한천 분해효소를 생산하는 해양 미생물 SH-1의 분리 및 특성 분석
이재학1, 이순열2*
1서일대학교식품영양과
2한경대학교생명공학과
Received: November 3, 2014 / Revised: December 4, 2014 / Accepted: December 5, 2014
서 론
최근지상및지하의자원에서해양자원쪽으로관심이이 동되는상황에서해조류또한여러분야에서관심을받고있 다
.
해조류는역사적으로오랜기간동안산업용으로또한 식용으로광범위하게사용되어온자원으로해조류의구성성분은다당류가주를이룬다
[5, 15].
천연자원의고갈에따른대체에너지개발
,
즉바이오에탄올생산을위한바이오매스 원료측면으로도해조류의응용기대감이매우높은상황이다
[2, 3, 6].
또한지구표면이아닌해양에서자라는생명체는지구표면위의생명체가생산하는생리활성물질과그분포 및 종류가 다르다는 면에서도 연구의 대상이 되고 있다
[5, 15].
특히해조류의다당류의분해산물인올리고당을활용한기능성음식및웰빙음식문화가대두되며해조류를이 용하려는연구가진행되고있고최근일부올리고당이의학
적으로효과가있는것으로밝혀지며
[14, 17]
올리고당을확보하는방법에대하여관심이증대되고있다
.
현재까지다당류를분해하는방법은여러가지가사용이 되고있지만화학적방법의경우에는탄수화물대부분이화 학적성분특히알칼리나산에대하여비교적안정적이므로 유효성분의분리에어려움이있으며비특이적으로분해되는
문제점이있다
[15].
그러므로산업적으로이용할수있는선택적올리고당을획득하기위해서는효소의사용이바람직 하다고볼수있다
.
이러한점에서최근해양유래다당류의 분해효소에대한연구가활발히진행되고있으며특히분해 효소를분비하는해양미생물이탐색연구결과가보고되고 있다[5, 13, 29].
한천은홍조류의세포벽구성성분으로다당류두종류
,
즉Isolation and Characterization of Marine Bacterial Strain SH-1 Producing Agar-Degrading Enzymes
Jae-Hag Lee
1and Soon-Youl Lee
2*
1
Department of Food and Nutrition, Seoil University, Seoul 131-702, Republic of Korea
2
Department of Biotechnology, Hankyong National University, Anseong 456-749, Republic of Korea
A marine bacterial strain producing agar-degrading enzymes was isolated from a mud flat in Jeboo-do (Korea) using a selective artificial sea water (ASW) agar plate containing agar as the sole carbon source. The isolate, designated as SH-1, was gram-neg- ative, aerobic, and motile with single polar flagellum. 16S rRNA gene sequence similarity analysis showed the isolate SH-1 had the highest homology (96.5%) to marine bacterium Neiella marina J221. Cells could grow at 28-37ºC but not at 42ºC, and the agarase activity of the cell culture supernatant was higher when grown at 28ºC than when grown at 37ºC. Cells could grow when concentrations of 1-5% (w/v) NaCl were added to the growth media with the best growth observed at 3% NaCl, and the agar- degrading enzyme activity of the cell culture supernatant was best when grown at 3% NaCl-containing growth media under the conditions we examined. The crude enzyme prepared from 48-h culture broth of strain SH-1 exhibited an optimum pH and tem- perature for agar-degrading activity at 7.0 and 40ºC, respectively. Zymogram analysis of the crude supernatant and cell extract showed that strain SH-1 produced at least 3 agar-degrading enzymes with molecular weights of 15, 35, and 52 KD. Thinlayer chromatography (TLC) analysis also suggested that HS-1 produces β-agarase to degrade agarose to neoagarooligosaccha- rides.
Keywords: Agar-degrading enzyme, marine bacterium, zymogram, enzyme activity
*Corresponding author
Tel: +82-31-670-5333, Fax: +82-31-670-5333 E-mail: [email protected]
© 2014, The Korean Society for Microbiology and Biotechnology
agarose (60-80%)
와agaropectin (20-40%)
의 복합체이다[10, 20].
