Original Articles Korean Circulation J 1998;;;;28((((6))))::::990-997
일시적 허혈의 반복에 의해 유도된 산소유리기에 대한 심장보호기전에 대한 연구
부산대학교 의과대학 소아과학교실
이 인 훈·박 희 주
Mechanisms of Cardioprotection to Oxygen Radicals Induced by Ischemic Preconditioning in Isolated Perfused Rat Heart
In Hoon Lee, MD and Hee Ju Park, MD
Department of Pediatrics, College of Medicine, Pusan National University, Pusan, Korea
ABSTRACT
Background:It has been known that a brief ischemic insult followed by reperfusion (‘preconditioning’) shows a protective effect on subsequent ischemic insult. The mechanism of this preconditioning was not fully understood, but oxygen free radicals, such as superoxide, hydrogen peroxide, which are released at the time of cardiac ischemia and reperfusion, were thought to have an important role in cardiac damage. We investigated the mechanism of preconditioning by examining the effects of oxidants on contractile function of isolated heart and the expression of cytokines and antioxidative genes. Methods:Isolated perfused rat hearts were exposed to a 5 minutes ischemia (1×PC) and to four times of 5 minutes ischemia, each seperated by 10 min (4×PC). Both 1×PC and 4×PC groups were reperfused for 60 minutes. Control experiments were performed by perfusiong the hearts with buffer using the same time frame without ischemia-reperfusion. The effects of oxidants in contractile function of isolated rat heart were observed. The induction of the expression of two cytokines (TNF-α and interleukin-1) and two antioxidative enzyme genes (Mn-superoxide dismutase and catalase) were examined in control, 1×PC and 4×PC hearts by RT-PCR. Results:In the control group, hydrogen peroxide or t-BHP inhibited contractile function of isolated heart as a dose-dependent manner, and IC 50 were 36.3 μM or 0.34 mM, respectively. In 4×PC hearts, inhibitory action of hydrogen peroxide on contractile function was diminished significantly, but the t-BHP action was not. In 4×PC hearts TNF-α, interleukin-1α and catalase gene expression was significantly increased compared to both 1×PC and control values. Conclusion:Repeated ischemia and reperfusion caused the induction of inflammatory and catalase genes, which may contribute to relative resistance of hydrogen peroxide in 4×PC hearts. ((((Korean Circulation J 1998;28((((6)))):990-997))))
KEY WORDS:Preconditioning·Ischemia·Cytokine·Antioxidative gene.
논문접수일:1997년 9월 22일 심사완료일:1998년 6월 25일
교신저자:이인훈, 602-060 부산광역시 서구 아미동 1가 10번지 부산대학교 의과대학 소아과학교실 전화:(051) 240-7298・전송:(051) 248-6205
E-mail:[email protected]
서 론
Superoxide, 과산화수소, 수산기 등 유리산소기(ox- ygen free radical)들은 매우 짧은 반감기와 높은 반응 성으로 심장의 수축과 전기적 특성에 영향을 미치는 것 으로 알려져 있으며1-3) 허혈-재관류에 의한 조직손상 에도 관여하는 것으로 보고되고 있다.1)4-6) 실제로 su- peroxide dismutase(SOD), catalase와 같은 유리산 소기 제거제(free radical scavenger) 투여시 유리 산 소기의 제거에 의하여 허혈-재관류에 의한 세포손상이 감소되고 수축력의 회복이 증가되는 것으로 보고되고 있다.7)8)
짧은 기간 동안의 심근 허혈이 이차적인 허혈에 대 한 심근 보호작용을 나타내는 것으로 알려져 있으며 이 를 preconditioning이라고 한다.9-11) Preconditioning 에 의한 심장보호는 이차적 허혈에 의한 조직괴사 뿐 아니라 부정맥을 감소시키는 것으로 알려져 있 다.9)12)13) Preconditioning에 의한 심장보호기전에 대 해서는 잘 알려져 있지 않으나 앞에서 기술한 바와 같 이 심장허혈시 유리산소기가 조직손상에 중요한 역할 을 하므로 preconditioning에 의해 세포내 항산화 방 어기전이 항진될 가능성이 있다. 세균에서는 catalase 와 Mn-SOD를 포함한 항산화효소들이 산화적 자극 에 의해 증가하는 것으로 보고되었다.14)15) Gorgias 등16)은 쥐의 해마(hippocampus)에서 preconditioning 에 의한 신경세포의 보호작용에 항산화효소의 활성증 가가 관여한다고 하였고, Zhou 등17) 및 Zhai 등18)은 분
리한 심장세포에서 저산소조건에 노출시킨 세포에서의 항산화효소의 활성증가를 관찰하였다. 그러나, Turrens 등19)은 토끼의 심장에서 5분간의 허혈에 의한 precon- ditioning에 의해 항산화효소의 활성의 변화가 관찰되 지 않는다고 하였고 Fu 등20)은 10분 허혈 후 30분 재 관류한 돼지의 심장에서 항산화효소의 유의한 변화를 관찰할 수 없다고 하였다.
