We described for the first time the freshwater dinoflagellate Unruhdinium penardii var. robustum, collected from Paldang Reservoir, Korea, on October 21, 2019. The overall shape was pentagonal to rhombic. The cell size was 31 μm in length (23~39 μm), and 29 μm in width (23~38 μm). Their plate formula was determined to 4', 0a, 6'', 5''', 0p, and 2''''; and the apical plate was symmetric. Two apical spines were prominent, and approximately 6 robust hypothecal spines were observed.
Molecular comparisons of the 18S rDNA sequences showed that our species was well-matched with the already-known U.
penardii (99.9% similarity); because U. penardii var. robustum 18S rDNA sequence was not available in the public database.
Additional 28S rDNA sequences had high similarity with already-known U. penardii var. robustum (99.8% similarity).
These morphological and molecular results confirmed that our dinoflagellate was clearly U. penardii var. robustum, and it was recorded for the first time in Korean freshwaters.
Keywords: Unruhdinium penardii, 18S rDNA, freshwater dino- flagellate, morphology, Paldang Reservoir
The dinoflagellates are a large group of primary producers in aquatic environments. Most are marine plankton, but they are
also present in freshwater habitats. Morphologically, they are divided into two forms (i.g., thecate and athecate); among the thecate, plate formula is an important key character to identify species morphologically. Until now, at least 3,561 species have been reported in the dinoflagellates, and about 452 species are described in freshwaters (Guiry and Guiry, 2020). The genus Peridiniopsis Lemmermann are one of the major freshwater dinoflagellates, and they are divided in nine groups by morphological characteristics like plate formula (Popovsky and Pfiester, 1990). Recently, the group Penardii was separated to be formed a new genus, Unruhdinium Gottschling, since they have distinct plate formula and diatom endosymbiont unlike other Peridiniopsis (Gottschling et al., 2017).
In Korea, the dinoflagellates have been studied for long time, because they are responsible for harmful algal blooms (Lee, 1999). Most studies have been carried out targeting marine toxic dinoflagellates, and extremely limited studies have been conducted in freshwaters. Upon searching the latest national database (National species list of Korea, 2019), we identified about 24 freshwater dinoflagellates, including the genus Ceratium Schrank, Peridiniosis Lemm, Parvodinium Carty, Peridinium Ehrenberg. It was just 5% when compared to the world records (Guiry and Guiry, 2020). Of them, Ceratium
New record of the dinoflagellate Unruhdinium penardii var. robustum (Dinophyceae) from Paldang Reservoir, Korea
Taehee Kim, Yeon-Su Lee, and Jang-Seu Ki*
Department of Life Science, Sangmyung University, Seoul 03016, Republic of Korea
팔당댐의 담수 와편모조류 Unruhdinium penardii var. robustum (Dinophyceae)에 관한 미기록종 연구
김태희 ・ 이연수 ・ 기장서*
상명대학교 생명과학과
(Received April 10, 2020; Revised May 20, 2020; Accepted May 20, 2020)
*For correspondence. E-mail: [email protected];
Tel.: +82-2-2287-5449; Fax: +82-2-2287-0070
hirundinella was firstly described from Han River (Chung et al., 1968). Since then, several freshwater dinoflagellates have been recorded via ecological surveys (Han et al., 1995; Kim, 1998; Kim et al., 2009), but their taxonomic identities have not been examined morphologically. To our knowledge, only three freshwater dinoflagellates had a morphologically accurate identification, which are Peridinium bipes f. occultatum, Peridinium aciculiferum, and Peridinium umbonatum (Ki and Han, 2005, 2008; Ki et al., 2005).
The Han River is an important water resource in Seoul and the metropolitan area, and thus many studies have been conducted on water quality managements (Yoo and Lim, 1990;
Kim et al., 2009). The Paldang Reservoir is where the three tributaries (the North Han river, the South Han River and Kyungan Stream) of the Han River join. In a recent study, we found blooms of a freshwater dinoflagellate in the Paldang Reservoir, and it was suspected to be Peridiniopsis sp. by 18S metagenomics (Boopathi and Ki, 2016); however, their taxonomic identities are still unknown to date.
