326 책임저자:이창훈, 100-715, 서울시 중구 필동 3가 26
동국대학교 약학과
Tel: 02-2260-8905, Fax: 02-2260-8973 E-mail: [email protected]
접수일:2011년 10월 31일, 1차 수정일:2011년 11월 4일, 2차 수정일:2011년 11월 9일, 게재승인일:2011년 11월 14일
Correspondence to:Chang Hoon Lee
College of Pharmacy, Dongguk University, 26, Pil-dong 3-ga, Jung-gu, Seoul 100-715, Korea
Tel: +82-2-2260-8905, Fax: +82-2-2260-8973 E-mail: [email protected]
Alpinia katsumadai Suppresses Migration and 12-O-tetradecanoylphorbol-13-acetate-induced
Invasion of HT-1080 Cells through Suppression of Transglutaminase-2, Matrix Metalloproteinase-2, and
Matrix Metalloproteinase-9 Expression
Mi Kyung Park and Chang Hoon Lee
College of Pharmacy, Dongguk University, Seoul 100-715, Korea
The cause of death of the majority of cancer patients is the development of metastatic lesions at sites distant from the primary tumor. We found extracts of Alpinia katsumadai (AKE) to have anti-migratory and anti-invasion activity. Migration and invasion of HT-1080 cells were measured by using multiwell chambers. Zymography and western blots were used to examine the effect of AKE on expression of matrix metalloproteinases (MMPs) and transglutaminase-2 (tgase-2). AKE dose-dependently suppressed the migration and 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced invasion of HT-1080 sarcoma cells.
AKE did not show antiproliferative effects on 3 hr pretreatment but 24 hr incubation revealed partly cytotoxic effects of AKE on HT-1080 cells. AKE suppressed the expressions of tgase-2, MMP-2, and MMP-9 in HT-1080 cells as well as suppressed MMP-9 activity. These results suggest that migration and invasion of HT-1080 cells are inhibited by AKE via suppressions of tgase-2, MMP-2, and MMP-9 and antiproliferative activity. (Cancer Prev Res 16, 326-332, 2011)
Key Words: Alpinia katsumadai, HT-1080, Invasion, Migration, MMP-2, 9, Transglutaminase-2
INTRODUCTION
Cancer is a hyperproliferative disorder that involves trans- formation, dysregulation of apoptosis, proliferation, invasion, angiogenesis and metastasis.1) Metastasis is the continued growth and spread of a cancer from its origin to near or distant locations within the body.2) The cause of death of the vast ma- jority of cancer patients is the development of metastatic lesions at sites distant from the primary tumor.3) Thus, high mortality rates associated with cancer are functions of the metastatic spread of tumor cells from the site of origin. In fact, metastases are the cause of 90% of cancer deaths.4)
Matrix metalloproteinases (MMPs) are a zinc-dependent fam- ily of endopeptidases implicated in carcinogenesis.5) MMPs have been regarded as potential diagnostic and prognostic bio- markers in many types and stages of cancer.6) During invasion, the localization of MMPs to specialized cell surface structures, called invadopodia, is a requisite for their ability to promote invasion. These structures represent the site where active ECM degradation takes place.7)
Transglutaminase 2 (tgase-2) is a multifunctional enzyme that might act as a protein cross-linking enzyme in pro- grammed cell death; it acts as a G-protein in adrenoreceptor signaling, as a protein disulphide isomerase in maintaining mi- tochondrial physiology and as a secreted protein in stabilizing
the extra-cellular matrix ECM.8) Drug-resistant and metastatic breast cancer and melanoma cells express elevated levels of tgase-2.9,10) The invasive and drug-resistant phenotypes of MDA-MB-231 breast carcinoma cells can be suppressed when chronic tgase activity was inhibited either by knocking down tgase-2 expression using RNAi or incubating the cells with the tgase inhibitor monodansylcadaverine (MDC).9,11,12)
Alpinia katsumadai (AK) has been widely used in traditional Chinese and Korean medicine to treat a variety of conditions including emesis and gastric disorders such as gastric pain and distended abdomen. It is also known to have significant anti- oxidant activity, antiemetic activity and as an inhibitor of pros- taglandin production in LPS-stimulated mouse peritoneal macrophages.13∼15) In addition, we recently reported that AK has anti-pruritic activities in a thromboxane A2-associated mouse model and antinociceptive activity.16) However, very lit- tle is known about the anti-migratory and anti-invasion activ- ities of AK. Therefore, in the current investigation, we assessed the effects of extracts of AK (AKE) on migration and invasion of HT-1080 cells.
