pISSN 1598-642X eISSN 2234-7305
메주로부터 지질분해 효소 생산 균주의 분리 및 배양학적 특성
윤혜주1,2·이유정1·여수환1·최혜선1·박혜영1·박희동2·백성열1*
1농촌진흥청 국립농업과학원 농식품자원부 발효이용과, 2경북대학교 농업생명과학대학 식품공학과
Received : May 10, 2012 / Revised : June 12, 2012 / Accepted : June 13, 2012
The Isolation and Culture Characterization of a Lipolytic Enzyme Producing Strain from Meju. Yun, Hye Ju1,2, You Jung Lee1, Soo-Hwan Yeo1,Hye Sun Choi1, Hye Young Park1, Heui-Dong Park2, Seong Yeol Baek1*. 1Fermentation & Food Processing Division, Department of Agro-food Resource, NAAS, RDA, Suwon 441-853, Korea, 2Department of Food Science & Technology, Agro Biotechnology Education Center, Kyungpook National University, Daegu 702-701, Korea − For screening of useful enzymes producing microor- ganisms from Meju, we isolated high lipase producing strains and their lipolytic enzyme activities were then tested. The lipolytic enzyme activities of isolated microorganisms were therefore tested on the Y124 strain.
The gene sequence analysis of ITS from Y124 strain revealed Yarrowia lipolytica. Lipase production by the Y124 strain was studied in media containing various carbon sources. The Y124 strain drastically increased lipolytic enzyme activity in YPO media containing olive oil, as well as in YPDO media containing both olive oil and glucose. Maximal lipase production was achieved in YPD (yeast extract-peptone-D-glucose) media containing 0.7% olive oil when cultured at 30oC for 8 hrs. The lipase produced from the Y124 strain showed the highest activity in p-NPO (p-nitrophenyl octanoate (C8)), amongst the various p-nitrophenyl esters.
Keywords: Yarrowia lipolytica, 16S rDNA, lipase activity, carbon source
서 론
Lipase(glycerol ester hydrolase, triacylglycerol lipase, E.C.
3.1.1.3)는 triglyceride의 ester 결합을 가수분해하여 글리세 롤과 지방산을 생성하는 효소이다[3]. 이 효소는 동물의 췌 액에서 처음으로 발견 이후 폐, 신장, 부신, 지방조직, 태반 등과 식물의 밀, 아주까리, 콩 등의 종자에 존재하며, 곰팡 이, 효모, 세균 등 미생물성 lipase도 발견되어 생물계 전반 에 널리 분포되어 있다[11].
미생물 중 lipase를 생산하는 것으로 알려진 세균으로는 Staphylococcus 속, Chromobacterium 속, Achromobacter 속, Micrococcus 속, Bacillus 속, Pseudomonas 속, Alcaligenes 속 등이 있으며[12], 효모로는 Candida rugosa, Yarrowia lipolytica 그리고 Geotrichium candidum 등이 있다[1, 4, 13]. 미생물이 생산하는 lipase는 다양한 기질특이성, 내열성, 유기용매내성 및 입체특이성 등을 나타내어 식품 첨가물(food additive material)과 산업용 시약(industrial reagents), 세탁 첨가제(detergent additives), 의약용 진단시약(diagnostic reagent), 유지제품(cocoa butter equivalent) 등 산업적으로
다양한 분야에서 이용되고 있다[5, 7, 18].
Lipase의 기질특이성과 입체이성질체에 대한 선택성을 이 용한 가수분해, 에스테르화 및 트란스에스테르화 반응으로 서 의약적으로 중요한 키랄물질을 생산하는 연구가 이루어 졌다[6, 15]. 미생물 유래 lipase는 공급되는 영양소에 따라 기질특이성과 작용기작 등이 다양하고[14], lipase의 역가도 배양조건에 따라 차이가 있다. 미생물에서 lipase 생산성에 관한 연구가 활발히 진행되고 있으며 앞으로도 더욱 그 필 요성이 요구된다. 산업적 및 상업적으로 유용 lipase는 생산 단가가 높으며, 고온이나 유기용매 상에서 불활성화 되는 단 점이 있어 이를 극복하기 위해서는 lipase의 발현 시스템 구 축과 효소 특성에 대한 분석도 필요하다[8].