응고력이강하여한천은그자체로연구용및산업적으로많이응용이되었고
[8]
의약제로[14, 17],
비만방지제로서
[14],
산업용으로지지체로서많은분야에사용되고나아가서식품및과자의원료로사용되고있다
.
하지만최근 올리고당의기능성식품으로,
의료용으로의관심이증대되 며한천을분해하여부가가치를높일수있는올리고당으로 분해하고자한천분해효소에연구가활발히진행되고있다[5, 27, 29].
한천분해효소
(agarase)
는주로해양미생물이생산하지만최근해양미생물이아닌미생물도생산한다고보고되고있
다
[27].
특히홍조류에기생또는공생하는미생물이생산하는데
agarose
분해형식에따라α-(1
→3)
결합을분해하여agarobiose
를생산하는α-agarase [25]
와β-(1
→4)
결합을분 해하여neoagarooligosaccharide
를 생산하는 β-agarase
로나눌수있다
[13, 25].
한천분해효소중기능성역할이월등히높은
neoagarooligosaccharide
를생산하는β-agarase
에 연구가집중이되고있다[5, 17].
현재까지많은 β-agarase
를 생산하는미생물들이분리되어효소에대한특성이연구되고
,
그β-agarase
유전자를클로닝하여대량생산하는연구도활발히진행되고있다
[23, 24, 29].
하지만β-agarase
중에 도특히기능성면에서효능이높다고알려진neoagarotetraose
또는neoagarohexose
로분해하는β-agarase
를생산하는미생물의계속적인분리및효소의특성연구는필요하다
[5, 17].
본연구에서는한천분해효소를생산하는세균을서해제 부도의갯벌에서분리하였으며
,
분리균주SH-1
이생산하는 한천분해효소의특성을조사하였다.
재료 및 방법
미생물 배양조건 및 시약
개펄에서미생물을분리하고배양하기위하여사용된배지 는
Artificial Sea Water (ASW; MgSO
412.3 g, KCl 0.74 g, (NH
4)
2HPO
40.13 g, NaCl 17.5 g, CaCl
20.14 g, agar 15 g, Tris base 6.2 g per/1 L, pH 7.2)
였으며[4]
필요 시Bacto- peptone 10 g, Yeast extract 3 g
을 첨가하였다(ASW-YP).
배지 및 효소 활성측정을 위한 시약은 주로
Sigma (MO,
USA)
사로부터구입하여사용하였으며PCR
증폭을위한효소등은
Takara (Japan)
사로부터구입하여사용하였다.
한천 분해효소 생산 균주 분리인천제부도개펄에서다양한해조류를포함하는개펄을 채취하여멸균해수적당량을첨가하여약
10
-2-10
-5배로희 석한후ASW
한천평판배지에도말한후28
oC
에서배양하였다
. 4-7
일이지난후한천이분해되어투명한환을만들거나움푹파이는세균들을한천분해효소를생산하는균주 로간주하였고필요시
Lugol’s iodine
용액(25 g/l Iodine, 50 g/l Potassium Iodine)
으로염색하여콜로니주변에한천 을분해하는투명환을형성하는것을확인하였다.
한천분해효소를생산하는것으로확인된후에는주로
ASW-YP
를사용하여균주를순수분리하였다
.
균주의전자현미경사진은2%
의Uranyl acetate
를사용하여negative
염색방법으로투 과전자현미경(JEOL, JEM 1010, USA)
을사용하여세포의 외관모습을촬영하였다(NICEM, Seoul).
16S rRNA 유전자 염기서열 분석과 계통수(phylogenetic tree) 작성
순수분리한균체로부터
genomic DNA
를획득하여16S
rRNA
유전자단편을증폭하기위한PCR
반응의주형으로사용하였다
[19].
