147 http://dx.doi.org/10.7850/jkso.2014.19.2.147
Free Access
동해 남부 해역 퇴적물과 저층 해수 세균 군집 조성의 계절적 변화 연구
최동한1,2
·
김병모3,4·
최태섭3·
이정석3,*·
노재훈2,5·
박영규6·
강성길71
한국해양과학기술원 해양바이오연구부
2
과학기술연합대학원대학교
3
(주)네오엔비즈 환경안전연구소
4
세종대학교 지구환경과학과
5
한국해양과학기술원 해양생태계연구부
6
한국해양과학기술원 해양순환·기후연구부
7
한국해양과학기술원 부설 선박해양플랜트연구소 해양CCS연구단
Seasonal Variation of Bacterial Community Composition in Sediments and Overlying Waters of the South East Sea
D ONG H AN CHOI
1,2, B YEONG -M O G IM
3,4, T AE S EOB C HOI
3, J UNG -S UK L EE
3,*, J AE H OON N OH
2,5, Y OUNG -G YU P ARK
6AND S EONG -G IL K ANG
71
Marine Biotechnology Research Division, Korea Institute of Ocean Science and Technology (KIOST), 787 Haean-ro, Sangnok-gu, Ansan-si, Gyeonggi-do 426-744, Korea
2
Korea University of Science and Technology, 217 Gajeong-ro, Yuseong-gu, Daejeon 305-333, Korea
3
NeoEnBiz Co. Daewoo Technopark A-1306, Dodang-dong, Bucheon-si, Kyeonggi-do 420-806, Korea
4
Department of Earth & Environmental Sciences, Sejong University, 209 Neungdong-ro, Gwangjin-gu, Seoul, 143-747, Korea
5
Marine Ecosystem Research Division, Korea Institute of Ocean Science and Technology (KIOST), 787 Haean-ro, Sangnok-gu, Ansan-si, Gyeonggi-do 426-744, Korea
6
Ocean Circulation and Climate Research Division, Korea Institute of Ocean Science and Technology (KIOST), 787 Haean-ro, Sangnok-gu, Ansan-si, Gyeonggi-do 426-744, Korea
7
Offshore CCS Research Unit, Korea Research Institute of Ships and Ocean Engineering, 32 1312beon-gil, Yuseong-daero, Yuseong-gu, Daejeon 305-343, Korea
해양 환경에서 세균은 다양한 생지화학적 순환에 있어서 중요한 역할을 수행하고 있으며, 그들의 다양성에 대한 정보는 생태계에서 세균의 생지화학적 기능을 이해하는데 중요하다. 본 연구는 CO
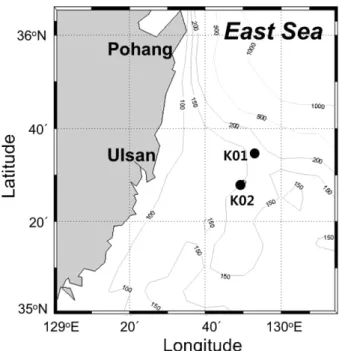
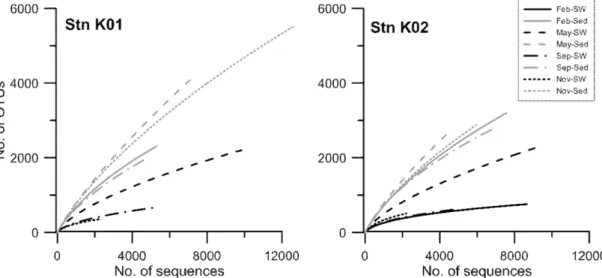
2해양지중저장의 후보지인 동해 남부해역의 표층 퇴적물과 퇴적물 위의 저층 해수에서 최신 연구 기법인 pyrosequencing을 통하여 세균의 계절적 다양성을 분석함으로써, 동해 퇴적물 세균상의 특성을 이해하고자 하였다. 퇴적물에서는 대부분의 시기에 Gammaproteobacteria 가 우점한 반면, 저층 해수에서는 Alphaproteobacteria가 우점하여 두 서식처에서 세균의 다양 성은 큰 차이를 보였다. 또한 속 수준의 다양성 분석에서도 저층 해수에서는 대부분의 시기에 SAR11 그룹에 속하 는 Pelagibacter가 가장 우점한 반면, 퇴적물 시료에서는 Gammaproteobacteria에 속하는 미동정 속이 가장 우점하 였다. 그러나 두 서식처 모두에서 5% 이상의 점유율을 보인 속의 수는 10 속 미만으로 소수였으며, 낮은 점유율을 갖는 많은 종류의 세균들이 군집 내에 공존하는 공통적인 특성이 나타났다. 본 연구의 세균 다양성 연구는 동해 저 층 해수 및 퇴적물의 세균 다양성에 대한 특성의 이해와 더불어 CO
2해양지중저장 사업의 진행에 따른 세균의 다 양성 및 기능 변화에 대한 사전 자료 및 해역이용영향평가 배경 자료로 활용될 수 있을 것이다.
Bacteria play an important role in biogeochemical cycles in marine environments and their functional attri- butes in ecosystems depend primarily on species composition. In this study, seasonal variation of bacterial diversity was investigated by pyrosequencing of 16S rDNA in surface sediment and overlying seawater col- lected in the south East Sea, planned for the site of CO
2sequestration by the carbon capture and storage (CCS) project. Gammaproteobacteria was dominant in the sediment in most seasons, whereas Alphaproteobacteria
Received February 18, 2014; Revised April 29, 2014; Accepted April 30, 2014
*Corresponding author: [email protected]