본 연구에서는 한국 전통 메주로부터 lipase를 고생산하는 효모 균주를 분리·동정하였고, 이들 균주가 생산하는 lipase 의 특성과 배양 최적 조건을 조사하였다.
재료 및 방법 시료
본 실험에 사용한 시료는 경기도 지역에서 제조된 메주를 수집하여 사용하였다. 분석에 사용한 시약은 Sigma aldrich (Sigma, ST. Louis, USA)에서 구입하여 사용하였다.
*Corresponding author
Tel: +82-31-299-0581, Fax: +82-31-299-0554 E-mail: [email protected]
균주 배양 및 선발
수집한 메주 시료(10 g)를 0.85% NaCl(90 mL)용액으로 현탁하여 YPD 고체배지(yeast extract 1%, peptone 2%, dextrose 2%)에 100 µL 도말하여 30oC에서 24시간 배양하 였다. 배양된 미생물의 형태적 차이를 이용하여 1차 선별한 다음, TBN-LB plate (200 mM NaCl, 10 mM CaCl2와 5%
(w/v) gum arabic solution이 포함된 45 mL solution에 tributyrin 5 mL을 첨가하여 waring blender로 2분간 mixing 한 다음 멸균 후 LB agar 배지 450 mL과 혼합)에 접종한 후[9], clear zone을 생성하는 균주를 선발하여 사용하였다.
지질분해 효소 생산 균주의 동정
DNA 유전자 염기서열 분석은 Solutions for Generic Technologies (Solgent) 사에 의뢰하여 분석하였으며, primer 로는 internal transcribed spacer (ITS) 영역의 Universal primer인 ITS1 (5'-TCCGTAGGTGAACCTGCGG-3')와 ITS4 (5'-TCCTCCGCTTATTGATATGC-3')를 사용하였다. Phylo- genetic tree 작성은 Lasergene사의 DNASTAR pro software (SeqMan Pro)와 The National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov/)에서 제공 하는 Advanced blast search 프로그램을 통하여 GenBank에 보고된 유사 균주와의 염기서열을 비교하여 계통분류학적 유연관계를 분석한 후, MEGA v4.0을 이용하여 Tamura-Nei distance model과 neighbor-joining method [17]에 의해 계 통도를 작성하였다.
지질분해 효소 활성 측정
Lipase 활성 측정은 microplate assay 방법[16]으로 측정 하였다. microplate assay는 p-nitrophenyl ester (pNP-ester) 를 기질로 사용하여 유리되는 p-nitrophenol의 양을 405 nm 에서 흡광도 증가량으로 측정하였으며 반응액은 기질의 종 류에 따라 두 가지 방법을 혼합하였다. Ester carbon의 길이 가 12개 이하의 탄소수를 가진 기질(pNP-acetate)의 경우, acetonitrile로 녹인 10 mM pNP-acetate 5 µL, ethanol 5 µL, 50 mM phosphate buffer (pH 7.4) 180 µL와 효소액 10µL를 섞은 다음 37oC에서 10분간 반응 후 405 nm에서 흡광도를 측정하였고, long chine인 pNP-palmitate를 기질로 사용할 경우 10 µL의 효소액과 170 µL의 buffer 혼합액(50 mM phosphate buffer containing 0.1% gum arabic and 0.2% deoxycholate) 그리고 isoprophanol로 녹인 8 mM의 기질 20 µL를 혼합하여 동일한 조건으로 반응시켰다. 효소 의 unit 측정은 1분간 1 µmol의 p-nitrophenyl를 생성하는 효소의 양을 1 unit로 하였다.
