2021, 32
(2)
,351–360
연속 시간 Markov chain 모형을 이용한 황소개구리 성장단계 전이확률 추정
†배
ᆨ자영1·강희진2·최일수3
13전남대학교 수학/통계학과 ·2전남대학교 생물과학생명기술학과
ᄌ ᅥ
ᆸᄉ ᅮ 2021ᄂ ᅧ ᆫ 2ᄋ ᅯ ᆯ 4ᄋ ᅵ ᆯ, ᄉ ᅮᄌ ᅥ ᆼ 2021ᄂ ᅧ ᆫ 3ᄋ ᅯ ᆯ 2ᄋ ᅵ ᆯ, ᄀ ᅦᄌ ᅢ ᄒ ᅪ ᆨᄌ ᅥ ᆼ 2021ᄂ ᅧ ᆫ 3ᄋ ᅯ ᆯ 4ᄋ ᅵ ᆯ
요 약
ᄋ ᅣ
ᆼᄉ ᅥᄅ ᅲᄂ ᅳ ᆫ ᄒ ᅪ ᆫᄀ ᅧ ᆼ ᄇ ᅧ ᆫᄒ ᅪᄋ ᅦ ᄆ ᅵ ᆫᄀ ᅡ ᆷᄒ ᅡᄋ ᅧ ᄌ ᅥ ᆫᄉ ᅦᄀ ᅨᄌ ᅥ ᆨᄋ ᅳᄅ ᅩ ᄉ ᅵ ᆷᄀ ᅡ ᆨᄒ ᅡ ᆫ ᄀ ᅡ ᆷᄉ ᅩᄎ ᅮᄉ ᅦᄅ ᅳ ᆯ ᄇ ᅩᄋ ᅵᄀ ᅩ ᄋ ᅵ ᆻᄃ ᅡ. ᄋ ᅣ ᆼᄉ ᅥᄅ ᅲᄂ ᅳ ᆫ ᄒ ᅪ ᆫᄀ ᅧ ᆼᄇ ᅧ ᆫ ᄒ
ᅪᄋ ᅦ ᄆ ᅵ ᆫᄀ ᅡ ᆷᄒ ᅡ ᆫ ᄌ ᅵᄑ ᅭᄌ ᅩ ᆼ ᄋ ᅳᄅ ᅩ ᄋ ᅡ ᆯᄅ ᅧᄌ ᅧᄋ ᅵ ᆻᄃ ᅡ. ᄋ ᅮᄅ ᅵᄂ ᅡᄅ ᅡᄋ ᅦ ᄋ ᅲᄋ ᅵ ᆸᄃ ᅬᄋ ᅥ ᄌ ᅥ ᆫᄀ ᅮ ᆨᄌ ᅥ ᆨᄋ ᅵ ᆫ ᄒ ᅪ ᆨ ᄉ ᅡ ᆫᄋ ᅳ ᆯ ᄇ ᅩᄋ ᅵᄂ ᅳ ᆫ ᄒ ᅪ ᆼᄉ ᅩᄀ ᅢᄀ ᅮᄅ ᅵᄂ ᅳ ᆫ ᄒ ᅪ ᆫ ᄀ ᅧ
ᆼᄇ ᅧ ᆫᄒ ᅪᄋ ᅦ ᄆ ᅵ ᆫᄀ ᅡ ᆷᄒ ᅡ ᆫ ᄋ ᅣ ᆼᄉ ᅥᄅ ᅲᄋ ᅵᄌ ᅡ ᄀ ᅪ ᆫ ᄅ ᅵᄇ ᅡ ᆼᄋ ᅡ ᆫᄋ ᅵ ᄑ ᅵ ᆯᄋ ᅭᄒ ᅡ ᆫ ᄋ ᅬᄅ ᅢᄉ ᅢ ᆼᄆ ᅮ ᆯ ᄅ ᅩᄉ ᅥ, ᄆ ᅢᄋ ᅮ ᄌ ᅮ ᆼ ᄋ ᅭᄒ ᅡ ᆫ ᄋ ᅧ ᆫᄀ ᅮᄌ ᅥ ᆨ ᄀ ᅡᄎ ᅵᄅ ᅳ ᆯ ᄌ ᅵᄂ ᅵ ᆫᄃ ᅡ.
