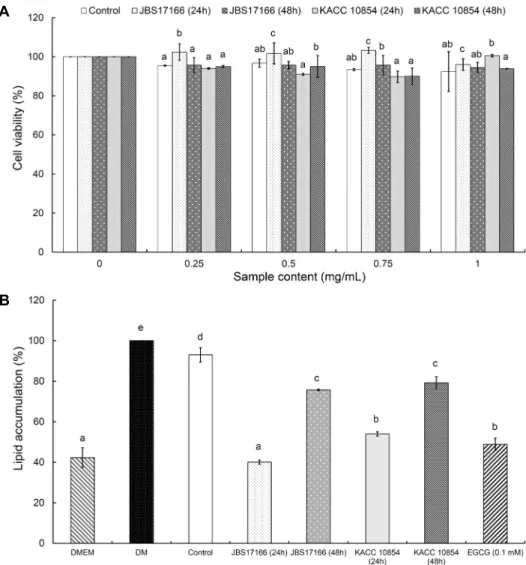
발효대두 추출물 처리농도에 따른 3T3-L1 preadipocyte cell의 세포생존율을 평가한 결과는 Fig. 1A와 같다. 추출물 의 농도는 0.25, 0.5, 0.75, 1 mg/mL로 세포배양 배지를 이용하여 희석한 후 각각 처리하였고, CCK-8 kit을 이용하 여 세포독성을 평가한 결과 1 mg/mL 농도까지 80% 이상의 생존율을 유지하는 것으로 확인되었다. 그러나 1 mg/mL 이상의 처리농도에서는 추출물이 용해되지 않고 현탁액 상 태가 되었기 때문에 시료의 최고 처리농도를 1 mg/mL로
A
B
Fig. 1. Anti-obesity activity of fermented soybean extracts on lipid accumulation in 3T3-L1 preadipocyte cell line. (A) Cell viability, (B) Lipid accumulation. Values represent mean±SD. Means with the different letters (a-e) above the bars are significantly different according to Duncan’s multiple range test (P<0.05). DMEM is undifferential media. DM is differential media. Control is non-fermented soy bean extract. EGCG is positive control and was analyzed at a concentration of 0.1 mM.
하여 지방축적률 억제활성을 비교하였다.
3T3-L1 preadipocyte cell line에 분화 유도제인 MDI와 각 추출물 시료를 1 mg/mL 농도로 처리한 시료구와 MDI만 처리한 DM의 지방축적률을 비교하였다(Fig. 1B). 즉 DM의 축적률을 100%로 하였을 때 비발효 추출물인 control의 축 적률은 93.03%였다. 한편 표준균주 KACC 10854(24 h)는 53.99%로 control보다는 축적률이 낮아졌으나 가장 낮은 축적률(40.05%)을 보인 JBS17166 균주(24 h)보다는 높은 유의적인 차이(P<0.05)를 보였다. 발효시간에 따른 지방 축 적률은 두 균주에서 모두 24시간 발효물이 48시간보다 더 낮아 발효시간이 경과함에 따라 항비만 효과가 감소하는 경 향을 보였다. 특히 비발효 추출물보다 발효 추출물이 유의적 으로 지방분화 억제효과를 보였으나 발효시간이 증가했을 때 항비만 효과는 오히려 감소하는 것으로 나타났다. Hwang 등(2015)은 3T3-L1 세포를 사용한 지방분화 실험에서 발 효된 콩의 지방축적 억제효과는 adipogenesis-related 단
백질 발현과 glucose uptake 정도에 따라 차이를 보이는 것으로 보고하였다. 따라서 본 연구에서 발효시간과 균주에 따라 지방 축적량에 미치는 영향이 다른 이유는 미생물(JBS 17166 균주 vs KACC 10854 균주) 및 발효시간(24 h vs 48 h)에 따른 지방분화 유전자의 발현량이 영향을 미쳤을 것으로 추정된다.