MATERIALS AND METHODS 1. Materials
Chemicals and reagents, unless specified otherwise, were pur- chased from Sigma-Aldrich Co. (St. Louis, MO).
2. Cell culture
HT-1080 (fibrosarcoma) cells were grown in RPMI1640 (Welgene Inc., Daegu, Korea) supplemented with 100 units/ml penicillin, 100μg/ml streptomycin, and 10% heat-inactivated fetal bovine serum (FBS; Welgene Inc.) in a humidified, 5%
CO2 incubator. Antibodies against MMP-2 and MMP-9 were obtained from Abcam (Cambridge, UK), antibody to tgase-2 (clone CUB 7402) was from NeoMarkers (Fremont, CA, USA) and antibody to β-actin was from Cell Signaling Technology (Beverly, MA, USA).
3. Preparation of AKE
Dried seeds of A. katsumadai were purchased at the Kyung- dong Oriental Drug Store (Seoul, Korea) in 2007. Preparation of AKE was performed as previously described.17)
4. Cytotoxicity of AKE
Cytotoxicity of AKE in HT-1080 cells was assessed using Ez-CyTox assay Kit (Daeillab service, Korea). Cells (100μl; 5
×103 cells/well) were seeded in 96-well plates in appropriate medium containing 10% FBS for 24 hr in a humidified, 5%
CO2 incubator. The next day, AKE or TPA (20 nM) were add- ed at various concentrations to the plates. Cells in media alone and media plus vehicle were used as negative controls. After incubation with the test agents for 24 hr, Ez-CyTox solution was added to each well of the plate. Absorbance at 460 nm was measured using a microplate reader (XMARKTM micro- plate spectrophotometer, Bio-rad, California, USA).
5. Cell migration assay
Migration assays were performed using multiwell chambers (Neuroprobe Inc. Gaithersburg, MD) coated with 10μg/ml fi- bronectin as a chemoattractant. Briefly, HT-1080 cells were suspended in RPMI at 1×106 cells/ml and a 25μl aliquot of this suspension was placed into the upper well of one chamber.
Next, an aliquot was separated from the 3%-serum-containing lower well using an 8μm polyhydrocarbon filter. After in- cubation for 3 hr at 37oC, the nonmigrated cells on the upper surface of the membrane were scrapped off, the migrated cells on the lower surface were stained with Diff-quick (Sysmax Co., Kobe, Japan). and subsequently counted under five randomly chosen high-power fields (400×).
6. Cell invasion assay
Invasion assays were performed using a 24-well transwell unit with polycarbonate filters of 6.5 mm diameter and 8.0 mm pore size (Corning Costar). Briefly, a fixed number of cells (5×104 cells/chamber) were used for each invasion assay. The lower and upper parts of the transwell were coated with 20 ml of a 1:2 mixture of Matrigel:RPMI. Cells were plated on the Matrigel-coated transwell in the presence of various con- centrations of AKE. The medium in the lower chambers con- tained 0.1 mg/ml of bovine serum albumin. After incubation for 24 hr at 37°C, cells invading the lower surface of the mem- brane were fixed with methanol and stained with hematoxylin and eosin (H&E stain), and subsequently were counted under five randomly chosen high-power fields (400×).
