Ex Vivo 마우스 비장세포에서 법제한 소리쟁이의 LPS에 대한 염증억제 증진 효과
⁃ 연구노트 ⁃
박의성1․송길훈1․이승민1․김태영2․박건영3
1연세대학교 식품영양학과
2(주)가화웰푸드
3차의과학대학교 식품생명공학과
Increased Anti-Inflammatory Effects of Processed Curly Dock (Rumex crispus L.) in Ex Vivo LPS-Induced Mice Splenocytes
Eui-Seong Park1, Gyl-Hoon Song1, Seung-Min Lee1, Tae-Young Kim2, and Kun-Young Park3
1Department of Food and Nutrition, Yonsei University
2Ga Hwa Welfood Co.
3Department of Food Science and Biotechnology, Cha University
ABSTRACT Ex vivo anti-inflammatory effects of curly dock (Rumex crispus L.) with Beopje (processed) in LPS-in- duced mice splenocytes were studied. We formed six experimental groups: no treatment, CD (curly dock), CD-B (curly dock with Beopje), LPS (induced lipopolysaccharide), LPS+CD (LPS+curly dock), and LPS+CD-B (LPS+curly dock with Beopje). LPS increased nitric oxide (NO) production, pro-inflammatory cytokines (TNF-α, IFN-γ, IL-1β, and IL-6), and inflammation related genes (iNOS and COX-2) in mice splenocytes. LPS+CD and LPS+CD-B showed significantly decreased NO production, expression of TNF-α, IFN-γ, IL-1β, and IL-6, and mRNA expression of iNOS and COX-2 compared to the LPS group (P<0.05). Especially, LPS+CD-B showed reduced NO production and mRNA expression of COX-2 at 48 hours and levels of IFN-γ, IL-1β, and IL-6 at 72 hours compared to LPS+CD in mice splenocytes. From the results above, CD-B exhibited the highest anti-inflammatory effects by inhibiting NO production, by regulating pro-inflammatory cytokines and inflammation related genes. These results might be due to the fact that curly dock contains polyphenols, flavonoids, saponin, chrysophanol, emodin, and anthraquinone, and Beopje processing might increase several active compounds and decrease anti-nutritional factors in curly dock.
Key words: splenocyte, curly dock (Rumex crispus L.), LPS, anti-inflammation
Received 7 March 2018; Accepted 1 May 2018
Corresponding author: Kun-Young Park, Department of Food Science and Biotechnology, Cha University, Seongnam, Gyeonggi 13488, Korea
E-mail: [email protected], Phone: +82-31-881-7159
서 론
소리쟁이(Rumex crispus L.)는 마디풀과(Polygona- ceae)에 속하는 다년생 식물로 우리나라 전국 각지에 서식 하며, 뿌리, 잎 각각이 생리활성 효과를 나타내는 것으로 알 려져 있다(1). 소리쟁이는 나물로 먹기도 하고 말리거나 가 공하여 약재로 사용되었으며(2), 섭취했을 때 마우스에 ara- chidonic acid로 유도한 염증반응을 억제시키고 간 보호 효 과가 있으며(3), 항산화(4), 항암 효과(5)와 지방세포의 분화 를 억제하는 것으로 보고되었다(6).
법제는 약재를 가공하여 독성을 줄이고 약효를 증진시키 는 우리나라의 전통적인 방법이며(7), 약재를 씻고 물에 불 리고 찌는 과정을 통해 약재의 anti-nutritional 한 요인을
제거하는 방법이다(8). 말린 생강을 법제하게 되면 6-gin- gerol이 법제하지 않은 생강에 비해 유의적으로 증가하고 (7), 법제한 생강은 일반 생강보다 위염(9), 대장염(10) 발생 을 더욱 억제하는 것으로 보고되어 있다. 옻나무는 주요성분 인 urushiol이 유발하는 T cell 매개성 면역반응으로 인해 사용이 제한되는데, 법제를 통해 urushiol을 제거하면 세포 독성은 줄어들고 폐암세포의 증식은 억제하였다(11).
하지만 지금까지 소리쟁이와 법제한 소리쟁이가 마우스 비장세포에서 lipopolysaccharide(LPS)로 유도된 염증반 응에 대한 연구와 법제와 염증의 관계에 대한 연구도 거의 없다.