항암 활성
발효대두 추출물의 항암 활성 평가는 시료가 암세포의 생 존율에 미치는 영향을 확인한 후 IC50 값을 구하여 비교하였 다(Table 3). 비발효구인 control의 경우 간암세포인 HepG2 cell line에 대한 IC50 값이 3.63 mg/mL였으며, 발효 추출물 인 JBS17166 균주(24 h)와 KACC 10854 균주(24 h)는 각각 1.80과 1.59 mg/mL였다. 대장암 세포인 HT-29 cell line에 대한 IC50 값은 control이 2.45 mg/mL였으며, JBS 17166 균주(24 h)와 KACC 10854 균주(24 h)는 각각 1.09
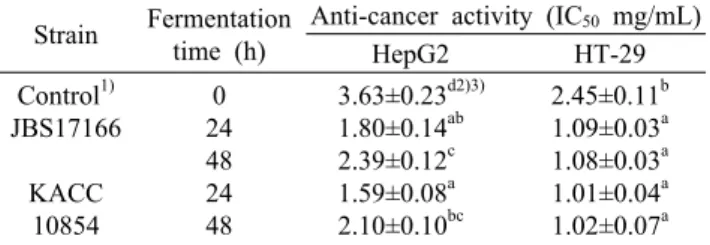
Table 3. Comparison of cytotoxicity of soybean extracts fer- mented with B. subtilis JBS17166 and B. subtilis KACC 10854 on tumor cells using CCK-8
Strain Fermentation time (h)
Anti-cancer activity (IC50 mg/mL)
HepG2 HT-29
Control1) JBS17166
KACC 10854
0 24 48 24 48
3.63±0.23d2)3) 1.80±0.14ab 2.39±0.12c 1.59±0.08a 2.10±0.10bc
2.45±0.11b 1.09±0.03a 1.08±0.03a 1.01±0.04a 1.02±0.07a The experiments were carried out in triplicate.
1)Control is non-fermented soybean extract.
2)Mean±SD.
3)Means with the different letters (a-c) within a column are sig- nificantly different according to Duncan’s multiple rage test (P<0.05).
mg/mL와 1.01 mg/mL로 확인되었다.
발효시간에 따른 발효대두 추출물의 항암 효과는 HepG2 세포에서는 유의적으로 감소하는 경향을 보였으나 HT-29 세포는 유의적인 차이를 보이지 않았다(P<0.05). JBS17166 균주와 KACC 10854 균주 추출물의 항암 효과는 균주에 대한 유의적인 차이는 없었으며 발효시간 경과에 따라 감소 하는 경향을 보였으나, 비발효구에 비해서는 항암 효과가 증가하는 것으로 분석되었다.
대두가 함유하고 있는 생리활성 물질로는 phytic acid, lignin, saponin, isoflavone 등이 알려져 있다(Kim 등, 1999).
또한 대두발효 시 B. subtilis로부터 생산되는 것으로 알려 진 menaquinone(vitamin K2)은 항암작용이 있는 것으로 알 려져 있고(Buchanan 등, 2016), B. subtilis JBS17166 균 주와 B. subtilis KACC 10854 균주를 사용하여 제조한 대 두발효 추출물의 IC50 값이 대조구의 IC50 값보다 낮은 것으 로 보아 대두 자체의 생리활성 물질과 발효과정 중 생성된 물질들의 시너지효과에 의한 것으로 판단된다.