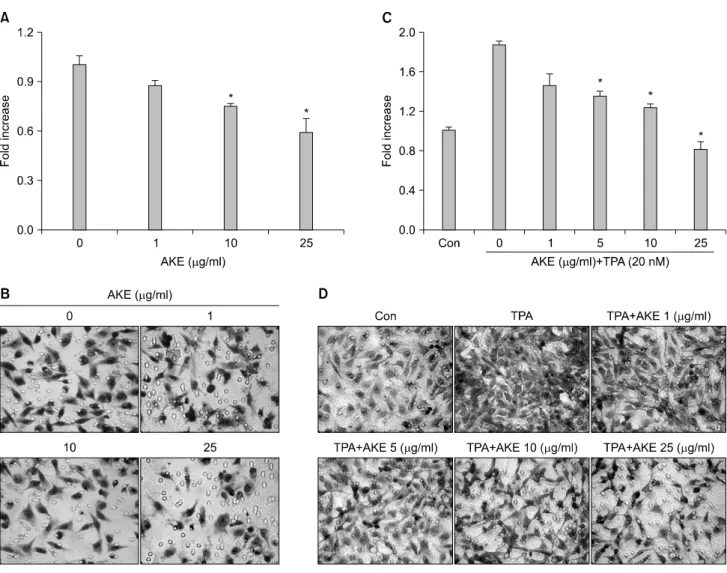
Fig. 1. AKE inhibits HT-1080 cell migration and TPA-induced invasion. (A) The inhibitory effect of AKE on the migration of HT-1080 cells. For the migration assay, the lower- and upper-chamber transwells were coated with fibronectin and HT-1080 cells were treated with vehicle (0μg/ml) or increasing concentrations (1, 10, 25μg/ml) of AKE for 3 hr. (B) Image of migrated HT-1080 cells. HT-1080 cells were allowed to migrate on fibronectin coated transwell inserts for 3 hr, stained and photographed. (C) The inhibitory effects of AKE on TPA-induced invasion. For the invasion assay, the lower- and upper-chamber transwells were coated with Matrigel. HT-1080 cells were pre-incubated with vehicle or increasing concentrations (1, 5, 10, 25μg/ml) of AKE and treated with TPA (20 nM) for 24 hr within a Matrigel invasion chamber. After 24 hr, cells on the bottom side of the filter were fixed, stained and photographed under the microscope, and the images were transferred and cells were counted as described in “Materials and methods.” The bar graphs represent the relative number of invaded cells as a percentage of vehicle from three independent experiments. *p<0.05 relative to control. (D) Image of migrated HT-1080 cells. HT-1080 cells were allowed to migrate on Matrigel coated transwell inserts for 24 hr, stained and photographed.
7. Gelatin zymography
MMP-2 and MMP-9 enzymatic activities were assayed by gelatin zymography.18) Samples of serum-free conditioned me- dium were electrophoresed on a 10% SDS-polyacryamide gel containing 0.1% gelatin. Afterwards, the gel was washed in 2.5% Triton X-100 for 1 hr and incubated at 37°C for 24 hr in activation buffer (50 mM Tris-HCl, pH 7.5, 150 mM
NaCl, 10 mM CaCl2 and 0.02% NaN3). After staining with Coomassie brilliant blue R-250, enzymatic activities were de- tected as clear bands against the blue background.
8. Western blot analysis
HT-1080 cells were cultured in RPMI media. The cells were then treated with TPA (20 nM) and AKE (1, 5, and 10μg/ml) for 24 hr. After incubation, the cells were collected and was-
washed twice with cold PBS. The cells were lysed in a lysis buffer [50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% triton X-100, 2 mM EDTA, 1% DOC (Deoxycholic acid), 0.1% SDS, 1 mM NaVO3, 10 mM NaF, 1 mM DTT] and centrifuged to yield whole-cell lysates. Proteinconcentration was measured using the Bradford method. Aliquots of the lysates (20μg of protein) were separated on a 4∼12% SDS-polyacrylamide gel and transferred onto a polyvinylidene fluoride (PVDF) mem- brane (Invitrogen, Carlsbad, CA, USA) with a glycine transfer buffer [192 mM glycine, 25 mM Tris-HCl (pH 8.8), 10%
MeOH (v/v)]. After blocking the non specific site with 3%
non-fat dry milk, the membrane was then incubated with spe- cific primary antibody in 3% BSA at 4°C for overnight. The membrane was further incubated for 60min with a perox- idase-conjugated secondary antibody (1:5,000, Santa Cruz, CA, USA) at room temperature. Immunoactive proteins were detected using the PowerOpti-ECL western blotting detection reagent (Animal Genetics Inc., Gyeonggi, Korea).