ᄒ
ᅧ ᆫᄉ ᅵ ᆯᄌ ᅥ ᆨᄋ ᅳᄅ ᅩ ᄒ ᅪ ᆼᄉ ᅩᄀ ᅢᄀ ᅮᄅ ᅵᄋ ᅦ ᄃ ᅢᄒ ᅡ ᆫ ᄌ ᅩᄉ ᅡᄂ ᅳ ᆫ ᄉ ᅵᄀ ᅡ ᆫᄀ ᅪ ᄇ ᅵᄋ ᅭ ᆼ ᄋ ᅴ ᄆ ᅮ ᆫ ᄌ ᅦᄅ ᅩ ᄆ ᅡ ᆭᄋ ᅳ ᆫ ᄌ ᅦᄋ ᅣ ᆨᄋ ᅳ ᆯ ᄀ ᅡ ᆽᄂ ᅳ ᆫ ᄃ ᅡ. ᄇ ᅩ ᆫ ᄂ ᅩ ᆫᄆ ᅮ ᆫ ᄋ ᅦᄉ ᅥᄂ ᅳ ᆫ ᄋ ᅧ ᄅ
ᅥ ᄂ ᅩ ᆫᄆ ᅮ ᆫ ᄋ ᅦᄉ ᅥ ᄌ ᅦᄉ ᅵᄒ ᅡᄂ ᅳ ᆫ ᄒ ᅪ ᆼᄉ ᅩᄀ ᅢᄀ ᅮᄅ ᅵᄋ ᅦ ᄃ ᅢᄒ ᅡ ᆫ ᄉ ᅢ ᆼᄐ ᅢᄌ ᅥ ᆨᄋ ᅵ ᆫ ᄐ ᅳ ᆨᄌ ᅵ ᆼᄋ ᅳᄅ ᅩ ᄒ ᅪ ᆼᄉ ᅩᄀ ᅢᄀ ᅮᄅ ᅵᄋ ᅴ ᄇ ᅧ ᆫᄋ ᅵᄒ ᅢ ᆼᄐ ᅢᄅ ᅳ ᆯ ᄎ ᅮᄌ ᅥ ᆼᄒ ᅢ ᆻᄃ ᅡ. ᄉ ᅵ ᄀ
ᅡ ᆫᄋ ᅴ ᄒ ᅳᄅ ᅳ ᆷ ᄋ ᅦ ᄄ ᅡᄅ ᅳ ᆫ ᄉ ᅥ ᆼᄌ ᅡ ᆼᄃ ᅡ ᆫᄀ ᅨᄅ ᅳ ᆯ ᄋ ᅧ ᆫᄉ ᅩ ᆨ ᄉ ᅵᄀ ᅡ ᆫ Markov chain ᄆ ᅩᄃ ᅦ ᆯᄋ ᅦ ᄌ ᅥ ᆨᄋ ᅭ ᆼ ᄒ ᅡᄋ ᅧ ᆻᄋ ᅳᄆ ᅧ ᄌ ᅥ ᆫᄋ ᅵ ᄒ ᅪ ᆨᄅ ᅲ ᆯ ᄀ ᅪ ᄑ ᅧ ᆼᄀ ᅲ ᆫ ᄉ ᅢ ᆼᄌ ᅩ ᆫ ᄀ
ᅵᄀ ᅡ ᆫᄋ ᅳ ᆯ ᄎ ᅮᄌ ᅥ ᆼᄒ ᅡᄋ ᅧ ᆻᄃ ᅡ. ᄐ ᅳ ᆨ ᄒ ᅵ ᄀ ᅡ ᆨ ᄃ ᅡ ᆫᄀ ᅨᄇ ᅧ ᆯ ᄉ ᅡᄆ ᅡ ᆼᄁ ᅡᄌ ᅵ ᄋ ᅵᄅ ᅳᄂ ᅳ ᆫ ᄑ ᅧ ᆼᄀ ᅲ ᆫ ᄉ ᅢ ᆼᄌ ᅩ ᆫ ᄀ ᅵᄀ ᅡ ᆫᄋ ᅳ ᆫ ᄋ ᅡ ᆯᄋ ᅦᄉ ᅥᄂ ᅳ ᆫ ᄑ ᅧ ᆼᄀ ᅲ ᆫ 104.825ᄋ ᅵ ᆯ, ᄋ ᅩ ᆯ ᄎ ᅢ
ᆼᄋ ᅵᄋ ᅦᄉ ᅥᄂ ᅳ ᆫ 520.125ᄋ ᅵ ᆯ, ᄉ ᅥ ᆼᄎ ᅦᄋ ᅦᄉ ᅥᄂ ᅳ ᆫ 2,920.0ᄋ ᅵ ᆯᄋ ᅵ ᄀ ᅥ ᆯᄅ ᅵᄂ ᅳ ᆫ ᄀ ᅥ ᆺᄋ ᅳᄅ ᅩ ᄂ ᅡᄐ ᅡᄂ ᅡ ᆻᄃ ᅡ.
ᄌ
ᅮᄋ ᅭᄋ ᅭ ᆼ ᄋ ᅥ: ᄋ ᅧ ᆫᄉ ᅩ ᆨ ᄉ ᅵᄀ ᅡ ᆫ Markov chain ᄆ ᅩᄃ ᅦ ᆯ, ᄋ ᅧ ᆫᄉ ᅩ ᆨᄌ ᅥ ᆫᄋ ᅵ ᄒ ᅪ ᆨᄅ ᅲ ᆯ, ᄌ ᅥ ᆫᄋ ᅵᄅ ᅲ ᆯ ᄒ ᅢ ᆼᄅ ᅧ ᆯ, ᄒ ᅪ ᆼᄉ ᅩᄀ ᅢᄀ ᅮᄅ ᅵ.