In the present study, we examined morphological characters of the bloom-forming dinoflagellate collected from Paldang Reservoir, and determined their 18S and 28S rDNA sequences using molecular cloning. We compared their morphological and molecular key characters with available freshwater dino- flagellates for their accurate taxonomy.
Materials and Methods
Water sampling and preservation
Water samples were collected on October 21 2019, at Paldang Reservoir (GPS code: 37°39'15.63''N, 127°17'15.89''E;
Fig. 1) when a freshwater dinoflagellate bloomed. Water tem- perature, pH and conductivity were measured from the monitoring site in the Paldang Reservoir using the YSI 566 Multi Probe System (YSI). In addition, dissolved oxygen (DO), total nitrogen (TN), and total phosphate (TP) data were brought from the public database of the Water Environment Information System (http://water.nier.go.kr).
For morphological observations, water samples were fixed with Lugol’s solution at 1% final concentration. In addition, a total of 300 ml water sample was filtered with 10 µm-pore sized
membrane filter (Cat. No. TCTP04700, 47 mm diameter, Millipore), and this filter was stored at -20°C, until DNA extraction with cetyltrimethylammonium bromide (CATB) buffer.
Morphological observations
Morphological features of dinoflagellate cells were observed with a light and epifluorescence microscope (Carl Zeiss Axioskop) equipped with a ProgRes® CF scan CCD camera (Jenoptik).
Lugol’s solution fixed dinoflagellate cells were prepared by calcofluor white staining method (Fritz and Triemer, 1985) and the plate formula was determined using epifluorescence microscope. Average body length, width and hypotheca spine length were calculated by measuring 30 cells. Digital images were taken with a ProgRes® CF scan CCD camera, and analyzed with ProgRes CapturePro 2.10.0.1 software (Jenoptik).
DNA extraction and PCR
Genomic DNA of Unruhdinium was extracted from filtered water samples, when the cells bloomed at Paldang Reservoir.
Total DNA on filtered membranes was extracted by the CTAB method described by Richards et al. (1994). Nuclear 18S and 28S rDNA sequences were separately amplified by polymerase
Fig. 1. A map of the Paldang Reservoir, Korea. The black circle represents
the sampling site.
chain reaction (PCR) with newly designed dinoflagellate- specific 18S primers (forward: Peri-18F340, 5'-GAC GGG TAA CGG AGA ATT AG-3'; reverse: Peri-18R1690, 5'-ATT CAC CGG ATC ACT CAA TC-3') and 28S primers (forward:
Fresh-dinos-28F80, 5'-AGT AAT GGC GAA TGA ACA GG-3'; reverse: Fresh-28R910, 5'-CCT ATA CCC ATG TAT GAC GAA C-3'). PCR was carried out in 20 μl reaction mixture containing 11.8 μl of sterile distilled water, 2 μl of 10× Ex PCR buffer (TaKaRa), 2 μl of a dNTP mix (4 mM each), 1 μl of each primer (10 pmoles), 0.2 μl Ex Taq polymerase (2.5 U), and 2 μl of template. PCR was performed on a Bio-Rad iCycler with the following conditions: pre-denaturation at 94°C for 3 min;
followed by 35 cycles of 94°C for 30 sec, 55°C for 30 sec, and 72°C for 75 sec; with a final extension at 72°C for 10 min. The resulting PCR products were subjected to electrophoresis in 1.0% agarose gel (Promega), stained with Midori Green, and visualized under ultraviolet light on a transilluminator.
Cloning and sequencing
PCR products were purified with the QIAquick PCR Purification Kit (Qiagen GmbH), ligated into pTOP TA V2 vector (TOPcloner
TMTA Kit, Enzynomics), and transformed into competent cells. DNA sequencing reactions of the clones were run with the ABI PRISM® BigDye™ Terminator Cycle Sequencing Ready Reaction Kit (PE Biosystems) using the PCR products and PCR primers. After that, the remaining DNA sequences were determined by primer walking. Labeled DNA fragments were analyzed on an automated DNA sequencer (Model 3700, Applied Biosystems).