요 약
프로바이오틱스 특성을 보유한 Bacillus 속 미생물을 선발 하여 대두발효에 이용하고, 제조된 발효물의 in vitro 항비만 과 항암 활성을 평가하였다. 프로바이오틱스 특성을 평가하 기 위하여 실시한 내산성, 내담즙성, 장부착능 측정 결과, B. subtilis JBS17166 균주는 우수한 생존율 및 장부착능을 보유하는 것으로 확인되었으며 B. cereus와 S. aureus에 대한 항균 활성이 있는 것으로 확인되었다. 최종 선발된 B. subtilis JBS17166 균주를 사용하여 제조한 대두발효 추출 물(24 h, 48 h)에 대한 항비만 및 항암 활성을 평가한 결과, 3T3-L1 세포주를 활용한 항비만 효과는 대두발효 추출물 (JBS17166, 24 h) 첨가구에서 59.95%의 지방분화 억제능 이 있는 것으로 확인되었다. 암세포주(HepG2 및 HT-29)에 대한 항암 활성의 경우 B. subtilis JBS17166 균주를 이용 하여 제조한 대두발효 추출물이 비발효 추출물(control)에
비해 약 2배 작은 IC50 값을 가지는 것으로 확인되었다. 이와 같은 결과를 토대로 프로바이오틱스로서 B. subtilis JBS 17166 균주를 사용하여 발효한 대두발효물은 항비만 및 항 암 효과를 보유한 신바이오틱스로 유용할 것으로 기대된다.
감사의 글
본 연구는 2018년 산업통상자원부 산업기술거점기관지원 (한국형 유용균주, 과제번호: R0004073) 사업의 지원에 의 해 수행되었습니다.
REFERENCES
Adami A, Cavazzoni V. Occurrence of selected bacterial groups in the faeces of piglets fed with Bacillus coagulans as pro- biotic. J Basic Microbiol. 1999. 39:3-9.
Azcarate-Peril MA, Tallon R, Klaenhammer TR. Temporal gene expression and probiotic attributes of Lactobacillus acid- ophilus during growth in milk. J Dairy Sci. 2009. 92:870-886.
Barbosa TM, Serra CR, La Ragione RM, Woodward MJ, Henri- ques AO. Screening for Bacillus isolates in the broiler gastro- intestinal tract. Appl Environ Microbiol. 2005. 71:968-978.
Bengoa AA, Zavala L, Carasi P, Trejo SA, Bronsoms S, Serradell MA, et al. Simulated gastrointestinal conditions increase ad- hesion ability of Lactobacillus paracasei strains isolated from kefir to Caco-2 cells and mucin. Food Res Int. 2018. 103:462- 467.
Buchanan GS, Melvin T, Merritt B, Bishop C, Shuler FD. Vita- min K2 (menaquinone) supplementation and its benefits in cardiovascular disease, osteoporosis, and cancer. Marshall Jour- nal of Medicine. 2016. http://dx.doi.org/10.18590/mjm.2016.
vol2.iss3.8
Chouraqui JP, Grathwohl D, Labaune JM, Hascoet JM, de Mont- golfier I, Leclaire M, et al. Assessment of the safety, toler- ance, and protective effect against diarrhea of infant formulas containing mixtures of probiotics or probiotics and prebiotics in a randomized controlled trial. Am J Clin Nutr. 2008. 87:
1365-1373.
Cui C, Shen CJ, Jia G, Wang KN. Effect of dietary Bacillus subtilis on proportion of Bacteroidetes and Firmicutes in swine intestine and lipid metabolism. Genet Mol Res. 2013.
12:1766-1776.
Ding WK, Shah NP. Acid, bile, and heat tolerance of free and microencapsulated probiotic bacteria. J Food Sci. 2007. 72:
M446-M450.
Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, et al. Diversity of the human intestinal microbial flora. Science. 2005. 308:1635-1638.
FAO/WHO. Report of joint FAO/WHO working group on draft- ing guidelines for the evaluation of probiotics in food. Food and Agriculture Organization/World Health Organization, Lon- don, Canada. 2002.
Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr. 1995. 125:1401-1412.
Gourineni VP, Verghese M, Boateng J, Shackelford L, Bhat NK, Walker LT. Combinational effects of prebiotics and soybean against azoxymethane-induced colon cancer in vivo. J Nutr Metab. 2011. 2011:868197. doi: 10.1155/2011/868197.
Hong HA, Duc le H, Cutting SM. The use of bacterial spore
formers as probiotics. FEMS Microbiol Rev. 2005. 29:813- 835.
Hsu CL, Yen GC. Effects of flavonoids and phenolic acids on the inhibition of adipogenesis in 3T3-L1 adipocytes. J Agric Food Chem. 2007. 55:8404-8410.