이때PCR
프라이머로세개의bacterial universal primer
세트(27F
와1492R, 9F
와1512R
그리고518F
와800R)
를사용하였으며,
증폭된DNA
단편의염기서 열은㈜제노텍(Daejeon, Korea)
에분석을의뢰하였다.
분석 된염기서열을바탕으로blastn
을수행하여염기서열에상 동성을보이는균주를확인하였다.
상동성을보이는표준균 주의16S rRNA
의유전자염기서열은EzTaxon server [7]
을 이용하여획득하였고가장높은상동성을보이는16S rRNA
유전자서열간의alignment
는ClustalW program [28]
을사 용하였다. Phylogenetic tree
작성은문헌을바탕으로Kimura’s two-parameter evolutionary model [16]
에의해계산하였고Neighbor-Joining [26]
방법으로작성하였다.
균주의 세포 성장과 agarase 활성 측정
균주의성장에따른
agarase
활성을관찰하기위해서,
기본적인조건으로
0.2% agarose
를첨가한ASW-YP
액체배지 에28
oC
에서진탕배양하면서12
시간간격으로샘플을채취 하였다.
온도에따른생장및효소활성을위한실험으로는28
oC, 37
oC, 42
oC
에서수행하였고, NaCl
농도에따른생장 및효소활성을위한실험으로는ASW-YP
를바탕으로하여 최종NaCl
농도를1%, 3%, 5%
로조정하여수행하였다.
균체의 성장은
600 nm
에서흡광도를측정하여결정하였고,
agarase
활성은균체를제거한샘플의상등액을이용하여0.2% agarose
를기질로하는3,5-dinitrosalicylic acid (DNS)
법[21]
으로540 nm
에서측정하였다.
모든실험은3
번반복 실험을수행하여통계처리하였다.
조효소액(crude enzyme) 준비
해수의평균염분농도인
3% NaCl
을 포함하는ASW-YP
배양액에서
28
oC
에서2
일간배양한균주배양액을원심분리 하여균체와상등액을분리하였다.
배양상등액에황산암모늄을최종
75%
가되도록첨가하여침전시켰다[11].
침전된 단백질용액을원심분리하고상등액을제거하여얻어진단 백질펠렛에25 mM Tris-buffer (pH 7.2)
를첨가하여펠렛을 용해시키고투석하여배양액조효소액을얻었다.
한편배양액으로부터회수된세포펠렛에배양액부피의
25 mM Tris buffer (pH 7.2)
를첨가하여세척하고적당량의 완충용액을첨가한세포현탁액을얻었다.
세포를파쇄하고 원심분리하여얻은상등액에황산암모늄으로단백질을침전 시켜펠렛을얻고,
여기에적당량의25 mM Tris-buffer (pH 7.2)
를첨가하여용해시키고투석하여세포질조효소액으로 사용하였다.
pH와 온도에 따른 agarase의 활성 측정
효소활성을위한최적
pH
조건을찾기위해서준비한배 양액조효소액으로두종류의buffer,
즉20 mM MOPS (pH 6.0-7.0), 20 mM Tris-Cl (pH 7.0-9.0)
를사용하여효소반응 을실시한후효소활성을측정하였다.
효소활성의최적온 도를찾기위해서20
oC
에서50
oC
까지의다양한온도조건 에서30
분간반응을유도하여활성을측정하였는데이때pH
는7.0
으로고정하고활성을측정하였다.
Zymogram assay 및 박막 크로마토그래피 분석
단백질농도는
Bio-Rad Dc protein assay kit (CA, USA)
를이용하여측정하였고standard
단백질로bovine serum albumin (BSA)
를사용하였다. SDS-PAGE
는Laemmli [18]
방법을바탕으로
10% polyacrylamide gel
를사용하였고전 기영동후commassie blue
염색을시행하였다.