9. Statistical analysis
All of the data are presented as means±S.D. Statistical sig- nificance was analyzed using a student’s t-test; p<0.05 was considered significant when compared with the control group.
RESULTS
1. AKE suppresses migration and invasion of HT-1080 cells
Migration and invasion properties of cells are considered crit- ical in the metastatic process of cancer.19) A dose-response study was done to examine the effects of AKE on migration of HT-1080 fibrosarcoma cell lines. Treatment with AKE for 3 hr revealed a dose-dependent suppression of the migration of HT-1080 cells (Fig. 1A, B). TPA treatment of HT-1080 cells significantly increased invasion and AKE dose-dependently sup- pressed TPA-induced invasion by HT-1080 cells (Fig. 1C, D).
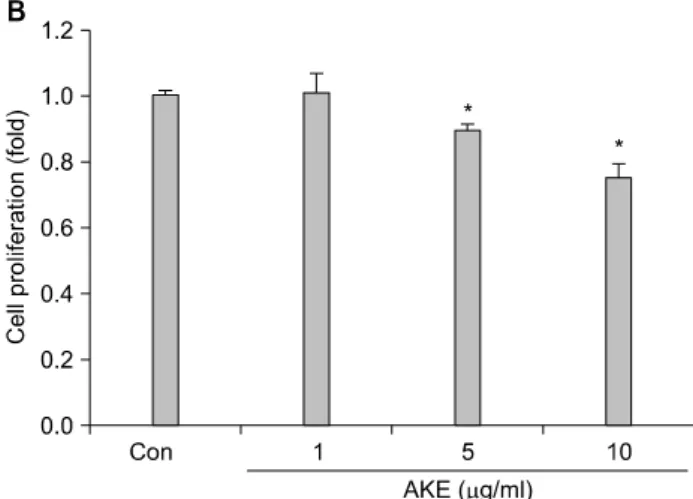
Fig. 2. Effects of AKE on cell proliferation. (A) The effect of AKE on the proliferation of HT-1080 cells for 4 hr incubation.
HT-1080 cells were cultured in RPMI media. The cells were then treated with AKE (1, 5, 10, 25μg/ml) for 3 hr. (B) The effect of AKE on the proliferation of HT-1080 cells for 24 hr incubation. HT-1080 cells were cultured in RPMI media. The cells were then treated with AKE (1, 5, 10μg/ml) for 24 hr.
(C) The effect of AKE on the proliferation of HT-1080 cells with TPA treatment for 24 hr incubation. HT-1080 cells were cultured in RPMI media. The cells were then treated with TPA (20 nM) and AKE (1, 5, 10μg/ml) for 24 hr. Relative cell proliferation was determined using EZ-Cytox cell viability assay kit. Data represent the means±S.D. of 4 wells. *p<0.05, compared to control group.
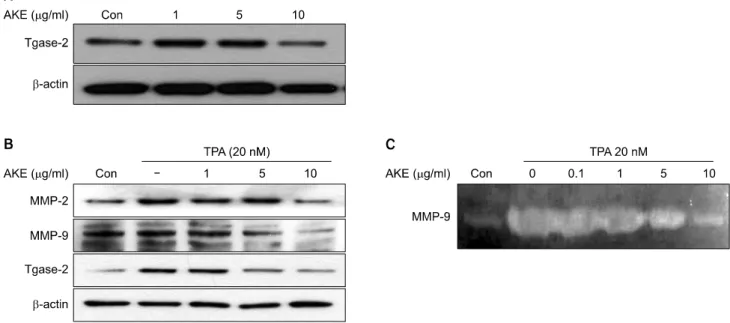
Fig. 3. Effects of AKE on expression of tgase-2, MMP-2 and MMP-9 in HT-1080 cells. (A) Effects of AKE on expression of tgase-2 in HT-1080 cells. HT-1080 cells were cultured in RPMI media. The cells were then treated with AKE (1, 5, 10μg/ml) for 24 hr.