만성적인 염증반응은 다양한 장기에 영향을 주고 비만, 당뇨, 심혈관계 질환, 암, 치매, 천식에 영향을 주는 것으로 알려져 있다(12). 다양한 염증관련 요인 중 LPS는 포유류의 세포를 자극하고 LPS binding protein(LBP)이나 toll-like receptor-4(TLR4)와 같은 단백질에 영향을 미친다(13,14).
LPS로 활성화된 TLR4는 nuclear factor κB(NF-κB)를 활 성화시키고, 활성화된 NF-κB는 pro-inflammatory cyto-
kine인 interleukin(IL)-1β, IL-6, tumor necrosis factor- α(TNF-α)와 type 1 interferon(IFN)을 활성화시키며, 핵에 서 염증관련 효소인 nitric oxide synthase(iNOS)와 cy- clooxygenase-2(COX-2)를 활성화시킨다(15-17). Nitric oxide(NO)는 iNOS의 생성을 증진시키고 염증반응의 활성 을 통해 reactive oxygen species(ROS)를 생성하는 것으 로 알려져 있다(18). 따라서 외부 자극으로 인한 염증 관련 유전자의 활성을 억제한다면 염증으로 유도되는 다양한 질 병을 억제할 수 있을 것으로 생각된다.
따라서 본 연구는 일반 소리쟁이와 법제한 소리쟁이를 LPS로 염증반응이 유도된 ex vivo 마우스의 비장세포에 처 리한 후 NO의 생성능과 inflammation 관련 유전인자 TNF- α, IFN-γ IL-1β, IL-6, iNOS, COX-2의 발현 정도를 살펴 보았다.
재료 및 방법
실험재료 준비
소리쟁이와 법제된 소리쟁이 시료는 Ga Wha Wellfood Co.(Jincheon, Korea)에서 제공받아 실험에 사용하였다.
법제 방법은 건조된 소리쟁이를 깨끗이 세척한 다음 4분간 찐 후에 그늘에서 2시간 동안 건조시킨다. 건조된 소리쟁이 는 3분간 볶아서 수분을 제거한 후 2시간 동안 그늘에서 건조시킨 다음 60°C 오븐에서 완전 건조시켰다(19). 건조가 끝난 샘플은 마쇄하여 powder로 만들었으며, 이후 실험을 위해 -20°C에서 보관하였다. 샘플 powder는 각각의 농도 에 맞추어 배지와 혼합한 다음 실험에 이용하였다.
비장세포 배양
비장세포의 채취와 배양은 Amrouche 등(20)의 방법을 다소 수정하여 실험하였다. 5주령의 C57BL/6 마우스(Ori- ent Bio, Seongnam, Korea) 10마리를 구입하여 반입된 즉 시 마우스의 비장을 specific pathogen free(SPF) room에 서 채취하고, 세포의 suspension을 만들기 위해 멸균된 실 험용 가위를 이용하여 잘게 마쇄하였다. 마쇄된 비장세포는 1,500 rpm에 10분간 원심분리 하고 resuspension 하여 free cell을 얻은 후 Dulbecco’s modified Eagle’s medium (DMEM, Sigma-Aldrich Co., St. Louis, MO, USA)에 10%
fetal bovine serum(FBS, Sigma-Aldrich Co.), 100 unit/
mL penicillin-streptomycin(Welgene Inc., Gyeongbuk, Korea)에 배양하여 실험에 사용하였다. 이 연구는 차의과학 대학교 동물윤리위원회의 허가를 받아 진행하였다(IACUC 170149).
NO 농도 측정
세포의 배지 내 NO 농도는 Griess reagent(Sigma- Aldrich Co.)를 이용하여 측정하였다. Hemocytometer를
이용하여 비장세포의 수를 측정하였고, 6-well plate에 well당 1.5×106 cells/mL 농도로 24시간 동안 seeding 하 였다. 배양된 비장세포에 FBS와 penicillin-streptomycin 이 없는 DMEM을 24시간 동안 처리하여 starvation을 유도 하였다. 이후 2 μg/mL LPS(Sigma-Aldrich Co.)와 필터 (0.2 μm, GVS Filtration Inc., Bloomer, WI, USA)를 통과 시킨 0.1 mg/mL 농도의 소리쟁이 시료를 48시간 및 72시 간 동안 처리하였다. 이후 배지를 채취하여 Griess reagent 를 처리하고, Wallac Victor3 1420 Multilabel Counter (PerkinElmer, Wellesley, MA, USA)를 이용하여 550 nm 에서 흡광도를 측정하였다(21).