배지에 따른 균주의 생육도 및 lipase 활성
배지에 따른 분리균의 lipase 활성을 알아보기 위해, YPD (yeast extract-peptone-D-glucose), YPO (yeast extract-peptone-
olive oil), YPDO (yeast extract-peptone-D-glucose-olive oil)배지를 사용하였다. 모든 배지는 1% yeast extract와 2%
peptone을 포함하고 YPD, YPDO는 2%의 dextrose를 포함 한다. 그리고 YPO, YPDO는 1%의 olive oil을 포함한다.
olive oil은 0.22 µm membrane filter로 필터 후, 멸균된 YP 및 YPD용액에 첨가하여 배지를 만든다. 균주를 YPD, YPDO 및 YPO 배지에 접종하여 30oC에서 72시간 동안 진 탕 배양하면서 경시적으로(0, 4, 8, 12, 24, 48, 72시간) 배 양액을 취하여 탁도(OD600)를 측정하여 균체의 생육도를 측 정하고, 10,000 rpm, 4oC에서 10분간 원심분리하여 균체를 제거하고 그 상징액을 조효소액으로 하여 lipase 효소 활성 을 microplate assay 방법으로 측정하였다. Microplate assay 는 dodecanoate (C12)를 기질로 사용하여 유리되는 p- nitrophenol의 양을 이용하여 405 nm에서의 흡광도 증가량 으로 측정하였다.
기질 농도에 따른 균주의 생육도 및 lipase 활성
기질 농도에 따른 lipase 활성을 조사하기 위해, YPDO 배 지에 olive oil을 농도별(0%, 0.3%, 0.5%, 0.7%, 1%)로 첨 가하여 배지를 만들고 분리 균주를 접종하여 30oC에서 72 시간 동안 진탕 배양하면서 경시적으로(0, 4, 8, 12, 24, 48, 72시간) 배양액을 취하여 균체의 생육도 및 lipase 효소 활 성을 측정하였다. Lipase 효소 활성은 dodecanoate (C12)를 기질로 사용하여 microplate assay 방법으로 측정하였다.
배양 온도에 따른 균주의 생육도 및 lipase 활성
배양 온도에 따른 lipase 활성을 알아보기 위해 YPO 배 지에 균주를 접종하여, 배양온도별(25oC, 30oC, 37oC)로 24 시간 동안 진탕 배양하면서 경시적으로(0, 4, 8, 12, 24시간) 배양액을 취하여 균체의 생육도 및 lipase 효소 활성을 측정 하였다. Lipase 효소 활성은 dodecanoate (C12)를 기질로 사 용하여 microplate assay 방법으로 측정하였다.
반응온도에 따른 lipase 활성
YPO 배지에 균주를 접종하여 30oC에서 8시간 동안 진탕 배양한 후 10,000 rpm, 4oC에서 10분간 원심분리하여 균체 를 제거하고 그 상징액을 조효소액으로 하였다. 균주가 생 산하는 lipase의 최적 활성 온도를 검토하기 위해 acetonitrile 로 녹인 10 mM pNP-acetate 5 µL, ethanol 5 µL, 50 mM phosphate buffer (pH 7.4) 180µL와 효소액 10 µL를 섞은 다음 반응온도를 각각 15oC, 20oC, 25oC, 30oC, 37oC로 하 여 dodecanoate (C12)를 기질로 사용하여 microplate assay 방법으로 lipase 활성을 측정하였다.
기질에 따른 lipase 활성
기질 종류에 따른 lipase 활성을 알아보기 위해 p-nitro- phenyl acetate (C2), p-nitrophenyl butyrate (C4), p-nitrophenyl
octanoate (C8), p-nitrophenyl dodecanoate (C12), p-nitro- phenyl palmitate (C16)의 기질들을 사용하였다. YPO 배지에 균주를 접종하여 30oC에서 8시간 동안 진탕 배양한 후 10,000 rpm, 4oC에서 10분간 원심분리하여 균체를 제거하고 그 상징액을 조효소액으로 하였으며 반응액은 기질의 종류 에 따라 두 가지 방법을 사용하였다. Ester carbon의 길이가 12개 이하의 탄소수를 가진 기질 (pNP-acetate)의 경우와 long chain pNP-ester인 pNP-palmitate를 기질로 사용하는 경우로 나누어 실험하였다. Lipase의 활성은 p-nitrophenyl dodecanoate (C12)에서 얻어지는 흡광도 값을 100으로 하여 다른 기질에 대한 상대적 활성을 측정하였다.