Phylogenetic and sequence similarity analyses For phylogenetic analysis, we constructed a data set of 18S rDNA sequences, including sequences of our species and those of other relatives obtained from the National Center for Biotechnology Information (NCBI) database. These sequences were aligned with MAFFT (Kuraku et al., 2013). Ambiguous regions were removed using Gblocks server with the least stringent settings (http://molevol.cmima.csic.es/castresana/G blocks_server.html). Maximum-Likelihood (ML) analysis was conducted with the same data matrix using the Tamura-Nei model in MEGA X (Kumar et al., 2018).
Additional 28S rDNA sequence of Korean Unruhdinium
was analyzed by BLAST searches in the National Center for Biotechnology Information (NCBI) and sequence similarity.
Results and Discussion
Environmental factors and biological data
In our field sampling, water temperature was recorded at 21.6°C, showing the fall characteristics of the temperate zone.
Additional environmental factors, such as pH, conductivity, DO, TN, and TP were presented in Table 1. As for phyto- plankton taxa, we identified 13 genera and 21 species, including a dinoflagellate, 5 diatoms, 7 Chlorophyta, 5 Zygnematophyceae, and 3 Euglenophyceae. The dinoflagellate Unruhdinium was composed of about 35% of the total cells. Until now, there was a little data on the appearance of the specific dinoflagellates in the Paldang Reservoir; however, previous studies had reported that certain dinoflagellates appeared mainly in the fall (Han et al., 1995; Kim et al., 2009).
Color, cell size, and shape
Cell shape of the Korean Unruhdinium sp. was pentagonal to rhombic (Fig. 2A), contrary to the oval shape of U. minimum and U. jiulongense (Zhang et al., 2014; You et al., 2015). The species presented a yellow-brownish color.
The cell was 31 μm in length (23~39 μm), and 29 μm in width (23~38 μm) (Fig. 3). Therefore, the ratio of length and width was approximately 0.94. Overall, these morphological characters were well matched with those described in Unruhdinium penardii var. robustum (Zhang et al., 2011).
However, the cell size of the Korean species was slightly smaller than the original description from Chinese freshwaters species.
Table 1. Environmental factors recorded Paldang Reservoir in October 21,
2019
Environmental factor Value
Water temperature (°C) 21.6
pH 8.1
Elecrical conductivity (μs/cm) 208
DO (mg/L) 8.6
TN (mg/L) 2.28
TP (mg/L) 0.03
General description and thecal plate formula
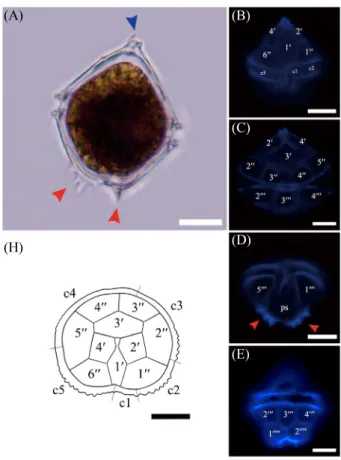
The epitheca of the Korean Unruhdinium sp. had prominent two apical spines and the plate pattern was symmetric (Fig. 2).
The plate formula of the epitheca was composed of 4 apicals, 0 intercalary, and 6 presingular plates (4', 0a, 6'') (Fig. 2B and C).
The plate 1' had a rhombic shape, and plates 2' and 4' were pentagonal. However, plate 3' was hexagonal, since this plate connected the plates 2', 4', 2'', 3'', 4'', and 5'' (Fig. 2H). In a view of the presingular plate’s shape, plate 1'', 3'', 4'', and 6'' were trapezoid, but plate 2'' and 5'' were trapezoid with 5 sides (Fig.
2H). The cingulum was composed of 5 plates (c1~c5), and c1 was very short compared to the others (Fig. 2B and H). In addition, upper and lower cingulum had some tooth shaped extension (Fig. 2H).
In addition, hypotheca plates of the Korean Unruhdinium sp.
comprised 5 postcingular and 2 antapical plates (5''', 0p, 2'''') as shown in the antapical view (Fig. 2D and E). Plates 1''', 2''', 4''', and 5''' were quadrangular, but plate 3''' was pentagonal because it connected the plates 2''', 4''', 1'''', 2'''' and the cingulum (Fig. 2E). Plate 1'''' and 2'''' were pentagonal and connected with 1''' and 5''', respectively. Approximately 6 robust hypothecal spines were observed around the plates 1'''' and 2'''' (Fig. 2A and D). The length of the hypothecal spines was 2 to 4.6 μm. All these plate formulas completely matched with those of Unruhdinium members, particularly U. penardii (Lemmermann, 1910).