Enzyme-linked immunosorbent assay (ELISA) Hemocytometer를 이용하여 비장세포의 수를 측정하였 고, 6-well plate에 well당 2×106 cells/mL 농도로 24시간 동안 37°C CO2 incubator에서 배양하였다. 이후 배지는 제 거하고 2 μg/mL LPS와 필터(0.2 μm, GVS Filtration Inc.) 를 통과시킨 0.1 mg/mL 농도의 소리쟁이 시료를 48시간 및 72시간 동안 처리하였다. 배지를 채취한 후 ELISA kit (BioLegend, San Diego, CA, USA)을 이용하여 IL-6, IL- 1β, IFNγ, TNF-α의 농도를 측정하였다. 본 실험은 제조사 에서 제공된 실험방법에 따라 실험을 진행하였다(21).
Real time-quantitative polymerase chain reaction (RT-qPCR)
Hemocytometer를 이용하여 비장세포의 수를 측정하였 고, 6-well plate에 well당 2×106 cells/mL 농도로 24시간 동안 37°C CO2 incubator에서 배양하였다. 이후 배지는 제 거하고 2 μg/mL LPS와 필터(0.2 μm, GVS Filtration Inc.) 를 통과시킨 0.1 mg/mL 농도의 소리쟁이 시료를 48시간 동안 처리하였다. 배지를 제거한 후 Trizol(Invitrogen, Carlsbad, CA, USA)을 이용하여 RNA를 분리하였고, Nano Drop ND-1000(NanoDrop Technologies Inc., Wilming- ton, DE, USA)을 이용하여 정량하였다. 정량된 RNA는 Superscript Ⅱ reverse transcriptase(Invitrogen)를 이용 하여 역전사하였고 cDNA로 합성하였다. cDNA와 각각의 primer, SYBR green(Solis BioDyne, Tartu, Estonia)을 혼합한 후 thermal cycler BioRad CFX-96 real time sys- tem(BioRad, Hercules, CA, USA)을 이용하여 유전자 발 현을 분석하였다. House keeping 유전자로 18S rRNA를 사용하였고, iNOS, COX-2 유전자의 발현을 분석하였으며, 사용된 primer 서열은 다음과 같다: 18S rRNA forward 5’-TCGAGGCCCTGTAATTGGAA-3’, reverse 5’-CCC TCCAATGGATCCTCGTT-3’; iNOS forward 5’-ATGG CTTGCCCCTGGAA-3’, reverse 5’-TATTGTTGGGC TGAGAA-3’; COX-2 forward 5’-GGCAGCAAATCCTT GC-3’, reverse 5’-TATTGTTGGGCTGAGAA-3’(22).
Table 1. Nitric oxide production in mice splenocyte by treatment of LPS, curly dock, and curly dock with Beopje (μM)
48 h 72 h
No treatment CD
CD-B LPS LPS+CD LPS+CD-B
0.93±0.08c 1.01±0.51c 0.77±0.13c 2.39±0.23a 1.48±0.17b 1.06±0.04c
1.38±0.20c 1.34±0.20c 1.00±0.16d 3.15±0.35a 2.43±0.08b 2.20±0.15b LPS: lipopolysaccharide, CD: curly dock (0.1 mg/mL), LPS+
CD: lipopolysaccharide+curly dock (0.1 mg/mL), CD-B: curly dock with Beopje (0.1 mg/mL), LPS+CD-B: lipopolysaccharide +curly dock with Beopje (0.1 mg/mL).
Means (±SD, n=3) with the different letters (a-d) within the same column are significantly different (P<0.05) by Duncan’s multi- ple range test.
통계처리
RT-qPCR을 제외한 모든 실험은 평균±표준편차(stand- ard deviation, SD)로 나타내었으며, RT-qPCR은 평균±표 준오차(standard error, SE)로 나타내었다. Student’s t- test와 one-way analysis of variance(ANOVA)인 Dun- can’s multiple range test를 이용하여 그룹 간의 유의성을 판별하였으며, P value가 0.05 이하일 때 유의성이 있다고 간주하였다. 모든 실험 분석은 SPSS v18 statistical soft- ware package(SPSS Inc., Westlands, Hong Kong)를 이 용하여 분석하였다.