결과 및 고찰
지질분해 효소 활성이 우수한 균주 선발 및 동정
경기도 일대에서 수집한 메주 시료에서 분리한 균을 TBN- LB plate에 접종하여 clear zone을 형성하는 균주를 1차적으 로 분리하였다. 지질 분해 활성을 보유한 균주 중 가장 넓은 투명환을 보여주는 Y124 균주를 선발하였다.
Y124 균주의 ITS 염기서열을 분석한 결과, Yarrowia sp.
로 나타났다. 그리고 Y124 균주의 계통학적 유연관계를 ITS 유전자의 염기서열을 기초로 Yarrowia 속(genus)의 표준 종 들과의 similarity를 조사한 결과, Yarrowia lipolytica와 100% 상동성을 보이는 것으로 나타났다. Y124 균주와 여러 종의 Yarrowia lipolytica에 속하는 다른 종과의 유사성을 Fig. 1에 나타내었고, 표준균주(Yarrowia lipolytica KACC 41237)와 지질분해 활성을 비교한 결과, 표준균주는 지질분 해 활성이 거의 없었지만 선발균주 Y124는 높은 지질분해 활성을 나타냈다(Fig. 2).
배지에 따른 분리균의 생육도 및 lipase 활성
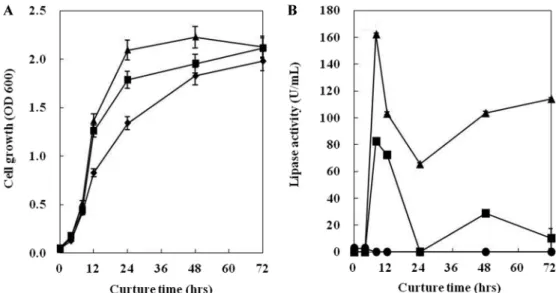
Olive oil과 glucose를 모두 포함하는 YPDO 배지와 탄소 원 소스로 olive oil과 glucose를 각각 포함하는 YPO 배지 와 YPD 배지에서 균주를 배양하였다. 탄소원으로 olive oil 을 단독으로 사용한 YPO 배지에서 8시간 배양하였을 때, lipase 활성이 가장 높게 나타났다. 그리고 균체의 생육은 48
시간까지 일정하게 증가하였고, 배양 8시간 이후에는 lipase 활성이 급격히 떨어졌다(Fig. 3). Amal Najjar[13] 등이 Yarrowia lipolytica를 YPO 배지에서 약 28시간 배양하였을 때 lipase 활성이 가장 높았다는 결과와는 차이가 있지만 동 일 배지에서 최대활성을 나타낸 것은 유사하였다. 그리고 glucose를 단독으로 사용한 YPD 배지에서는 lipase 활성이 거의 없었으며, olive oil과 glucose를 모두 포함하는 YPDO 배지에서는 lipase 활성이 YPO 배지 약 2배정도 낮았다.
Olive oil 농도에 따른 분리균의 생육도 및 lipase 활성 YPDO 배지에 olive oil을 0%, 0.3%, 0.5%, 0.7%, 1%
농도로 첨가하여 분리균 Y124의 생육도를 조사한 결과, olive oil의 농도에 따른 차이와 상관없이 배양 초기에 서서 히 증가하기 시작하여 유도기부터 정지기를 거치는 일반적 인 세포 성장곡선을 나타냈다(Fig. 4). 그리고 olive oil의 농 도에 따른 lipase 활성을 측정한 결과, olive oil 무첨가보다 첨가하였을때 lipase의 생산이 증가되었다. 따라서 lipase의 생산은 olive oil에 의해 유도되고, 0.7% olive oil을 첨가하 여 8시간 배양했을 때, lipase 활성이 134 U/mL으로 가장
Fig. 1. Phylogenetic tree based on ITS I-5.8S-ITS II DNA sequences showing the positions of the strain Y124 producing lipolytic enzymes isolated from Meju.