효소활성을 보기 위한zymographic analysis
를위해gel
을 준비할 때agarose
를첨가하여separating gel
을만들었다[12]. SDS- PAGE
가끝난gel
은2.5% Triton X-100
용액에서40
분간세 척하여SDS
를제거하였다. SDS
가제거된gel
은agarase
효 소활성을나타낼수있는25 mM Tris buffer (pH 7.2)
에서42
oC
에서30-60
분간반응시킨후Lugol
시약이나commassie
blue
시약으로발색하여확인하였다. Agarase
에의한가수분해산물분석은
agarose 0.2%
를포함하는50 mM sodium phosphate buffer (pH 7.0)
에조효소액을첨가하여40
oC
에 서반응시킨 후Temujin
등의 방법으로[29] silica gel 60 plate (Merck, USA)
를 이용하여n-butanol-acetic acid-DDW (2:1:1)
을 전개용매로 하여Thin layer chromatography (TLC)
를수행하였다.
결과 및 고찰
SH-1 균주의 분리 및 형태학적, 유전학적 특성 분석 제부도개펄에서식하는균주들중한천분해효소를생
산하는해양미생물을분리하고자염분농도가해양의평균 염분농도와같은농도인
3%(w/v)
를포함하는artificial sea
water
에한천을넣은ASW
고체배지를이용하여한천분해환을보이는균주를선별하여
SH-1
이라고명명하였다. SH-
1
균주는ASW-YP
고체배지에서28
oC
에서24
시간배양하면약
1-2 mm
정도의비교적작은약한미색의콜로니를형성하고콜로니주위에
2-3 mm
의투명환을보인다.
분리균주
SH-1
은고체배지에접종한지1
일이후부터한천분해능을보였으며배양시간이지남에따라점점높은효소활성을 보였다
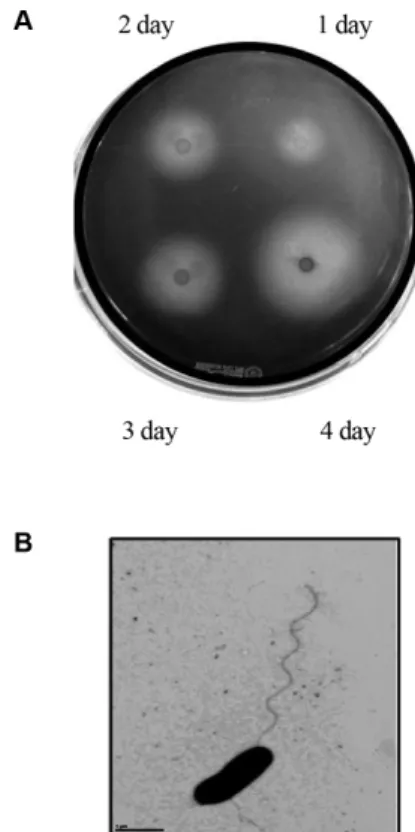
(Fig. 1A).
SH-1
균주는그람음성균이며투과전자현미경(transmission electron microscopy)
사진촬영결과포자를형성하지않는 박테리아인것으로보인다.
그크기는직경이약0.3-0.7
μm
이며그길이는1-3
μm
정도의간균이었다.
편모는한쪽극에약
10-15
μm
길이의 한개의 편모를 가지는 균이었다(Fig. 1B).
Fig. 1. Agar-degrading activity and transmission electron
micrograph (TEM) of the marine bacterium SH-1. (A) Zymo-
gram plate assay result as a function of cultivation time of the SH-
1. The strain was cultivated on ASW-YP agar plate and Lugol’s
idodine solution was overlaid to detect reducing sugars, degraded
product from agar by agarase. (B) Transmission electron micro-
graph of SH-1. The bacterial cells were removed from ASW-YP
culture plate and examined by transmission electron microscopy
after negative staining with 2% uranyl acetate. Bar, 1 μm.
분리된균주를동정하고자
16S rRNA
유전자를PCR
방 법을이용하여증폭한후그염기서열을분석하였다.
결정된 염기서열을바탕으로blastn
프로그램으로유사균들의16S
rRNA
유전자와의상동성을비교한뒤계통수를작성하였다(Fig. 2).