따라서 본 실험에서는 분리된 관류심장에서 산화제 에 의한 심장수축력의 변화를 관찰하고 이들의 효과가 반복적 허혈에 의한 preconditioning에 의해 영향을 받 는지를 확인하고 이 조건에서 항산화효소 및 cytokine 들의 발현이 변화하는지를 역전사 중합연쇄반응(RT- PCR : reverse transcription polymerase chain reaction)을 이용하여 확인하고자 한다.
실험재료 및 방법
적출 심장 표본제작(Isolated langendorf heart prepa- ration)
150~250 g의 Sprague-Dawley rat를 80 mg/kg 의 pentobarbital을 복강주사하여 마취하고 심장을 신 속히 적출하여 냉한 식염수에 넣어 심장박동을 정지시 킨 후 심장을 대동맥울 통하여 삽관하고 60 mmHg의 압력으로 관류하고 Langendorf 심장 관류 장치에 걸 었다. 관류용액으로 95% O2/5% CO2를 함유한 37℃의 Krebs-Henseleit bicarbonate(KHB) 용액을 사용하였 고 KHB용액의 조성은 다음과 같았다(단위 mM) : NaCl 119, NaHCO3 25, KCl 4.6, KH2PO4 1.2, MgSO4
Table 1. Primers for polymerase chain reaction
Product Sequence of sense (S) and antisense (AS) primers Size (bp) S:5'-TGAAG CAGTGGAAGGAGCAGC-3'
Catalase
AS:5'-GCCATAGCCATTCATGTGCCG-3' 645
S:5'-CCTCAGCAATGTTGTGTCGG-3' Mn-SOD
AS:5'-CGGAAGGGCTTCACTTCTTG-3' 701
S:5'-TACTGAACTTCGGGGTGATTGGTCC-3' Tumor necrosis factor(TNF-α)
AS:5'-CAGCCTTGTCCCTTQAAGAGAACC-3' 295
S:5'-CTAAGAACTACTTCACATCCGCA-3' IL-1
AS:5'-CTGGAATAAAACCCATGAGGTAGG-3' 623
S:5'-TTGTAACCAACTGGGACGATATGG-3' β-Actin
AS:5'-GATCTTGATCTTCATGGTGCTAGG-3' 764
1.2, CaCl2 2.5, and glucose 10. 심첨부에 고리를 연 결하여 장력변환기로 수축력을 측정하였고 30분 내지 1시간후 심장의 수축력이 일정한 크기에 도달했을 때 대동맥 삽관을 차단함으로서 일정기간 동안 허혈을 유 발하거나 산화제를 투여하여 수축력의 변화를 측정하 였다.
심장조직에서 총 RNA의 분리
5분간 1회 허혈시키거나 5분간 4회 10분간격으로 반복허혈시킨 심장을 1시간 재관류한 후 조직을 분리 하여 총 RNA를 분리하였다. 조직 100 mg 당 2 ml의 RNA를 B를 가한 후 Glass-Teflon 균등분산기 (homogenizer)로 조직을 파쇄한 후 2 ml의 균등질 (homogenate)당 0.2 ml의 chloroform을 가하고 15 초간 섞은 후 12,000 g에서 15분간 원심 분리하였다.
상층액을 취하여 동일 용량의 isopropanol을 가하고 12,000 g에서 15분간 원심분리하고 침전된 RNA를 70% ethanol로 세척 후 diethylpyrocarbonate로 처 리한 멸균증류수에 용해시켜 -80℃에 저장하였다.