Fig. 2. Lipolytic activities of the strain Y124 producing lipoly- tic enzymes isolated from Meju. Symbols: A, Type strain KACC41237; 2, Isolated strain Y124.
높게 나타났다(Fig. 4). 이는 박 등[14]이 보고한 내용과는 lipase 활성에 차이가 있는 것으로 보아 미생물이 생산하는 효소의 다양성을 알 수 있다.
생육 온도에 따른 분리균의 생육도 및 lipase 활성 생육온도에 따른 분리균 Y124의 lipase 활성은 YPO배지 를 사용하였다. 30oC에 배양하였을 때, 배양 8시간에 가장 높은 활성이 나타났고, 25oC와 37oC에 배양하였을 때는 배 양 12시간에 활성이 가장 높게 나타났다(Fig. 5). 균주의 생 육도를 살펴 보면 25oC와 37oC에 배양하였을 때보다 30oC 에 배양하였을 때 배양 8시간 이후부터 급격히 증가하였고, 25oC와 37oC에 배양하였을 때는 배양 12시간 이후부터 생 육도가 증가하는 것으로 나타났다(Fig. 5). 이것으로 Y124
균주는 배양액 내 균체의 수가 어느 정도 이상이 되었을 때 lipase 생산이 유도되는 것으로 생각된다.
반응온도에 따른 lipase 활성
Y124 균주가 생산하는 lipase의 최적 활성 온도를 검토하 기 위해, 기질용액과 조효소용액을 혼합 한 다음, 반응온도 를 각각 15oC, 20oC, 25oC, 30oC, 37oC로 하여 lipase 활성 을 측정한 결과, 30oC에서 lipase 활성이 가장 높았다. 그러 나 30oC와 37oC의 반응온도에 따른 lipase 활성의 차이는 크 지 않았지만 중온(15oC, 20oC, 25oC)보다 높았다(Fig. 6).
Y124 균주가 생산하는 lipase의 최적 온도는 30oC이지만 이 보다 낮은 온도에서도 높은 활성을 나타내어 lipase의 활성 온도 범위가 넓어 Y124 균주가 생산하는 lipase는 여러 온 Fig. 3. Time course of lipolytic enzyme activities in the strain Y124 at culture medium. Symbols: A, cell growth; B, lipase activity; ●, YPD; ■, YPDO; ▲, YPO.
Fig. 4. Time course of lipolytic enzyme activities in the strain Y124 at different concentration of olive oil. Symbols: A, cell growth; B, lipase activity; Concentration of olive oil: ●, 0%; ■, 0.3%; ▲, 0.5%; □, 0.7%; ○, 1.0%.
도에서 다양하게 활용될 수 있을 것으로 생각된다.
다른 탄소 길이를 갖는 기질에 대한 lipase 활성
Y124 균주가 생산한 lipase의 기질 친화도를 확인하기 위 하여 다양한 p-nitrophenyl esters를 기질로 사용하였다. Y124 균주에서 생산하는 lipase의 경우 p-nitrophenyl octanoate (C8)에서 가장 높은 활성이 나타났으며, 두 번째로 p-nitro- phenyl palmitate (C16)에 대해 높은 활성이 나타났다(Fig.