분리균주는Neiella marina J221
과가장높은약96.5%
의상동성을보였다. Neiella marina J221
균은2012
년 해삼(Apostichopus japonicas)
내장에서발견된균으로그람 음성균이며, 1
개에서수개의편모를가진해양미생물로보 고되었지만[9]
아직까지이와 같은 속으로보고된균주는Neiella marina H014
하나정도이다(GenBank: KJ577009.1).
그다음으로높은유사성을보인균군으로는
Pseudoalteromonas
속이약
92%
이하의상동성을보인다.
가장높은상동성을보이는
Neiella marina J221
은agarase, alginase, cellulase,
amylase
등을생산하는균으로보고되었지만[9]
본연구에서분리한
SH-1
의균체크기가Neiella marina J221
보다좀 더크고편모도하나인것을볼때분리균주SH-1
을Neiella
속으로판정하기도아직은어려운상황으로분리균주의동 정에관하여는더욱자세한연구가필요하다.
SH-1 균주가 생산하는 한천 분해 효소의 특성 분석
SH-1
균주가생산하여분비하는한천분해효소의특성을분석하고자우선균주의생장조건에따른균주의효소활성 에대하여알아보았다
. 28
oC, 37
oC
그리고42
oC
에서분리균 주를배양한결과, 37
oC
에서생장속도가다소빠른것외에 는36
시간정도가지나면28
oC
와37
oC
에서생장속도가같아졌다
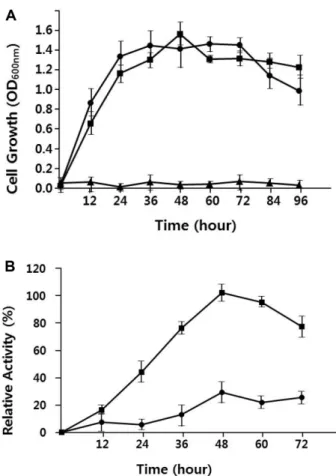
(Fig. 3A).
생장온도에따른효소활성을조사하여본결과비록생장속도는
37
oC
에서더좋은생장을보였지만 효소활성은28
oC
에서최대5
배정도의높은활성을보였다Fig. 2. Phylogenetic tree analysis of strain SH-1. Tree was constructed from the nucleotide sequence of 16S rRNA gene using the neighbor-joining method [26]. The scale bar indicates a genetic distance of 0.2. The number shown next to each node indicates the per- centage bootstrap value of 1,000 replicates.
Fig. 3. Effect of growth temperature on cell growth and aga-
rase production. (A) Comparison of cell growth in ASW-YP at the
different growth temperature (42
oC, ▲; 37
oC, ●; 28
oC, ■). (B)
Comparison of agarase activity produced by the cells grown at two
different growth temperatures (37
oC, ●; 28
oC, ■). All data are the
average of three parallel replicates.
(Fig. 3B).
배양액의염분농도에따른효소활성을조사하고자
NaCl
농도를1%, 3%,
그리고5%(w/v)
를포함하는배양액에서분 리균주를배양하며그생장곡선을그려보았다. Fig. 4A
에 서보듯이균주는3%
에서가장좋은생장을보였다.
염분농도에 따른 효소 활성을 알아본 결과
Fig. 4B
에서와 같이NaCl 3%
에서생장했을경우가NaCl 1%
에서생장했을경우보다약
1.6
배높은활성을보였다.
효소활성을위한최적조건을알아보고자배양액조효소 액을준비하여서로다른
pH
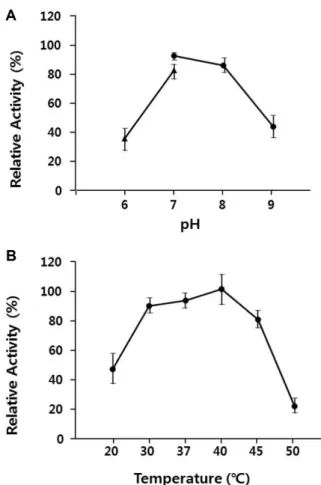
와온도에따른효소활성을조 사하였다.
