Water Extract of Samultang Reduces Apoptotic Cell Death by H2
7
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
수치
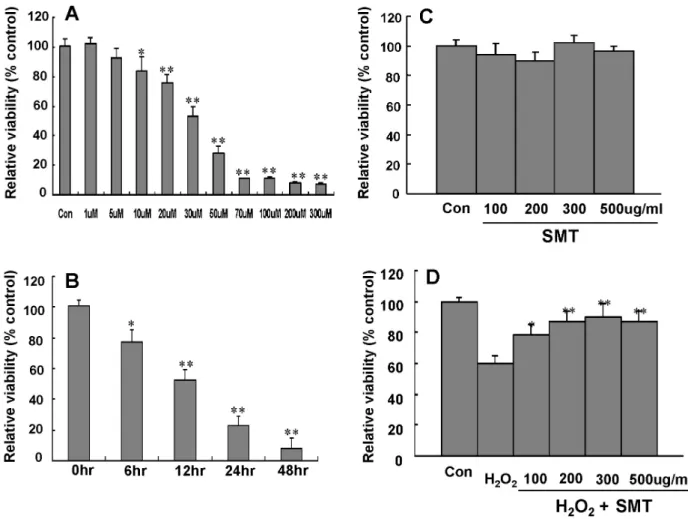
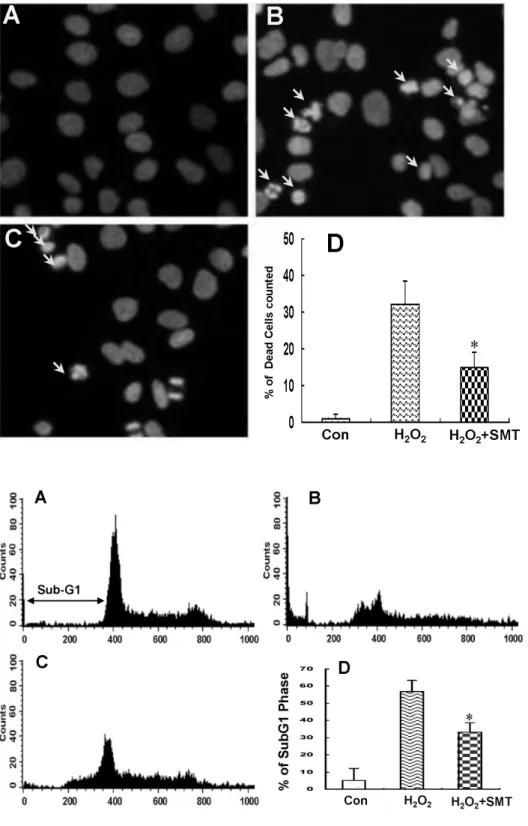
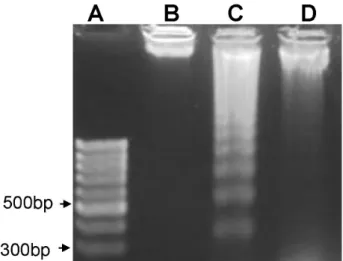
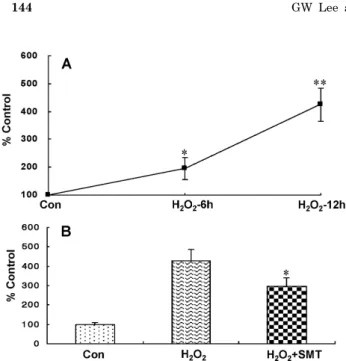
관련 문서
The purpose of this research was to suggest the water quality improvement in streams by evaluating the distribution characteristics of organics and ammonia nitrogen
The purpose of this study was to evaluate the effect of these substances on bone regeneration by applying these materials to bone defects after cyst
The purpose of this study was to investigate the effects of a disability awareness program on improving the attitudes of regular elementary school
interaction with anti-apoptotic Bcl-2 family members, we investigated whether Mcl-1 can inhibit the Noxa-induced cell death.. As expected, Mcl-1 significantly inhibits
A Study on the Development of Ship’s Ballast Water A Study on the Development of Ship’s Ballast Water A Study on the Development of Ship’s Ballast Water A Study on the
Objective: The purpose of this study was to investigate the relationship between the awareness of death and the ethical attitude to Do-Not-Resuscitate (DNR) among some
The purpose of this study is to analyze the effects of service quality on customer satisfaction and revisit intention perceived by Chinese golf range
The purpose of this study is to evaluate the marginal and internal fit of coping made by CAD/CAM using different scanning methods.. Zirconia coping was made