Whole-cell lysates (20μg) were prepared, was subjected to 4∼12% SDS-PAGE and expression of several proteins were determined by western blotting. (B) Effects of AKE on expression of tgase-2, MMP-2, and MMP-9 in HT-1080 cells with TPA treatment. HT-1080 cells were cultured in RPMI media. The cells were then treated with TPA (20 nM) and AKE (1, 5, 10μg/ml) for 24 hr. Whole-cell lysates (20μg) were prepared and protein was subjected to 4∼12% SDS- PAGE, then expression of several proteins were determined by western blotting. (C) Zymographic analysis of AKE’s effects on MMP activity of TPA-treated HT-1080 cells. Zymography of HT-1080 cells treated with AKE. HT-1080 cells were treated with AKE (0.1, 1, 5, 10μg/ml) for 24 hr in the absence or presence of TPA (20 nM). The culture medium was collected and then analyzed for gelatinolytic activity by zymography.
2. AKE suppressed the proliferation of HT-1080 cells
The effect of AKE on HT-1080 cells proliferation was de- termined by MTT assay after 4 or 24 hr of incubation and cell viability was measured by trypan blue exclusion test. In Fig. 2, no cytotoxic effects of AKE were observed at 1∼25μg/ml with a 3 hr pre-incubation of HT-1080 cells with AKE (Fig. 2A).
However, AKE just slightly inhibited the proliferation of HT-1080 cells at 5∼10μg/ml (Fig. 2B). AKE showed even more significant inhibitory effect on HT-1080 cells under TPA-treated conditions (Fig. 2C).
3. AKE suppresses the expression of MMP-2, MMP-9 and tgase-2 in HT-1080 cells
MMP-2 and MMP-9 play a critical role in tumor invasion and tgase-2 is also implicated in migration and invasion of many cancer cells including breast cancer and pancreatic can- cers.12) But the role of tgase-2 expression has not been reported
with respect to migration and invasion of HT-1080 cells.
Therefore, MMP-2, MMP-9 and tgase-2, were examined in HT-1080 cells by western blotting and MMP’s activities were also analyzed by a gelatin zymography assay.
Tgase-2 expression was observed in HT-1080 cells and AKE treatment decreased the expression of tgase-2 (Fig. 3A). Ex- pression of MMP-2, MMP-9 and tgase-2 were also examined under the same conditions of TPA-induced invasion of HT- 1080 cells. The treatment of HT-1080 cells with TPA resulted in increased expression of tgase-2, MMP-2 and MMP-9 (Fig.
3B). AKE dose-dependently suppressed expressions of tgase-2, MMP-2 and MMP-9 after TPA treatment (Fig. 3B). AKE also suppressed MMP-9 activity of HT-1080 cells in the con- ditioned medium (Fig. 3C).
DISCUSSION AND CONCLUSION
In this study we showed that AKE inhibited the migration and TPA-induced invasion of HT-1080 cells by suppressing the
expressions of tgase-2, MMP-2 and MMP-9 as well as inhibit- ing the proliferation of HT-1080 cells.
Recently, targeted cancer therapy in the clinic has prolonged the lives of patients, but at high cost. Some cancers excised by surgery often recur in patients with metastases. In partic- ular, many complex events are involved in the recurrence can- cer and metastasis and it is very difficult to control these issues by targeted therapy by drugs. In these cases, natural products can be alternative ways to control metastasis/recurrence since natural products are composed of many compounds that may have anti-cancer or anti-metastatic properties.