Moreover, in an earlier study, Zhang et al. (2011) reported a new Peridiniopsis species (= Unruhdinium penardii var.
robustum) in a comparison with U. penardii. The plate formula of both species is the same; however, U. penardii var. robustum has two prominent apical spines and numerous robust hypothecal spines (Zhang et al., 2011). In addition, upon comparisons of Unruhdinium species spines (e.g., U. kevei, U.
niei, and U. penardii), we found that U. penardii var. robustum has the largest number of spines, and other 3 species have only 0 to 4 spines (Grigorszky et al., 2001; Liu et al., 2008; Zhang et al., 2011). Although the cell size of Korean Unruhdinium sp.
was a little small, the plate formula, the presence of epical spines and the large number hypothecal spines support that our Korean Unruhdinium was morphologically attributed to U.
penardii var. robustum.
(A) (B)
(C)
(D)
(E) (H)
Fig. 2.
Light, Epifluorescence micrographs and drawing of the Korean Unruhdinium penardii var. robustum collected from Paldang Reservoir.
(A) Entire cell shape. (B) epitheca and cingulum in ventral view. (C) epitheca in dorsal views. (D) hypotheca in ventral view. (E) hypothecain dorsal view. (H) The line drawing was based on the epifluorescence micrographs of (B) and (C). Light microscope; (A). Epifluorescence microscope; (B), (C), (D), (E). Blue and red arrows represent apical and hypothecal spines, respectively. Ps=posterior sulcal. Scale bar = 10 µm.
Fig. 3.
Body length and width of Korean Unruhdinium penardii var.
robustum. Each was determined by calculating the average measurements
of 30 cells.
Molecular affiliation of Korean Unruhdinium by 18S and 28S rDNA
With molecular cloning, we determined 1,286 bp of 18S rDNA sequences from 5 clones (GenBank accession nos.
MT273080 - MT273084). Upon comparisons, we found that all the 18S rDNA sequences were nearly identical (more than 99.9% similarity among our sequences). In addition, BLAST searches showed that our Korean Unruhdinium 18S rDNA sequences had top-hits with species from the same genus:
Unruhdinium penardii (AB353771 and HM596543), followed by U. minimum (JQ639767) and U. niei (HM596542) with high similarities (> 98.9%). Additional phylogenetic analysis of our 18S rDNA sequences and other freshwater dinoflagellates showed that the Korean Unruhdinium clustered with other Unruhdinium species and formed a sisterhood clade with two different sequences of U. penardii available in the public database (AB353771, HM596543).
In addition to this, we determined 823 bp of 28S rDNA from Unruhdinium penardii clones (GenBank no. MT453902). A BLAST search showed that our sequence had top-hit with Unruhdinium penardii var. robustum (HM596558) with
99.8% DNA similarity. This clearly supported that the Korean Unruhdinium belonged to U. penardii var. robustum genetically.
In conclusion, we examined morphological and molecular traits of Korean U. penardii var. robustum. Based on cell shape, plate formula, spines and sequence similarity, we report the first record of U. penardii var. robustum, in Korean freshwaters.
적 요
본 연구는 2019년 10월 21일 한국 팔당댐으로부터 채집한 담수 와편모조류인 Unruhdinium penardii var. robustum을 최 초로 기술하였다. 전체적인 모양은 마름모꼴의 오각형이다.
세포크기는 31 μm (23~39 μm)의 길이와 29 μm (23~38 μm)의 너비이다. 각판 배열은 4', 0a, 6'', 5''', 0p, 2''''로 관찰되었다. 상 각 각판은 대칭이다. 두개의 정단 가시(apical spines)는 뚜렷 하게 관찰되며, 하각 가시(hypothecal spines)는 약 6개가 관 찰되었다. U. penardii var. robustum은 기존에 보고된 U.
penardii의 18S rDNA 염기서열과 높은 유사도(99.9%)를 보 였고, 분자계통학적으로 근연관계를 형성하였다. 추가적인 28S rDNA 염기서열 분석에서 기존에 보고된 U. penardii var.
Fig. 4.