7). 이 등[10]에 의하면 Aeromonas sp. LPB4가 생산하는 lipase는 C12나 C16보다 탄소 길이가 짧은 C3에서 C10에 대 한 높은 활성을 보여주고 있고, Suzuki 등[19]이 분리한 Pseudomonas sp. Strain B11-2에 의해 생산되는 lipase의 경우도 C4~C6 acyl chain을 갖는 기질에 대해 높은 활성을 나 타내어 Y124 균주와는 차이가 있었다. 최근에, 최 등[2]이 분
리한 Gelidibacter sp. YH333와 Vibrio sp. YH339에 의해 생 산되는 lipase는 각각 C12와 C8, C12와 C16에서 높은 활성을 보여주어 Y124균주와 같이 긴 탄소 사슬을 갖는 기질에서 보 다 높은 활성을 보여준 결과와 유사하였다. 특히 p-nitrophenyl octanoate (C8), p-nitrophenyl palmitate (C16)와 p-nitrophenyl dodecanoate (C12)에서 높은 활성을 보여준 것은 기존에 분 리된 lipase와 차이가 있어 향후 분리정제 등의 연구를 통해 효소의 특성이 규명될 필요가 있다.
Fig. 5. Time course of lipolytic enzyme activities in the strain Y124 at culture temperature. Symbols: A, cell growth; B, lipase activ- ity; ●, 25oC; ■, 30oC; ▲, 37oC.
Fig. 6. Effect of reaction temperature on activity of lipolytic
enzyme activities in the strain Y124. Fig. 7. Substrate specificity of lipolytic enzyme activities in the strain Y124 using p-nitrophenyl acetate (C2), p-nitrophenyl butyrate (C4), p-nitrophenyl octanoate (C8), p-nitrophenyl dodecanoate (C12), p-nitrophenyl palmitate (C16). O.D. 405 nm value after lipase assay with p-nitrophenyl dodecanoate (C12) was counted as 100%. Symbols: C2, acetate; C4, butyrate; C8, octanoate;
C12, dodecanoate; C16, palmitate.
요 약
경기도 일대에서 수집한 메주 시료에서 지질분해 활성을 나타내는 균주 Y124를 분리하여 동정한 결과 Yarrowia lipolytica와 100% 상동성을 보였다. 분리 균주가 생산하는 lipase의 조효소에 대한 일반적인 특성을 조사한 결과, 탄소 원으로 olive oil을 단독으로 사용한 YPO 배지에서 8시간 배양하였을 때 lipase 활성이 가장 높게 나타났다. YPD 배 지에서는 lipase 활성이 거의 없었으며, olive oil과 glucose 를 모두 포함하는 YPDO 배지에서는 lipase 활성이 YPO 배 지 보다 낮았다. 그리고 olive oil 농도에 따른 lipase 활성을 측정한 결과, olive oil 무첨가보다 0.7% 첨가하여 8시간 배 양했을 때 lipase 활성이134 U/mL으로 가장 높게 나타나 lipase의 생산이 olive oil의 첨가에 의해 유도되는 것으로 생 각된다. 생육온도에 따른 lipase 활성 측정한 결과, 30oC에 배양하였을 때 배양 8시간에 가장 높은 활성이 나타났고, 25oC와 37oC에 배양하였을 때는 배양 12시간에 활성이 가 장 높게 나타났으며, Y124균주의 lipase 활성 최적 온도는 30oC로 나타났다. 그리고 lipase의 기질 친화도를 확인한 결 과 Y124균주가 생산하는 lipase의 경우 p-nitrophenyl octanoate (C8)에서 가장 높은 활성이 나타났다.
Acknowledgements
This study was carried out with the support of “Coopera- tive Research Program for Agricultural Science & Technology Development (Project No. PJ006763)”, Rural Development Administration, Republic of Korea and “Research Program for Agricultural Science & Technology Development (Project No. PJ006764)”, National Academy of Agricultural Science, Rural Development Administration, Republic of Korea.
REFERENCES
1. Beopoulos, A., J. Cescut, R. Haddouche, J. L. Uribelarrea, C.
Molina-Jouve, and J. M. Nicaud. 2009. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 48: 375- 387.
2. Choi, C. S., S. Y. Lee, and J. H. Lee. 2009. Characterization of lipase produced from the microorganisms isolated from mud-flat. Korean J. Food & Nutr. 22: 14-19.
3. Desnuelle, P. 1972. The Lipases, In P. D. Boyer ed., pp. 575- 616, Academic Press, New York and London.
4. Fickers, P., J. M. Nicaud, C. Gaillardin, J. Destain, and P.
Thonart. 2004. Carbon and nitrogen sources modulate lipase production in the yeast Yarrowia lipolytica. J. Appl. Microbiol.