RNA의 양은 260 nm에서의 흡광도로 계산하고 RNA 의 통합성을 확인하기 위하여 2 μg의 RNA를 form- aldehyde agarose gel을 이용하여 전기영동하였다.
역전사 중합연쇄반응
첫 가닥의 cDNA를 합성하기 위하여 1 μg의 총 RNA를 oligo-(dT)17 시발체(primer)와 murine Mo- loney leukemia virus RT(Promega)를 이용하여 역 전사를 하였고 합성한 cDNA에 발현을 조사하고자 하 는 10 ρmole의 sense와 antisense 시발체를 첨가하 여 중합연쇄반응을 실시하였다. 중합연쇄반응을 위하여 사용한 sense 및 antisense 시발체는 Table 1에 나타 낸 바와 같다(Table 1).
95℃에서 5분간 변성시킨 후 2.5 U의 Taq 중합효 소(Promega)를 첨가하고 94℃에서 1분, 55℃에서 1 분, 72℃에서 1분으로 30주기(cycle)를 증폭하였고 β-actin의 경우 25주기를 증폭하였고 72℃에서 10 분간 최종 증폭을 실시하였다. 중합연쇄반응물질(PCR product)을 2% agarose 교화체에 전기영동하여 기대 되는 크기의 DNA가 존재하는 지를 확인하고 이를 PGEM-T vector(Promega)에 cloning한 후 T7이나 SP6 시발체를 이용하여 염기서열을 결정하였다. DNA 의 염기서열은 USB의 Sequenase v 2.0 효소를 이용 하여 이중 가닥의 dideoxynucleotide로 종료하였다.
역전사 중합연쇄반응시 동일한 양의 RNA가 사용되었는 지를 확인하기 위하여 β-actin의 시발체를 이용하여 동 일 조건에서 중합연쇄반응을 실시하였다. 중합연쇄반응
Fig. 1. Quantitative analysis of β-actin by RT-PCR. Sh- own are the value is plotted versus the total RNA. The values were normalized as the ratio to the amount of PCR products amplified for 30 cycles from 1 μg total heart RNA. Results from RT-PCR analysis with the β-actin primer pair using dilutions of the total heart RNA corresp- onding to 0.1 to 1 μg. After electrophoresis, a video image was obtained and stored by computer. The peak area of each band was determined with software.
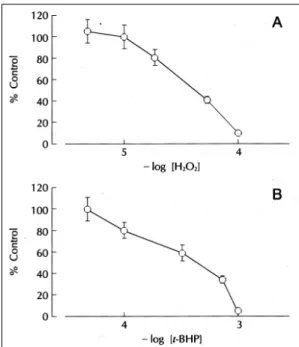
Fig. 2. Effects of oxidants in contractile function of isola- ted rat heart. Indicated concentrations of hydrogen per- oxide (A) or t-BHP (B) was added into perfusate (n=6).
물질의 정량을 위하여 DNA를 2% agarose 교화체에서 전기영동하고 ethidium bromide로 염색한 후 영상분석 기(Biorad)을 이용하여 정량하여 비교하였다.
중합연쇄반응물질의 정량분석이 가능한 중합연쇄반 응 조건을 확인하기 위하여 총 심장 RNA의 양을 변화 시키면서 중합연쇄반응 주기에 따른 β-actin의 정량 분석을 실시한 결과 25주기까지 직선적으로 증가되는 양상을 나타내었다(Fig. 1). Interleukin-1α, TNF- α, catalase 및 Mn-SOD의 경우 중합연쇄반응의 30 주기까지 RNA의 양에 따라 중합연쇄반응물질의 양이 직선적으로 증가하며 다음 실험에서는 β-actin의 경 우 25 주기를, 나머지 cDNA들의 경우 30주기를 증폭 하였다.
실험결과는 평균±평균치의 표준오차로 표시하였고 유의성 검정은 Student’s t-test를 이용하여 p치가 0.05미만인 것을 유의한 것으로 하였다.