1. 서론 ᄋ
ᅣᆼ서류는 분류학적으로 척삭동물문 (Phylum Chordata) 양서강 (Class Amphibia)으로 분류되고, ᄆ
ᅮ족영원목 (Order Gymnophiona),개구리목 (Order Anura),도롱뇽목 (Order Caudata)으로 나누어 ᄌ
ᅵᆫ다 (고영민, 2011). 양서류는생태계 먹이사슬에서 중간자 위치로, 상위포식자에게는먹이자원이 되 ᄀ
ᅩ 곤충류를포식하여 먹이사슬에서 중요한 역할을수행한다 (Kim 과 Song, 2010). 하지만, 생활사 측 ᄆ
ᅧᆫ에서환경 변화에 민감한 특징을가지고 있으며 (An 등, 2020),기후 변화, 질병, 서식지 손실 등의 원 ᄋ
ᅵᆫ으로 전세계 양서류의 약 1/3이 심각한 감소를보이고 있다 (Stuart 등, 2004).
ᄋ
ᅣᆼ서류는변온척추동물로서, 성장 (Growth)과 발달 (Development)을 통해 알 (egg), 유생 (larvae), ᄋ
ᅡ성체 (juvenile), 성체 (adult)를거치는생활사를갖는다 (Vonesh 와 De la Cruz, 2002). 양서류 중 ᄀ
ᅢ구리목 (무미목), 도롱뇽목 (유미목)양서류는 수생생활을 하는 유생 (larvae)에서 변태 (metamor- phosis)과정을거쳐서 육상생활이 가능한 작은성체가된다 (Alford와 Harris, 1988; Hota, 1994). 양 ᄉ
ᅥ류는생활사 전반에 걸쳐 외부환경에 민감한 영향을받으므로 (Trochet 등, 2014),양서류의 생활사 르
ᆯ파악하고 예측하는것은매우 중요하다. 특히 양서류의 발달에 온도는 중요한 요인이 되며, 낮은 온
†
ᄇ ᅩ ᆫ ᄀ ᅧ ᆯᄀ ᅪᄆ ᅮ ᆯᄋ ᅳ ᆫ ᄒ ᅪ ᆫᄀ ᅧ ᆼᄇ ᅮᄋ ᅴ ᄌ ᅢᄋ ᅯ ᆫ ᄋ ᅳᄅ ᅩ ᄒ ᅡ ᆫᄀ ᅮ ᆨᄒ ᅪ ᆫᄀ ᅧ ᆼᄉ ᅡ ᆫᄋ ᅥ ᆸᄀ ᅵᄉ ᅮ ᆯᄋ ᅯ ᆫ ᄋ ᅴ ᄉ ᅢ ᆼᄆ ᅮ ᆯ ᄃ ᅡᄋ ᅣ ᆼᄉ ᅥ ᆼ ᄋ ᅱᄒ ᅧ ᆸ ᄋ ᅬᄅ ᅢᄉ ᅢ ᆼᄆ ᅮ ᆯ ᄀ ᅪ ᆫ ᄅ ᅵ ᄀ ᅵᄉ ᅮ ᆯ ᄀ ᅢᄇ ᅡ ᆯᄉ ᅡᄋ ᅥ ᆸᄋ ᅴ ᄌ
ᅵᄋ ᅯ ᆫᄋ ᅳ ᆯ ᄇ ᅡ ᆮᄋ ᅡ ᄋ ᅧ ᆫᄀ ᅮᄃ ᅬᄋ ᅥ ᆻᄉ ᅳ ᆸ ᄂ ᅵᄃ ᅡ (RE201807039).
1
(61186) ᄀ ᅪ ᆼ ᄌ ᅮ ᄀ ᅪ ᆼᄋ ᅧ ᆨᄉ ᅵ ᄇ ᅮ ᆨ ᄀ ᅮ ᄋ ᅭ ᆼᄇ ᅩ ᆼ ᄅ ᅩ 77, ᄌ ᅥ ᆫᄂ ᅡ ᆷᄃ ᅢᄒ ᅡ ᆨᄀ ᅭ ᄐ ᅩ ᆼ ᄀ ᅨᄒ ᅡ ᆨᄀ ᅪ, ᄇ ᅡ ᆨᄉ ᅡᄀ ᅪᄌ ᅥ ᆼ.
2
(61186) ᄀ ᅪ ᆼ ᄌ ᅮ ᄀ ᅪ ᆼᄋ ᅧ ᆨᄉ ᅵ ᄇ ᅮ ᆨ ᄀ ᅮ ᄋ ᅭ ᆼᄇ ᅩ ᆼ ᄅ ᅩ 77, ᄌ ᅥ ᆫᄂ ᅡ ᆷᄃ ᅢᄒ ᅡ ᆨᄀ ᅭ ᄉ ᅢ ᆼᄆ ᅮ ᆯ ᄒ ᅡ ᆨᄀ ᅪ, ᄇ ᅡ ᆨᄉ ᅡᄀ ᅪᄌ ᅥ ᆼ.
3
ᄀ ᅭᄉ ᅵ ᆫᄌ ᅥᄌ ᅡ: (61186) ᄀ ᅪ ᆼ ᄌ ᅮ ᄀ ᅪ ᆼᄋ ᅧ ᆨᄉ ᅵ ᄇ ᅮ ᆨ ᄀ ᅮ ᄋ ᅭ ᆼᄇ ᅩ ᆼ ᄅ ᅩ 77, ᄌ ᅥ ᆫᄂ ᅡ ᆷᄃ ᅢᄒ ᅡ ᆨᄀ ᅭ ᄐ ᅩ ᆼ ᄀ ᅨᄒ ᅡ ᆨᄀ ᅪ, ᄀ ᅭᄉ ᅮ.