Hwang JW, Do HJ, Kim OY, Chung JH, Lee JY, Park YS, et al. Fermented soy bean extract suppresses differentiation of 3T3-L1 preadipocytes and facilitates its glucose utilization.
J Funct Foods. 2015. 15:516-524.
Kanamori Y, Sugiyama M, Komura M, Nakahara S, Sato K, Iwanka T, et al. Synbiotic therapy: An important supportive therapy for pediatric patients with severe respiratory diseases.
Int J Prob Preb. 2006. 1:161-168.
Kim SH, Yang JL, Song YS. Physiological functions of chung- kukjang. Food Industry and Nutrition. 1999. 4(2):40-46.
Kim SY, Shin KS, Lee H. Screening of lactic acid bacteria with potent adhesive property in human colon using colonic mucin- binding assay. Korean J Food Sci Technol. 2004. 36:959-697.
Kim YH. Probiotics, prebiotics, synbiotics and human health. BT News. 2016. 23:17-22.
Lim ES, Kim YM, Lee EW. Probiotic properties and safety as- sessment of lactic acid bacteria isolated from salt-fermented anchovy. Korean J Food Sci Technol. 2016. 48:306-316.
Maftei NM. Probiotic, prebiotic and synbiotic products in human health. In: Solis-Oviedo RL, editor. Frontiers and New Trends in the Science of Fermented Food and Beverages. 1st ed.
2019. doi: 10.5772/intechopen.81553.
Markowiak P, Slizewska K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients. 2017. doi: 10.3390/
nu9091021.
Murphy PA, Song T, Buseman G, Barua K, Beecher GR, Trainer D, et al. Isoflavones in retail and institutional soy foods. J Agric Food Chem. 1999. 47:2697-2704.
Nam Y, Jung H, Karuppasamy S, Lee JY, Kang KD, Hwang KY, et al. Anti-hyperlipidemic effect of soybean extract fer- mented by Bacillus subtilis MORI in db/db mice. Lab Anim
Res. 2012. 28:123-129.
O’Bryan CA, Pak D, Crandall PG, Lee SO, Ricke SC. The role of prebiotics and probiotics in human health. J Prob Health.
2013. 1:108.
Park HJ, Oh HH, Jeong DY, Kim YS. Probiotic characteristics of Bacillus tequilensis JBC17126 isolated from Korean tradi- tional soybean paste. J Korean Soc Food Sci Nutr. 2019. 48:
710-717.
Seo JH, Lee H. Characteristics and immunomodulating activity of lactic acid bacteria for the potential probiotics. Korean J Food Sci Technol. 2007. 39:681-687.
Seo WT, Nam SH, Lee CK, Cho KM. Identification of potential Bacillus subtilis probiotics from Korean soybean paste and their antimicrobial and immune activities. J Food Sci Nutr.
2011. 16:37-44.
Stein T. Bacillus subtilis antibiotics: structures, syntheses and specific functions. Mol Microbiol. 2005. 56:845-857.
Tennyson CA, Friedman G. Microecology, obesity, and probi- otics. Curr Opin Endocrinol Diabetes Obes. 2008. 15:422- 4227.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with in- creased capacity for energy harvest. Nature. 2006. 444:1027- 1031.
Van Loo J, Clune Y, Bennett M, Collins JK. The SYNCAN project: goals, set-up, first results and settings of the human intervention study. Br J Nutr. 2005. 93:S91-S98.
Yoon JA, Shin KO. Prebiotics. Korean J Food Nutr. 2017. 30:
191-202.
Yoon SS, Park YS, Choi HJ. Genetics and research revolutions in the lactic acid bacteria: focused on probiotics and immu- nomodulation. Curr Top LAB Probiotics. 2013. 1:9-19.
Zebisch K, Voigt V, Wabitsch M, Brandsch M. Protocol for effective differentiation of 3T3-L1 cells to adipocytes. Anal Biochem. 2012. 425:88-90.