실 험 결 과
산화제의 심장수축에 대한 영향
산화제에 의한 심장수축력의 영향을 관찰하기 위하 여 여러 농도의 과산화수소를 관류액에 첨가하여 심장 수축력의 변화를 관찰하였다(Fig. 2). 과산화수소는 20 μM의 농도로부터 수축력을 억제하였으며 50% 억제 농도는 36.3±3.2 μM이었다. t-butyl hydroperoxi- de(t-BHP)는 과산화수소와 함께 급성 산화적 요인에 의한 세포손상을 연구하기 위한 모형물질(model sub- stance)로 널리 이용되고 있다.21-24) 심장관류액에 t- BHP를 첨가하였을 때 심장의 수축력은 농도의존적으 로 감소하였으며 50% 억제농도는 0.34±0.41 mM이 었다(Fig. 2). 과산화수소와 t-BHP에 의한 심장수축 력의 감소는 약물제거에 의하여 가역적으로는 회복되 지 않았다. 이들 산화제에 의한 수축력의 감소효과가 preconditioning에 의하여 영향을 받는지를 조사하기 위하여 5분간의 전허혈(global ischemia)를 1회 가하 고 1시간 재관류시킨 군(1 PC군)과 5분간의 전허혈를 10분 간격으로 4회 반복하고 1시간 재관류한 군(4×
PC군)에서 과산화수소와 t-BHP의 효과를 측정하였다.
1회 및 4회의 반복적 허혈후에도 심장의 수축력은 1시 간 재관류후 각각 허혈을 시키기 전에 비하여 97.5±
8.6% 및 88.3±9.8%로 회복되었다. 과산화수소에 의
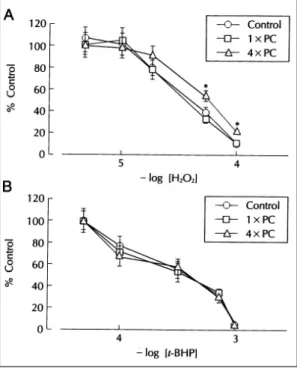
한 수축력의 억제효과는 1×PC군에서는 대조군과 유 의한 차이가 없었으나 4×PC군에서는 유의하게 감소 하여 50% 억제농도가 36.3±3.2 μM에서 61.7±7.7 μM로 증가하였다(p<0.05)(Fig. 3A). t-BHP의 효과 는 1×PC군과 4×PC군에서 대조군과 유의한 차이가 없었다(Fig. 3B).
항산화효소 및 cytokine 발현의 변화
위에서 관찰된 반복허혈에 의한 과산화수소의 심근수 축 억제작용의 감약이 반복허혈에 의한 유전자 발현의 변화에 의한 것인지를 확인하기 위하여 interleukin-1α, tumor necrosis factor-α(TNF-α), catalase, Mn- SOD, β-actin의 시발체들을 이용한 역전사 중합연쇄 반응을 실시하였다. Fig. 4에 표시된 바와 같이 35 주 기로 증폭시켰을 때 시발체로부터 예측한 것과 동일한 크기의 중합연쇄반응물질이 증폭되었으며 이들 중합연 쇄반응물질의 염기서열을 확인한 결과 기존에 알려진
Fig. 3. Effects of repetitive ischemia on hydrogen per- oxide or t-BHP action in contractile function of isolated rat heart. Isolated-perfused rat hearts were made isc- haemic for 5 min followed by 10 min reperfusion (1×
PC). Another group of experiments was performed by repeating the ischemia-reperfusion episode four times (4×PC). Indicated concentrations of hydrogen pero- xide (A) or t-BHP (B) was added into perfusate. *p<0.05 compared with the control group (n=6)
cDNA와 일치하는 것으로 밝혀졌다(Fig. 4).
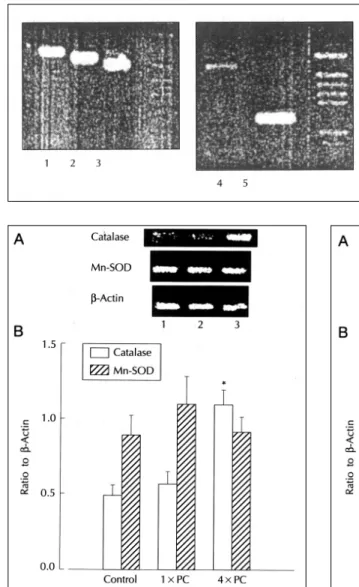
항산화효소인 catalase 및 Mn-SOD의 발현변화 를 실험방법에서 언급한 조건으로 역전사 중합연쇄반 응에 의하여 분석한 결과, catalase의 경우 1×PC군 은 대조군과 유의한 차이가 없었으나 4×PC군에서 발현이 유의하게 증가하였다. Mn-SOD의 발현은 3 군에서 유의한 차이가 관찰되지 않았다(Fig. 5). 이러 한 결과가 cytokine의 발현 변화와 상관관계가 있는 지를 확인하기 위하여 TNF-α와 interleukin-1α
의 발현 변화를 분석한 결과 4×PC군에서 이들의 발 현이 대조군과 1×PC군에 비하여 유의하게 증가됨이 관찰되었다(Fig. 6).