We became interested in AK after finding that it has COX-2-inhibitory activity and shows topical anti-inflammatory and anti-pruritic activities in Thromboxane receptor-mediated scratching models.15,16) AK was also selected based on its tradi- tional use in Chinese medicine. AK is a member of the Zingi- beraceae family of plants. Previous investigations on AK have reported a variety of chalcones, flavonoids, diarylheptanoids, monoterpenes, sesquiterpenoids, and labdanes.13)
We examined whether AKE inhibited migration and TPA- induced invasion by HT-1080, a cell line commonly used in the study of cancer cell migration and invasion.20,21) In the re- sult, AKE dose-dependently suppressed migration and TPA-in- duced invasion of HT-1080 cells (Fig. 1). It is not known which compounds of AKE contribute to the anti-migratory and anti-invasive effects. Anti-invasive effects of AKE were due to decreased expressions of MMP-2 and MMP-9 (Fig. 3). But AKE also showed anti-proliferative effects on HT-1080 cells (Fig. 2). Thus, anti-proliferative effects also contributed the an- ti-invasive activity of AKE. It is quite interesting that the an- ti-proliferative effects of AKE are even stronger in TPA-treated HT-1080 cells (Fig. 2). This observation suggested AKE may be useful in controlling proliferative cancer cells. It is not clear which components of AKE contribute to the anti-proliferative activity, but cardamonin and pinocembrin from AK might be candidates since these compounds showed cytotoxicity to other cancer cells.22)
MMP-2, MMP-9 and tgase-2 expressions were suppressed by AKE (Fig. 3). These observations might be related to the fact that AK suppresses NF-ĸB as cardamonin from Alpinia spp.
can lower NF-ĸB activity. Therefore, cardamonin might con- tribute to the effects of AKE on expressions of MMP-2, MMP-9 and tgase-2. Further studies are ongoing to determine the role of cardamonin in MMP-2, MMP-9 and tgase-2
expressions.
By western blot analysis, expressions of MMP-2 and MMP-9 were suppressed by AKE but only decreased activity of MMP-9 was confirmed by gelatin zymography (Fig. 3B, C). The dis- crepancy between results of western blot and zymography can- not be explained presently, but it might be related to differ- ential expression of tissue inhibitor of metalloproteinases.23) In sum, AKE showed anti-migratory and anti-invasive effects on HT-1080 cells by suppressing the expressions of MMP-2, MMP-9 and tgase-2. These results suggested that AKE could be one of the new natural products useful in controlling the metastasis of cancers including fibrosarcomas.
ACKNOWLEDGEMENT
This study was supported by grants from the Research Pro- gram for New Drug Target Discovery Grant (2011-0030173), from the Ministry of Education, Science & Technology, Korea (2011-0015839) and from the the GRRC program of Gyeonggi province (GRRC Dongguk2011-B01).
REFERENCES
1) Ichikawa H, Nakamura Y, Kashiwada Y, Aggarwal BB. Anti- cancer drugs designed by mother nature: ancient drugs but modern targets. Curr Pharm Des 13, 3400-3416, 2007.
2) Langley RR, Fidler IJ. Tumor cell-organ microenvironment interactions in the pathogenesis of cancer metastasis. Endocr Rev 28, 297-321, 2007.
3) Mendoza M, Khanna C. Revisiting the seed and soil in cancer metastasis. Int J Biochem Cell Biol 41, 1452-1462, 2009.
4) Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 144, 646-674, 2011.
5) Egeblad M, Werb Z. New functions for the matrix metal- loproteinases in cancer progression. Nat Rev Cancer 2, 161- 174, 2002.
6) Roy R, Yang J, Moses MA. Matrix metalloproteinases as novel biomarkers and potential therapeutic targets in human cancer.
J Clin Oncol 27, 5287-5297, 2009.
7) Gialeli C, Theocharis AD, Karamanos NK. Roles of matrix metalloproteinases in cancer progression and their pharma- cological targeting. FEBS J 278, 16-27, 2011.
8) Lee CH, Kim SY. NF-ĸB and therapeeutic approach. Biomol
& Ther 17, 219-240, 2009.