E-mail: [email protected]
ᄃ
ᅩ는양서류의 성장과 발달을억제시킨다. 대개 온도가 증가함에 따라 발달 기간이 짧아지는것으로 나 ᄐ
ᅡ난다 (Herreid 와 Kinney, 1967; Vignoli 등, 2007).
ᄋ
ᅧᆫ속시간 Markov chain (Continuous time Markov chain, CTMC)모델은연속된시간에서 발생하 느
ᆫ 사건의 상태 변화를나타내는모델로 사회 과학, 생태학, 의학 등 다양한 분야에서 적용되고 있다.
CTMC모델은연속된시간을고려하기 때문에 이산 시간 Markov chain (Discrete time Markov chain, DTMC)보다 현실적인 문제를해결하는데 유용하다. 또한 시간에 따른전이확률의 변화를 볼수 있다 느
ᆫ 장점이 있다. Kalbfleisch와 Lawless (1985)는 panel data에 CTMC 모델을 적용하고 최우추정값 으
ᆯ 구하는 효율적인 방법을 제시했다. 실제 청소년의 흡연 연구에서 교육을 통해 흡연 상태가 어떻게 ᄇ
ᅧᆫ화하는지 추적 조사한 데이터를이용해 CTMC 모델을 적용하였다 (Kalbfleisch와 Lawless, 1985).
Etterson 등 (2009)은조류의 계절별 번식률을 추정하기 위해 Markov chain 모델을 이용하여 암컷당 ᄇ
ᅥᆫ식 성공률을추정하였다. 의학 분야에서는 HIV감염으로의 진행 단계를 CTMC모델을이용하여 두 ᄀ
ᅡ지 마커 CD4 수치와 anti-CD3의 반응성에 따른결과를비교하였다. 두 마커 모두 HIV 감염으로 진 ᄒ
ᅢᆼ하며 위험을 높이는데 요인으로 관련이 있었다. 또한 CTMC 모델은 HIV 감염을 진단하는 마커를 ᄋ
ᅧᆫ구하는데 유용하다는결과를얻었다 (Hendriks 등, 1996). 최근연구로는간경화환자가 사망에 이르 느
ᆫ과정에서의 전이확률을 CTMC모델을이용하여 계산하였다. 그 결과 간경화환자가 간암으로 진행 ᄒ
ᅡᆯ확률은성별, 연령에 따라 다르지만 C형 간염바이러스 및 알코올남용과관련된질병에 영향을미치 ᄌ
ᅵ 않음을보였다 (Bartolomeo 등, 2011). 또한 혼합 분포 (mixture distribution)를사용한 모형들도 ᄉ
ᅡ용하는 등 응용범위를넓혀가고 있다 (Wilson 등 , 2018; Ko와 Choi, 2018; Ahn과 Kim, 2018).
보
ᆫ연구에서는 CTMC모델을이용하여황소개구리의 성장과정을모델링하고 생활사 각 단계에 해당 ᄒ
ᅡ는전이확률과 사망에 이르기 까지의 각 단계별 평균생존기간을추정했다.
2. 자료 요약 화
ᆼ소개구리 (American Bullfrog, Lithobates catesbeianus)는전세계에 가장 많이확산된양서류로서 (Lowe 등, 2000), 토착종의 번식 간섭, 항아리곰팡이 (Batrachochytrium dendrobatidis)의확산, 생태 ᄀ
ᅨ 교란 등의 문제를야기하고 있다 (Kraus, 2015; Gobel 등, 2019). 이러한 문제를 근거로 국제자연 ᄇ
ᅩ전연맹 (International Union for Conservation of Nature, IUCN)은황소개구리를 100대 악성 생태 ᄀ
ᅨ위해외래생물종으로 지정하였으며 (Lowe 등, 2000), 국내환경부에서는생태계교란 생물로 지정하여 과
ᆫ리하고 있다 (Ministry of Environment, 2020).
화
ᆼ소개구리의 성장 과정은알, 유생, 아성체, 성체의 네 단계로 나눌수 있다. 유생 단계는알에서 세 ᄑ
ᅩ 분열 과정을거쳐 도달하는단계로 15-32℃ 사이의 적정한 온도를유지하면 평균 3-5일 정도를거쳐 오
ᆯ챙이로 도달하는것으로 알려져 있다. 이 과정에서 기온이 32℃를넘는다면 기형으로 부화할 가능성 ᄋ
ᅵ 높아지고 35.9℃ 이상의 온도에서는부화하지 못하고 사망에 이르는것으로 알려져 있다 (Gosner와 Black, 1957; Howard, 1978). 아성체 단계는 올챙이에서 변태 과정을마쳤지만 아직 생식 능력이 없는 ᄉ
ᅡᆼ태로 번식이 불가능한 단계를말한다. 올챙이는 물속에서 평균 1-3년 정도의 시간을보내고 아성체로 바
ᆯ달한다. 이 과정에서도 따뜻한 지역이 추운지역에서 보다 빠른시간 내에 올챙이에서 아성체로 발달 ᄒ
ᅡᆷ을알 수 있었다 (Burry와 Whelan, 1985). 황소개구리의 수명은 8-10년 정도로 추정되며 사육시설 ᄎ
ᅥ럼 통제된 환경에서는 16년까지 생존한 경우도 발견되었다 (Durham, 1963). 성체가된이후에도 실 ᄒ
ᅥᆷ실과 같이 적절한환경으로 통제된 경우 오랜 시간 생존함을알 수 있다.