고 안
허혈이나 심근의 저산소증을 동반한 조건에서 항산 화 효소의 활성감소가 보고되어 있고4)25) 허혈후 재관 류시 유리 산소기의 생성이 과도하게 발생되어26) 심근 Fig. 4. PCR amplification of TNF-α, inte- rleukin-1α, catalase, Mn-SOD and β- actin cDNA samples generated by rev- erse transcription of total RNA isolated from rat heart. Amplification products were run on 2% agarose and stained with ethidium bromide. Lane 1, β-ac- tin;lane 2, Mn-SOD;lane 3, cat-alase;
lane 4, interleukin-1α;lane 5, TNF-α.
PGEM DNA marker (Pro-mega) was used as a DNA molecular weight marker.
Fig. 5. A) Representative photograph of catalase and Mn-SOD gene products amplified by RT-PCR. Lane 1, control;lane 2, 1×PC;lane 3, 4×PC. B) Quantita- tion of catalase and Mn-SOD gene expression after re- petitive ischemia in heart. The ratio of gene products to the internal standard, α-actin gene product was used to quantitate the message level. *p<0.05 comp- ared with the control group (n=5)
Fig. 6. A) Representative photograph of interleukin-1 α and TNF-α gene products amplified by RT-PCR.
Lane 1, control;lane 2, 1×PC;lane 3, 4×PC. B) Qu- antitation of interleukin-1 and TNF-α gene expression after repetitive ischemia in heart. The ratio of gene products to the internal standard, β-actin gene product was used to quantitate the message level.
*p<0.05 compared with the control group (n=4)
세포의 산화성 손상을 유발하게 되는 것으로 알려져 있 다.27) 본 실험에서는 허혈성‘preconditioning’이 반응 성 산소 중간물질(reactive oxygen intermediates)에 대한 심근의 저항성 증가에 의한 것인지를 확인하기 위 한 것이다. 이를 위하여 심근의 수축력에 대한 과산화 수소 및 t-BHP의 효과를 관찰하였고 이들의 작용에 대한 반복적 허혈의 효과를 관찰하였으며 이들 조건에 서 cytokine 및 항산화효소의 발현이 변화하는지를 확 인하였다.
본 실험에서 Turrens 등19)의 실험결과와 같이 5분 간의 허혈에 1회 노출시는 유의한 효과가 없었으나 일 시적 허혈의 반복에 의해 catalase의 RNA양이 증가되 었다. Chandrasekar 등28)은 15분 허혈후 1시간 재관 류한 심장에서 항산화효소의 발현증가를 관찰하였다.
항산화효소의 발현에 대한 이러한 실험결과들의 차이 는 명확히 설명하기 어려우나 허혈 및 재관류시간 등의 실험조건의 차이에 의하여 기인된 것으로 생각된다. 반 복허혈을 가한 군에서 과산화수소처리에 의해 나타나 는 심근수축력의 농도의존적 감소작용이 대조군에 비 하여 유의하게 억제되었으며 내재성 항산화효소인 catalase의 발현도 증가되었다. 본 실험에서 catalase 의 활성을 직접 측정하지는 않았으나 본 실험에서 RT-PCR에 의하여 평가된 RNA의 양은 steady state level을 나타내는 것으로 catalase 단백질량의 변화와 동반될 수 있는 것으로 생각된다. 실제로 심근세포에서 5분 anoxia후 1시간 reoxygenation에 의해서도 황산 화효소의 활성증가되는 것으로 보고되었다.17) 이러한 내재성 항산화 효소의 활성증가가 유리 산소기에 의한 심근손상을 방지하는 예는 이미 신생동물의 심근29)이 나 비후된 심근30)에서 보고된 바 있다. 본 실험에서 catalase와 달리 Mn-SOD의 RNA양은 반복적 허혈에 의한 preconditioning에 의하여 영향을 받지 않았으나 이미 만들어진 Mn-SOD의 활성화에 의한 Mn-SOD 의 활성증가가 동반될 가능성은 배제할 수 없다.