9) Kim DS, Park SS, Nam BH, Kim IH, Kim SY. Reversal of drug resistance in breast cancer cells by transglutaminase 2 inhibition and nuclear factor-kappaB inactivation. Cancer Res 66, 10936-10943, 2006.
10) Mehta K, Fok JY, Mangala LS. Tissue transglutaminase: from
biological glue to cell survival cues. Front Biosci 11, 173-185, 2006.
11) Antonyak MA, Miller AM, Jansen JM, Boehm JE, Balkman CE, Wakshlag JJ, Page RL, Cerione RA. Augmentation of tissue transglutaminase expression and activation by epidermal growth factor inhibit doxorubicin-induced apoptosis in human breast cancer cells. J Biol Chem 279, 41461-41467, 2004.
12) Mangala LS, Fok JY, Zorrilla-Calancha IR, Verma A, Mehta K. Tissue transglutaminase expression promotes cell attach- ment, invasion and survival in breast cancer cells. Oncogene 26, 2459-2470, 2007.
13) Lee SE, Shin HT, Hwang HJ, Kim JH. Antioxidant activity of extracts from Alpinia katsumadai seed. Phytother Res 17, 1041-1047, 2003.
14) Yang Y, Kinoshita K, Koyama K, Takahashi K, Tai T, Nu- noura Y, Watanabe K. Two novel anti-emetic principles of Alpinia katsumadai. J Nat Prod 62, 1672-1674, 1999.
15) Noh MS, Ha JY, Lee CH, Lee WY, Lee SH, Lee JJ. Inhibitory activities of natural products on lipopolysaccharide induced prostaglandin production in mouse macrophages. Yakhakhoe Chi 42, 558-566, 1998.
16) Choi JK, Kim KM, Kim DK, Yeom MH, Koh JY, Jung SJ, Kim HJ, Oh SH, Kim SY, Lee CH. Topical anti-inflammatory and antipruritic effects of Alpinia katsumadai extracts. J Dermatol Sci 53, 81-84, 2009.
17) Choi JK, Kim KM, Yeom MH, Cho HY, Lee HJ, Park MK, Jeong KC, Lee BI, Noh MS, Lee CH. Antinociceptive effects
of Alpinia katsumadai via cyclooxygenase-2 inhibition. Biomol Ther 18, 159-165, 2010.
18) Kim JA, Kang Y, Thapa D, Lee JS, Park MA, Lee KH, Lyoo WS, Lee YR. Anti-invasive and anti-angiogenic effects of xanthohumol and its synthetic derivatives. Biomol Ther 17, 422-429, 2009.
19) Koh MS, Hwang JS, Moon A. Lycopene inhibits proliferation, invasion and migration of human breast cancer cells. Biomol Ther 18, 92-98, 2010.
20) Rajapakse N, Mendis E, Kim MM, Kim SK. Sulfated gluco- samine inhibits MMP-2 and MMP-9 expressions in human fibrosarcoma cells. Bioorg Med Chem 15, 4891-4896, 2007.
21) Sato T, Koike L, Miyata Y, Hirata M, Mimaki Y, Sashida Y, Yano M, Ito A. Inhibition of activator protein-1 binding activity and phosphatidylinositol 3-kinase pathway by nobile- tin, a polymethoxy flavonoid, results in augmentation of tissue inhibitor of metalloproteinases-1 production and suppression of production of matrix metalloproteinases-1 and -9 in human fibrosarcoma HT-1080 cells. Cancer Res 62, 1025-1029, 2002.
22) Tang J, Li N, Dai H, Wang K. Chemical constituents from seeds of Alpinia katsumadai, inhibition on NF-kappaB acti- vation and anti-tumor effect. Zhongguo Zhong Yao Za Zhi 35, 1710-1714, 2010.
23) Yan L, Lin B, Gao L, Gao S, Liu C, Wang C, Wang Y, Zhang S, Iwamori M. Lewis (y) antigen overexpression increases the expression of MMP-2 and MMP-9 and invasion of human ovarian cancer cells. Int J Mol Sci 11, 4441-4452, 2010.