화
ᆼ소개구리의 성장 과정에서 온도는 중요한 요인이며 15℃ 이하의환경에서는다음성장 단계로 넘어 ᄀ
ᅡ지 못하거나 넘어가는데 많은시간을요구함을알 수 있다 (Viparina와 Just, 1975). 양서류 발달에 ᄂ
ᅡ
ᆽ은 온도가 성장과 발달을억제시킨다는 특징을황소개구리 역시 그대로 반영하고 있다 (Wheeler 등,
2015).
3. 연속시간 Markov chain 모형 화
ᆼ소개구리의 성장과정을알, 유생, 성체 (아성체 포함)으로 정의하고 CTMC 모델을적용하고자 한 ᄃ
ᅡ. CTMC 모델의 적용을위한황소개구리의 성장 과정에서 각 상태 (state) 간의 이동과정을 Figure 3.1에 나타내었다. 알, 유생에서는 아래와 같이 정의 된순간 전이률 (instantaneous transition rate) ᄋ
ᅳ로 다음발달 단계로 이동한다.
qij= lim
∆t→0
P (transition i to j in [t, t + ∆t]| state i in t)
∆t , i, j = 1, 2, 3 i ̸= j.
ᄄ
ᅩ한 알, 유생, 성체에서는아래와 같이 정의된 순간전이률 (instantaneous transition rate)로 사망단 ᄀ
ᅨ에 이르게된다.
µk= lim
∆t→0
P (transition k to Death in [t, t + ∆t]| state k in t)
∆t , k = 1, 2, 3.
ᄋ
ᅨ를 들어, q12는단위 시간당 알에서 유생으로 진행하는비율을나타내고 µ3는성체에서 사망하는비율 ᄋ
ᅳᆯ나타낸다.
Figure 3.1 Life cycle of bullfrog
CTMC모델에서 시간에 따른각 상태 (state) 간의 전이확률, P(t)은아래와 같이 계산된다.
P(t) = etQ=
∞
X
n=0
1 n!(tQ)n, ᄋ
ᅧ기서 Q는 순간 전이률 행렬 (instantaneous transition rate matrix, infinitesimal generator)로 다음 ᄀ
ᅪ 같이 정의된다.
Q =
−(q12+ µ1) q12 0 µ1
0 −(q23+ µ2) q23 µ2
0 0 −µ3 µ3
0 0 0 0
.
Q의 대각화를 통해 전이확률은다음과 같이 계산될수 있다.
P(t) = SetDS−1, ᄋ
ᅧ기서 D는 Q의 고유치 (eigenvalue)를 주대각 원소로 가지는 대각행렬이고 S는 Q의 고유벡터 (eigenvector)를 열벡터로 이루어진 행렬이다. 황소개구리 성장 과정에서 정의한 Q 행렬의 고유값 (eigenvalue)은 λ1 = −(q12+ µ1), λ2 = −(q23+ µ2), λ3 = −µ3, λ4 = 0이고 이에 해당하는고유벡터 (eigenvector)는다음과 같다.
1 0 0 0
,
1
q12+µ1−q23−µ2 q12
0 0
,
q12 q12+µ1−µ3
1
q23+µ2−µ3 q23
0
,
1 1 1 1
.
ᄄ
ᅡ라서황소개구리의 성장과정에서 시간에 따른각 상태 (state) 간의 전이확률, P(t)은다음과 같다.
P(t) = 1 AC
etλ1 C(etλ2− etλ1) etλ1− etλ2+ AB(etλ3− etλ1) C(1−etλ2)−AC(1−etλ1)
+(AB−1)(1−Betλ3)
0 ACetλ2 A(etλ3− etλ2) A(etλ2− etλ3) + AC(1 − etλ2) 0 0 ACetλ3 AC(1 − etλ3)
0 0 0 AC
,
ᄋ
ᅧ기서 A = (q12+ µ1− q23− µ2)/q12, B = q12/(q12+ µ1− µ3), C = (q23+ µ2− µ3)/q23이다.
보
ᆫ 연구에서 CTMC 모델을 이용한 황소개구리의 성장 과정은 흡수상태 (absorbing state)를 포함 ᄒ
ᅡ고 있다. 흡수상태 (absorbing state)는 이 단계로 상태 변화하면 다른단계로 이동할 수 없는 상태 (state)로황소개구리의 성장 모델에서는알, 유생 또는성체에서 사망에 이르면 다른단계로 이동할 수 어
ᆹ으므로 사망이 흡수상태 (absorbing state)라고 할 수 있다. 흡수상태 (absorbing state)를포함하는 ᄆ
ᅩ델에서는각 상태 (state) i에서 흡수상태 (absorbing state)까지 도달하는데 걸리는평균시간(ai)을 ᄋ
ᅡ래와 같이 계산할 수 있다 (Dobrow, 2016).
ai=X
j
Fij,
ᄋ
ᅧ기서 F 는 F = −V−1이고 V는 Q에서 다음과 같이 정의되는 행렬이다.