96: 742-749.
5. Guptaa, R., N. Gupta and P. Rathi. 2004. Bacterial lipases:
an overview of production, purification and biochemical
properties. Appl. Microbiol. Biotechnol. 64: 763-781.
6. Jaeger, K. E. and T. Eggert. 2002. Lipase for biotechnology.
Curr. Opin. Biotechnol. 13: 390-397.
7. Jaeger, K. E., B. W. Kijkstra, and M. T. Reetz. 1999. Bacterial biocatalysts: molecular biology, three-dimensional structures, and biotechnological applications of lipase. Annu. Rev.
Microbiol. 53: 315-351.
8. Jo, J. W., S. H. Hur, Y. S. Han, and J. Y. Kim. 2009. Isola- tion of lipase producing Bacillus subtilis and some charac- teristics of the enzyme. J. Appl. Biol. Chem. 52: 151-156.
9. Kim, H. K., Y. J. Jung, W. C. Choi, H. S. Ryu, T. K. Oh, and J. K. Lee. 2004. Sequence-based approach to finding func- tional lipases from microbial genome databases. FEMS Microbiol. Lett. 235: 349-355.
10. Lee, H. K., M. J. Ahn, S. H. Kwak, W. H. Song, and B. C.
Jeong. 2003. Purification and characterization of cold active lipase from psychrotrophic Aeromonas sp. LPB 4. J. Microbiol.
41: 22-27.
11. Lee, J. M., R. S. Kim, B. O. Kim, Y. D. Park, and I. N. Jin.
1993. Isolation of a pseudomonas aerusinosa stain producing an extracellular alkaline lipase catabolitely regulated by glucose, and purification of the lipase. Kor. J. Appl. Microbiol.
Biotechnol. 21: 161-168.
12. Lin, S. F., C. M. Chiou, C. M. Yeh, and Y. C. Tsai. 1996.
Purification and partial characterization of an alkaline lipase from Pseudomonas pseudoalcaligenes F-lll. Appl. Environ.
Microbiol. 62: 1093-1095.
13. Najjar, A., S. Robert, C. Guerin, M. Violet-Asther, and F.
Carriere. 2011. Quantitative study of lipase secretion, extracellular lipolysis, and lipid storage in the yeast Yarrowia lipolytica grown in the presence of olive oil: analogies with lipolysis in humans. Appl. Microbiol. Biotechnol. 89: 1947- 1962.
14. Park, M. H., H. J. Ryu, and K. K. Oh. 2004. Isolation of lipase producing yeast and optimization of cultivation condi- tion. Korean J. Biotechnol. Bioeng. 19: 148-153.
15. Reetz, M. T. 2002. Lipases as practical biocatalysts. Curr.
Opin. Biotechnol. 6: 145-150.
16. Ryu, H. S., H. K. Kim, W. C. Choi, M. H. Kim, S. Y. Park, N. S. Han, T. K. Oh, and J. K. Lee. 2006. New cold-adapted lipase from Photobacterium lipolyticum sp. nov. that is closely related to filamentous fungal lipases. Appl. Microbiol.
Biotechnol. 70: 321-326.
17. Saitou, N. and M. Nei. 1987. The neighbor joining-methods:
a new method for reconstructig phylogenetic trees. Mol.
Biol. Evol. 4: 406-425.
18. Schmidt-Dannert, C. 1999. Recombinant microbial lipase for biotechnological applications. Bioorg. Med. Chem. 7:
2123-2130
19. Suzuki, T., T. Nakayama, D.W. Choo, Y. Hirano, T. Kurihara, T. Nishino, and N. Esaki. 2003. Cloning, heterologous ex- pression, renaturation, and characterization of a cold-adapted esterase with unique primary structure from a psychrotroph Pseudomonas sp. strain B11-1. Protein Expr. Purify. 30:
171-178.