본 실험에서 t-BHP에 의한 심장수축작용의 억제는 과산화수소와는 달리 반복허혈에 의한 preconditioning 에 의해 영향을 받지 않았다. 이러한 차이는 t-BHP가 세포내에서 수산기가 아닌 알콕시기(alkoxyl radical) 및 메틸기(methyl radical) 등으로 대사되는 것과 관련 이 있는 것으로 생각된다.31) 실제로 t-BHP에 의한 세 포기능의 저해효과는 과산화수소와 달리 catalase에 의
하여 영향을 적게 받는 것으로 알려져 있다.22)32) TNF-α와 같은 cytokine들이 허혈에 의한 심근손 상에 관여하는 것으로 알려져 왔으나 최근 TNF-α 및 interleukin-1α 등의 염증매개물질이 심근 항산화 효소의 활성을 증가시키고33) interleukin-1α 전처치 시 심근허혈에 의한 조직손상이 감소되며 이 작용이 interleukin-1α 전처치에 의한 항산화효소의 활성증 가와 관련이 있는 것으로 알려져 이들 cytokine들이 허혈-재관류에 의한 조직손상에 대한 내용성(tolera- nce state)을 유도하는 것으로 보고되어 있다.34) 이와 유사한 효과가 폐조직35)과 간세포36)에서도 관찰된 바 있다. 본 실험에서도 반복허혈에 의해 이들 cytokine의 발현증가가 확인되었다(Fig. 6). 물론 이들 cytokine의 발현 증가가 직접적으로 catalase의 RNA발현증가에 관여하는지, 서로 독립적으로 작용하는 지는 본 실험의 결과만으로는 확인하기 어려우나 이들의 발현증가가 산화제에 대한 심장보호작용에 관여할 가능성이 있다.
요 약
연구배경:
짧은 기간 동안의 심근 허혈이 이차적인 허혈에 대한 심근 보호작용을 나타내는데 이를 Preconditioning이 라고 한다. 이러한 반복허혈에 의한 심근 보호작용의 기전은 잘 알려져 있지 않으나 심장허혈시 유리산소기 가 조직손상에 중요한 역할을 하는 것으로 알려져 세포 내 항산화효소들의 증가가 그 기전으로 작용할 가능성 이 많다. 저자들은 Preconditioning의 기전을 규명하기 위하여 심근수축력에 대한 과산화수소나 t-butyl hy- droperoxide(t-BHP)의 작용 효과와 cytokine 및 항 산화효소의 발현에 대한 반복허혈의 효과에 대한 실험 을 하였다.
방 법:
Sprague-Dawley rat의 심장을 분리하여 Langen- dorf 관류장치에서 관류하고 5분간 허혈후 1시간 재관 류한 군(1×PC군), 5분 허혈 -10분 재관류를 4회 반 복하고 1시간 재관류한 군(4×PC군) 및 대조군으로 나누어 실시하였다. 각 군에서 산화제에 의한 심장의 수축력의 변화를 관찰하였으며 각 군의 심장조직에서 cytokine(TNF-α, interleukin-1)과 항산화효소(Mn- superoxide dimutase, catalase)의 유전자 발현을 역전
사 중합연쇄반응분석에 의해 조사하였다.
결 과:
대조군에서 심근의 수축력은 심장관류액에 과산화수 소나 t-BHP의 첨가에 의하여 농도의존적으로 감소하 였고 50% 억제농도는 각각 36.3 μM 및 0.34 mM이 었다. 4×PC군에서만 과산화수소에 의한 수축력의 감 소효과가 대조군 및 1×PC군에 비해 유의하게 감약되 었으나 t-BHP에 의한 효과는 3군간에 유의한 차이가 없었다. 역전사 중합연쇄반응분석의 결과는 4×PC군에 서 TNF-α, interleukin-1α 및 catalase의 RNA양 이 대조군 및 1×PC군에 비해 유의하게 증가됨이 확 인되었다.