Q = V * 0 0
! .
화
ᆼ소개구리 성장 모델에서 F는다음과 같이 계산된다.
F = 1
µ3(q12+ µ1)(q23+ µ2)
(q23+ µ2)µ3 q12µ3 q12q23
0 (q12+ µ1)µ3 (q12+ µ1)q23
0 0 (q12+ µ1)(q23+ µ2)
.
4. 자료분석 결과 이
ᆯ반적으로 온도가 낮은지역에서는단계별 성장 과정의 길이가 길어지고 온도가 높은지역에서는짧 ᄋ
ᅡ지는과정을갖는다. 올챙이에서 개구리로 변태되는기간은루이지에나 (Louisiana)에서 평균 6개월
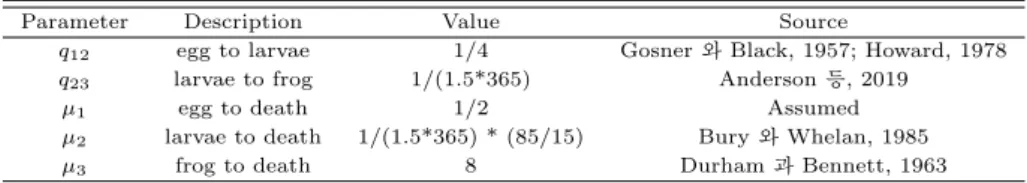
Table 4.1 Summary table of instantaneous rate
Parameter Description Value Source
q12 egg to larvae 1/4 Gosner와 Black, 1957; Howard, 1978 q23 larvae to frog 1/(1.5*365) Anderson 등, 2019
µ1 egg to death 1/2 Assumed
µ2 larvae to death 1/(1.5*365) * (85/15) Bury와 Whelan, 1985
µ3 frog to death 8 Durham과 Bennett, 1963
ᄋ
ᅵ하인 반면 캐나다 노바 스코티아 (Nova scotia)에서는평균 3년이 걸리는것으로 알려져 있다. Table 4.1 은최적 성장온도인 온대지방을 기준으로한 온도에서황소개구리 성장단계에서 전이율을나타내는 ᄆ
ᅩ수값을나타내고 이에 따른 순간 전이률 행렬 (instantaneous transition rate matrix, infinitesimal generator), Q는다음과 같이 계산된다.
Q =
−0.75 0.25 0 0.5 0 −0.0121 0.0018 0.0103 0 0 −0.0003 0.0003
0 0 0 0
= V * 0 0
! .
Q의 고유치 (eigenvalues)는 0.75, 0.0121, 0.0003, 0이고 이에따른 CTMC모델에서 시간에 따른황소 ᄀ
ᅢ구리의 성장단계의 전이확률은다음과 같이 계산된다.
Pij(t) =
e−0.75t
19.1219, i = 1, j = 1.
e−0.0121t−e−0.75t
2.9512 , i = 1, j = 2.
e−0.75t−e−0.0121t
19.1219 + 0.0514(e0.0003t− e−0.75t), i = 1, j = 3.
1−e−0.0121t
2.9512 − (1 − e−0.75t) +−0.0157(1−0.3334e−0.0003t)
19.1219 , i = 1, j = 4.
e−0.0121t, i = 2, j = 2.
e−0.0003t−e−0.0121t
6.4791 , i = 2, j = 3.
e−0.0003t−e−0.0121t
6.4791 + (1 − e−0.0121t), i = 2, j = 4.
e−0.0127t, i = 3, j = 3.
1 − e−0.0127t, i = 3, j = 4.
1, i = 4, j = 4.
0, otherwise.
(4.1)
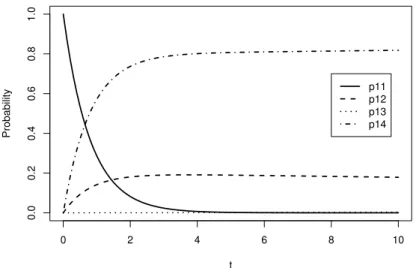
Figure 4.1부터 Figure 4.4는각 단계로 옮겨가는전이확률 식 (4.1)를도식화하였다. Figure 4.1은 ᄎ
ᅩ기 10일 동안에 알에서 이동하는전이확률을나타내고 있고 이에 따르면 초기에 사망확률 P14이 급격 ᄒ
ᅡ게 증가하고 올챙이로 전이할확률 P12는 2일경에 20%에 수렴하게 된다. 또한 알에서 머무를 확률 P11은초기에 급격하게 감소하여 4일 이후가 되면 거의 0에 가까운작은값을가진다. 알에서 개구리로 ᄇ
ᅧᆫ태될 확률 P13은초기에 점점 증가하다 304일경에 2.705%로 가장큰값을가지고 그 후에 시간이 지 ᄂ
ᅡ
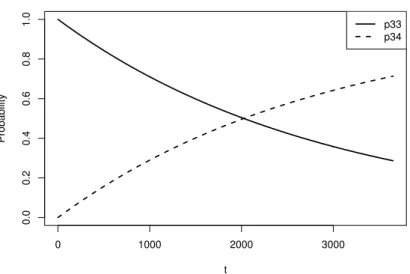
ᆷ에 따라 감소하였다. Figure 4.3은 올챙이에서 이동하는전이확률을나타내었다. 올챙이에서 성체로 ᄇ
ᅧᆫ이할확률 P23은 303일째까지 증가하여 13.527%로 가장큰값을가지고 그 이후에는점점 감소하고 이
ᆻ다. 성제가된후 사망할확률 P34는시간이 지남에 따라 점점 증가함을알 수 있다.