다른한천분해효소와유사하게[13, 14] SH-1
이 생산하는한천분해효소는pH 7.0-8.0
사이에서가장높은활성을보였다
(Fig. 5A).
온도에따른효소활성을보기위하여
20, 30, 37, 40, 45,
그리고50
oC
에서효소활성을측정 한결과Fig. 5B
에서보듯이30-45
oC
사이에서최대활성대Fig. 4. Effect of salt concentration of the medium on cell growth and agarase production. (A) Cell growth in ASW-YP with the different NaCl concentration at 1, 3 and 5% (w/v) (1%, ▲;
3%, ●; 5%, ■). (B) Comparison of agarase activity produced by the cells grown at the different NaCl concentration (1%, ▲; 3%, ●). All data are the average of three parallel replicates.
Fig. 5. Effect of pH on agarase activity produced by SH-1 strain. (A) Enzyme activity on the different pH (pH 6-7, ▲; pH 7- 9, ●). (B) Enzyme activity on the different temperatures. All data are the average of three parallel replicates.
Fig. 6. Zymogram assay of the agarases produced from SH-
1. Lane 1, crude enzyme extract of cell supernatant; lane 2, crude
enzyme extract of the cell. Size of protein markers are indicated
on the left (in KD).
비
80%
이상의상대적인효소활성을보였으며그중40
oC
에서가장높은활성을보였다.
배양액과세포질조효소액을한천을포함하는
PAGE
이 후zymogram
분석을실시하여Fig. 6
과같은결과를얻었 다.
해양박테리아SH-1
은적어도3
개이상의한천분해효 소를생산하는것으로보이며그중가장많이발현되는것 으로보이는분해효소의분자량은약50-55, 35, 15 KD
정 도되는것으로보인다.
그중약50-55 KD
의분자량을가 지는것은세포밖으로분비되었으며약35 KD
분자량의효 소는분해효소는소량이세포밖으로분비되는것으로보인 다.
이는 β-agarase
의크기와 매우유사한 크기이므로[13, 15]
β-agarase
가분비되는것으로추정된다.
참고로대부분 의α-agarase
는85 KD
이상의크기를가지는것으로보고되고 있다
[1, 22].
또한 준비한 배양액 조효소액에 의한agarose
분해산물을TLC
분석한 결과neoagarohexose, neoagarotetrose, neoagarobiose
와같은Rf
값을가지는산 물로분해되는것으로보아분리한균주SH-1
은β-agarase
를생산하는것으로추정된다(Fig. 7).
요 약
한천을분해하는해양미생물을한천을유일한탄소원으로 하는인공해수한천배지를이용하여제부도개펄에서분
리하였다
. SH-1
으로명명한분리된균주는그람음성균이며한개의극성편모를가지는균이었다
. 16S rRNA
유전자의염기서열의유사성분석결과분리된균주는
Neiella marina
J221 [9]
과가장높은상동성을보였다(96.5%).
분리균주는28-37
oC
에서생장하였지만42
oC
에서는생장하지못하였고 한천분해효소의활성은37
oC
보다28
oC
에서높은활성을보 였다.
또한SH-1
균주는1-5% NaCl (w/v)
를포함하는배양액 에서생장이가능했으며3%
의농도에서가장좋은생장을 보였고한천을분해하는효소의활성도3%
염분농도의배 양액에서가장높았다. 48
시간배양한세포배양액을농축하 여조효소액을준비하여효소의적정pH
와적정온도를조 사한결과pH 7.0
에서와40
oC
에서최적효소활성을보였 다.
조효소액을사용하여zymogram
분석을실시한결과분 자량15, 35, 52 KD
크기의3
개이상의한천분해효소를생 산하는것으로보인다.
박막크로마토그라피(TLC)
분석결과 아가로스를분해하여네오올리고당을생성하는β-agarase
를생산하는것으로추정된다.
Acknowledgments
The present research has been conducted by the Research Grant of Seoil University in 2013.
References