결 론:
심근의 반복적 허혈에 의한 preconditioning에 의하 여 과산화수소에 대한 심장수축억제작용이 감약되었고 cytokine 및 항산화효소의 유전자 발현이 증가되었다.
중심 단어:Preconditioning・허혈・Cytokine・항산 화효소.
REFERENCES
1) Hess ML, Manson NH, Okabe E. Involvement of free radicals in the pathophysiology of ischemic heart disease.
Can J Physiol Pharmacol 1982;60:1382-9.
2) Gupta M, Singal PK. Time course of structure, function and metabolic changes due to an exogenous source of ox- ygen metabolites in rat heart. Can J Physiol Pharmacol 1989;67:1549-59.
3) Beresewicz A, Horackova M. Alterations in electrical and contractile behaviour of isolated cardiomyocytes by hydrogen peroxide. J Mol Cell Cardiol 1991;23:899-918.
4) Ferrari RC, Ceconi C, Curello S, Guarnieri C, Caldarera CM, Albertini A, Visioli O. Oxygen-mediated myocardial damage during ischemia and reperfusion–Role of the cel- lular defenses against oxygen toxicity. J Mol Cell Cardiol 1985;17:937-45.
5) Pallandi RT, Perry MA, Campbell TJ. Proarrhythmic effects of an oxygen-derived free radical generating sytem on action potentials recorded from guinea pig ventricular myocardium. Br J Pharmacol 1987;91:49-59.
6) Werns SW, Lucchessi BR. The role of the polymorphon- uclear leukocytes in mediating myocardial reperfusion injury. –In Oxygen Radicals in the Pathophysiology of Heart Disease, edited by Singal PK and Boston MA: Klu- wer Academic;1988, p.122-44.
7) Shlafer M, Kane PF, Kirsh M. Superoxide dismutase plus catalase enhances the efficacy of hypothermic cardi- oplegia to protect the globally ischemic, reperfused heart.
J Thorac Cardiovasc Surg 1982;83:830-9.
8) Jolly SR, Kane WJ, Bailie MB, A Abrams GD, Lucc- hesi BR. Canine myocardial reperfusion injury. Its redu-
ctiion by the combined administratioin of superoxide di- smutase and catalase. Circ Res 1984;54:277-85.
9) Murry CE, Jennings RB, Reimer KA. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986;74:1124-36.
10) Downey JM. Ischemic preconditioning: Nature’s own cardio-protective intervention. Trends Cardiovasc Med 1992;2:170-6.
11) Warker DM, Yellon DM. Ischemic preconditioning: From mechanisms to exploitation. Cardiovasc Res 1992; 26:734-9.
12) Murry CE, Richard VJ, Jennings RB and Reimer KA.
Myocardial protection is lost before contractile function recovers from ischemic preconditioning. Am J Physiol 1991;260:790-804.
13) Liu GS, Thornton J, van Winkle DM, Stanley AWH, Olson RA, Downey JM. Protection against infarction afforded by pre-conditioning is mediated by A 1-aden- osine receptors in the rabbit heart. Circulation 1991;84: 350-6.
14) Stevens JB, Autor AP. Induction of superoxide dismutase by oxygen in neonatal rat lung. J Biol Chem 1977;252: 3509-14.
15) Richter HE, Loewen PC. Induction of catalase in Es- cherichia coli by ascorbic acid involves hydrogen peroxide.
Biochem Biophys Res Commun 1981;100:1039-46.
16) Gorgias N, Maidatsi P, Tsolaki M, Alvanou A, Kiriazis G, Kaidoglou K, Giala M. Hypoxic pretreatment protects against neuronal damage of the rat hippocampus induced by severe hypoxia. Brain Res 1996;714:215-25.
17) Zhou X, Zhai X, Ashraf M. Direct evidence that initial oxidative stress triggered by preconditioning contributes to second window of protection by endogenous antiox- idant enzyme in myocytes. Circulation 1996;93:1177-84.
18) Zhai X, Zhou X, Ashraf M. Late ischemic preconditioning is mediated in myocytes by enhanced endogenous antio- xidant activity stimulated by oxygen-derived free radicals.
Ann NY Acad Sci 1996;793:156-66.