0 2 4 6 8 10
0.00.20.40.60.81.0
t
Probability
p11 p12 p13 p14
Figure 4.1 Transition probability plot from egg (early period)
0 1000 2000 3000
0.00.20.40.60.81.0
t
Probability
p12 p13 p14
Figure 4.2 Transition probability plot from egg (whole period)
ᄄ
ᅩ한 F = −V−1이므로 F는다음과 같다.
F =
0.8 16.425 87.6 0.0 82.125 438.0 0.0 0.0 2920.0
.
화
ᆼ소개구리의 각 성장단계에서 사망에 이르기까지 걸리는평균시간은알에서 올챙이까지 0.8일, 올 채
ᆼ이에서 성체까지 16.425일, 성체에서 87.6일로써 알에서 사망에 이르는 평균시간은 104.825일이고, 오
ᆯ챙이에서는 520.125일, 성체에서는 2,920.0일이 걸리는것으로 나타났다.
0 1000 2000 3000
0.00.20.40.60.81.0
t
Probability
p22 p23 p24
Figure 4.3 Transition probability plot from larvae
0 1000 2000 3000
0.00.20.40.60.81.0
t
Probability
p33 p34
Figure 4.4 Transition probability plot from adult
5. 결론 새
ᆼ태계 교란 야생생물로 분류되는황소개구리에 대한 생태학적인 조사 및 표본수집은 방대한 시간과 ᄂ
ᅩ력이 투입되는과정을거쳐야 한다. 본 논문에서는이러한 과정에서 얻을수 있는성장단계에서의 변 ᄋ
ᅵ확률과 사망에 이르기까지의 평균시간을수리적인 모형으로 구할 수 있는방법을 제시하였다. 이에 ᄄ
ᅡ르면 알에서 올챙이로 가는 초기확률이 현저히 작으며 성체가 되면 안정적으로 사망에 이르는 확률 ᄋ
ᅵ 작아져 생존율을유지하고 있음을알 수 있다. 사망에 이르는평균생존시간은 알은 104일, 올챙이
느
ᆫ 520.125일과 성체는 2,920.0일을나타냈다. 이는황소개구리의 초기사망율이 높은것을설명하고 있 ᄃ
ᅡ. 모형의 모수들에 대한 온도에 따른전이율이 달라진다 것은알려진 사실이다 (Na 등, 2015). 따라 ᄉ
ᅥ 생존온도에 따른전이확률을구하고 생존확률에 대응되는개체군의 변화를예측하는데 본 논문의 결 ᄀ
ᅪ는 유용할 것으로 생각된다. 특히 각 성장단계의 기간과 온도함수의 상관관계를 규명하는함수를 구 ᄒ
ᅡ여 미래의 기후변화에 대한황소개구리의 개체군변화를예측하는데에도 기여할 수 있을것으로 기대 되
ᆫ다.
References
Ahn, C. and Kim, D. (2018). Variable selection based on semi-parametric estimator of conditional mu- tual information assuming normal mixture in high-dimensional data. Journal of the Korean Data &
Information Science Society, 29, 1339-1351.
Alford, R. A. and Harris, R. N. (1988). Effects of larval growth history on anuran metamorphosis. The American Naturalist, 131, 91-106.
An, C. K., Hong, S. G., Na, S., Doh, J., Oh, K. C. and Yi, H. (2020). Comparison Study of the Snout- vent Length (SVL) and the biomass for the climate change sensitive species, Narrow-mouthed Toads (Kaloula borealis, Endangered Species II), at the three different areas (Seoul, Nonsan, Busan) of South Korea. Journal of Wetlands Research, 22, 8-14.
Anderson, R. B., Rose, J. P. and Lawler, S. P. (2019). Evolutionary experience with the invasive Lithobates catesbeianus predicts lower survival of larval rana draytonii. Herpetological Conservation and Biology, 14, 349-359.
Bartolomeo, N., Trerotoli, P. and Serio, G. (2011). Progression of liver cirrhosis to HCC: an application of hidden Markov model. BMC Medical Research Methodology, 11, 1-8.
Durham, L. and Bennett, G. W. (1963). Age, growth, and homing in the bullfrog. The Journal of Wildlife Management, 107-123.
Etterson, M. A., Bennett, R. S., Kershner, E. L. and Walk, J. W. (2009). Markov chain estimation of avian seasonal fecundity. Ecological Applications, 19, 622-630.
Gobel, N., Laufer, G. and Cortizas, S. (2019). Changes in aquatic communities recently invaded by a top predator: evidence of American bullfrogs in Acegua, Uruguay. Aquatic Sciences, 81, 8.
Gosner, K. L. and Black, I. H. (1957). The effects of acidity on the development and hatching of New Jersey frogs. Ecology, 38, 256-262.
Howard, R. D. (1978). The evolution of mating strategies in bullfrogs, Rana catesbeiana. Evolution, 850- 871.
Hendriks, J. C., Satten, G. A., Longini, I. M., Van Druten, H. A., Schellekens, P. T. A., Coutinho, R. A. and Van Griensven, G. J. (1996). Use of immunological markers and continuous-time Markov models to estimate progression of HIV infection in homosexual men. Aids-London-Current Science Then Rapid Science Publishers Then Lippincott Raven-, 10, 649-656.
Hota, A. K. (1994). Growth in amphibians. Gerontology, 40, 147-160.
Kalbfleisch, J. D. and Lawless, J. F. (1985). The analysis of panel data under a Markov assumption. Journal of the American Statistical Association, 80, 863-871.
Ko, K. and Choi, I. (2018). Incubation ratio estimation for Plasmodium vivax malaria in Korea using truncated mixture model. Journal of the Korean Data & Information Science Society, 29, 1025-1035.
Kraus, F. (2015). Impacts from invasive reptiles and amphibians. Annual Review of Ecology, Evolution, and Systematics, 46, 75-97.
Kim, J. B. and Song, J. Y. (2010). Amphibians and reptiles in Korea, World Science Publishing, Seoul.
Na, S., Shim, J. E., Kim, H. J., An, C. K. and Yi, H. (2015). The effect of three different water temperatures in our research facility on Huanren brown frog (Rana huanrensis) egg’s hatching rate, hatching periods, and larvae’s growth. Journal of Wetlands Research, 17, 320-324.
Stuart, S. N., Chanson, J. S., Cox, N. A., Young, B. E., Rodrigues, A. S., Fischman, D. L. and Waller, R. W.
(2004). Status and trends of amphibian declines and extinctions worldwide. Science, 306, 1783-1786.
Trochet, A., Moulherat, S., Calvez, O., Stevens, V. M., Clobert, J. and Schmeller, D. S. (2014). A database of life-history traits of European amphibians. Biodiversity Data Journal.
Viparina, S. and Just, J. J. (1975). The life period, growth and differentiation of Rana catesbeiana larvae
occurring in nature. Copeia, 103-109.
Vonesh, J. R. and Dela Cruz, O. (2002). Complex life cycles and density dependence: assessing the contri- bution of egg mortality to amphibian declines. Oecologia, 133, 325-333.
Bury, R. B. and Whelan, J. A. (1985). Ecology and management of the bullfrog, US Department of the Interior, Fish and Wildlife Service.
Dobrow, R. P. (2016). Introduction to stochastic processes with R, John Wiley & Sons.
Lowe, S., Browne, M., Boudjelas, S. and De Poorter, M. (2000). 100 of the world’s worst invasive alien species: A selection from the global invasive species database, Invasive Species Specialist Group, Auck- land.
Wilson, K., Hanks, E. and Johnson, D. (2018). Estimating animal utilization densities using continu- ous?time Markov chain models. Methods in Ecology and Evolution, 9, 1232-1240.
Ko, Y. M. (2011). Study of life-cycle of Rana dyvowskii in JEJU island , Doctoral dissertation, Graduate School Jeju National University, Jeju.
Ministry of Environment (2020). Ecosystem eisturbance organism designation notice 2020-61 in biodiversity
conservation and use act (21-2), Vol. Biodiversity Conservation and Use Act (21-2), Sejong, Ministry
of Environment.
2021, 32
(2)
,351–360
A transition probability estimation of the Bullfrog lifespan using continuous time Markov chain model
†Jayeong Paek1· Heejin Kang2· Ilsu Choi3
13 Department of Mathematics and Statistics, Chonnam National University
2Department of Biology, Chonnam National University
Received 4 February 2021, revised 2 March 2021, accepted 4 March 2021
Abstract
Amphibians are sensitive to environmental changes and are showing a severe de- cline worldwide. Amphibians are known as indicator species sensitive to environmental changes. The bullfrog, which has been introduced into Korea and has spread nation- wide, is an amphibian sensitive to environmental changes and an alien organism requir- ing management measures, and has very important research value. In reality, research on bullfrogs has many limitations in terms of time and cost. In this paper, we estimate the variation in the behavior of the bullfrog ecological features of the bullfrog presented in several papers. The stage of growth over time was applied to the continuous-time Markov chain model to estimate the transition probabilities and mean survival time.
In particular, the average survival time to death at each stage was 104.825 days for eggs, 520.125 days for tadpoles, and 2920.0 days for adults.
Keywords: Bullfrog, continuous time Markov chain model, continuous transition prob- ability, transition rate matrix.
†
This research was supported by Korea Environment Industry & Technology Institute (KEITI) through the Project for the Development of Biological Diversity Threats Outbreak Management Technology (RE201807039), funded by Korea Ministry of Environment (MOE).
1
Ph. D. student, Department of Mathematics and Statistics, Chonnam National University, Gwangju 61186, Korea.
2
Ph. D. student, Department of Biological Science and Biotech, Chonnam National University, Gwangju 61186, Korea.
3