19) Turrens JF, Thornton J, Barnard ML, Snyder S, Liu G, Downey JM. Protection from reperfusion injury by pre- conditioning hearts does not involve increased antioxid- ant defenses. Am J Physiol 1992;262:585-9.
20) Fu LX, Kirkeboen KA, Liang QM, Sjogren KG, Hjalm- arson A, Ilebekk A. Free radical scavenging enzymes and G-protein mediated receptor signalling systems in is- chaemically preconditioned porcine myocardium. Card- iovasc Res 1993;27:612-6.
21) Rush GF, Gorski JR, Ripple MG, Sominski J, Bugelski P, Hewitt WR. Organic hydroperoxide-induced lipid per- oxidation and cell death in isolated hepatocytes. Toxicol Appl Pharmacol 1985;78:473-83.
22) Stark PE, Farber JL. Ferric iron and superoxide ions are required for the killing of cultured hepatocytes by hyd- rogen peroxide. Evidence for the participation of hydr- oxyl radicals formed by an iron-catalyzed Haber-Weiss reaction. J Biol Chem 1985;260:10099-104.
23) Schnellmann RG. Mechanisms of t-butyl hydroperoxide- induced toxicity to rabbit renal proximal tubules. Am J Physiol 1988;255:28-33.
24) Chen Q, Stevens JL. Inhibition of iodoacetamide and t- butyl hydroperoxide toxicity in LLC-PK 1 cells by antiox-
idants. Arch Biochem Biophys 1991;284:422-30.
25) Dhaliwal H, Kirshenbaum LA, Randhawa, Singal PK.
Correlation between antioxidant changes during hypoxia and recovery on reoxygenation. Am J Physiol 1991;261: 632-8.
26) Bolli R, Jeroudi MO, Patel BS, Aruoma OI, Halliwell B, Lai EK, McCay PB. Marked reduction of free radical generation and contractile dysfunction by antioxidant therapy began at the time of reperfusion-evidence that myocardial stunning is a manifestation of reperfusion inj- ury. Circ Res 1989;65:607-22.
27) Singal PK, Kirshenbaum LA. A relative deficit in antio- xidant reserve may contribute in cardiac failure. Can J Cardiol 1990;6:47-9.
28) Chandrasekar B, Colston JT, Freeman GL. Induction of proinflammatory cytokine and antioxidant enzyme gene expression following brief myocardial ischaemia. Clin Exp Immunol 1997;108:346-51.
29) Rowland RT, Meng X, Ao L, Terada LS, Harken AH, Brown JM. Mechanisms of immature myocardial toler- ance to ischemia: Phenotypic differences in antioxidants, stress proteins, and oxidases. Surgery 1995;118:446-52.
30) Kirshenbaum LA, Singal PK. Increase in endogenous antioxidant enzymes protects hearts against reperfusion injury. Am J Physiol 1993;265:484-93.
31) Massa EM, Giulivi C. Alkoxyl and methyl radical form- ation during cleavage of t-butyl hydroperoxide by a mit- ochondrial membrane-bound, redox active copper pool: An EPR study. Free Radic Biol Med 1993;14:559-65.
32) Runge MM, Frank P, Novak RF. Differential effects of organic hydroperoxides and hydrogen peroxide on prote- olysis in human erythrocytes. Chem Res Toxicol 1989;2: 76-83.
33) Wong GHW, Goeddel DV. Induction of manganous sup- eroxide dismutase by tumor necrosis factor: Possible pro- tective mechanism. Science Wash DC 1988;242:941-3.
34) Maulik N, Engelman RM, Wei Z, Lu D, Rousou JA, Das DK. Interleukin-1α preconditioning reduces myoca- rdial ischemia reperfusion injury. Circulation 1993;88: 387-94.
35) White CW, Ghezzi P, Dinarello CA, Caldwell SA, Mc- Murtry IF, Repine JE. Recombinant tumor necrosis factor/cachectin and interleukin 1 pretreatment decreases lung oxidized glutathione accumulation, lung injury, and mortality in rats exposed to hyperoxia. J Clin Invest 1987; 79:1868-73.
36) Imanishi H, Scales WE, Campbell DA Jr. Tumor nec- rosis factor alpha alters the cytotoxic effect of hydrogen peroxide in cultured hepatocytes. Biochem Biophys Res Commun 1997;230:120-4.