결과 및 고찰
마우스 비장세포에서 NO 생성
Raw 264.7 마우스 대식세포에서 소리쟁이(CD)와 법제 된 소리쟁이(CD-B)는 0.01~0.1 mg/mL의 농도에서 48시 간과 72시간 모두 세포독성이 나타나지 않았다(data not shown). 따라서 0.1 mg/mL의 농도로 실험을 진행하였다.
마우스의 비장세포에서 NO 생성능은 LPS 처리군이 미처리 군에 비해 48시간과 72시간 모두 유의적으로 증가하였다 (Table 1). 그러나 LPS(48 h: 2.39±0.23 μM, 72 h: 3.15±
0.35 μM)군에 비해 LPS+CD(48 h: 1.48±0.17 μM, 72 h:
2.43±0.08 μM)와 LPS+CD-B(48 h: 1.06±0.04 μM, 72 h: 2.20±0.15 μM)를 처리한 군은 NO 생성능이 유의적으로 감소하였다(P<0.05). 또한, 48시간에 LPS+CD-B(1.06±
0.04 μM)는 LPS+CD(1.48±0.17 μM)보다 NO 생성능이 유의적으로 감소하였으나(P<0.05), 72시간에는 큰 차이를 보이지 않았다.
소리쟁이는 폴리페놀과 플라보노이드 성분들이 풍부하고 항산화능이 높은 것으로 보고되어 있으며(23), 한의학에서 는 소리쟁이를 피부병, 비장질환, 임파선 질환과 여러 종양 이나 암의 보조치료제로 이용하기도 한다(4). 생강의 경우 법제하게 되면 법제하지 않은 생강에 비해 6-gingerol의 함 량이 증가하는 것으로 보고되었다(7). 따라서 소리쟁이가 LPS로 유도된 염증반응을 억제하여 NO의 생성을 감소시킨
것으로 생각된다. 또한, 소리쟁이를 법제하면 법제하지 않은 소리쟁이보다 NO의 생성이 더욱 억제된 것은 법제를 통해 독성을 감소시키고, 유효성분을 증진시켜(7,8) 더욱 효과적 으로 염증반응을 억제한 것으로 생각된다.
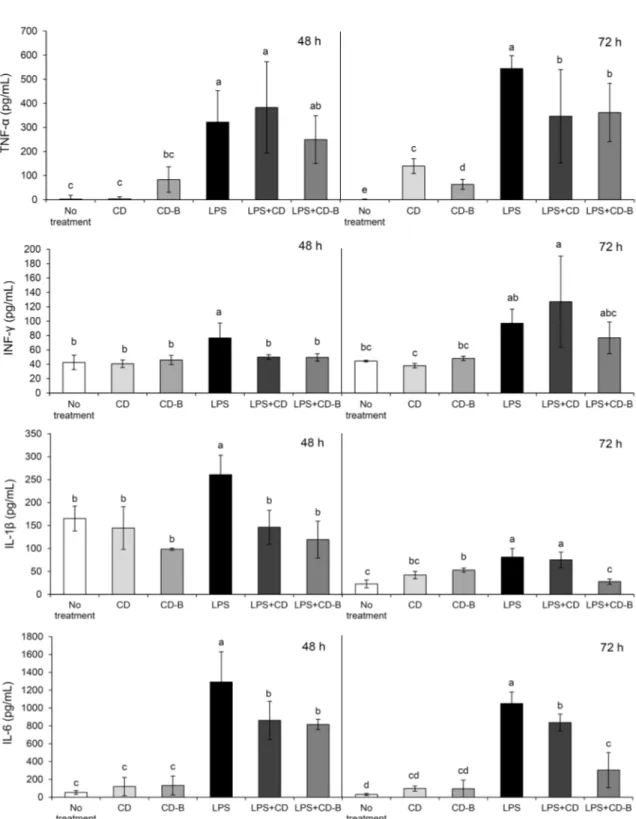
마우스 비장세포에서 pro-inflammatory cytokine의 함량 마우스의 비장세포에 LPS를 처리하면 미처리군에 비해 TNF-α, IFN-γ, IL-1β와 IL-6가 유의적으로 급격히 증가 하였다(P<0.05)(Fig. 1). 하지만 LPS와 함께 CD, CD-B를 처리한 군은 LPS군에 비해 각각의 사이토카인이 유의적으 로 감소하였다(P<0.05). 특히 IFN-γ는 72시간에서 LPS+
CD-B(76.73±21.89 pg/mL)가 LPS+CD(127.07±63.29 pg/mL)보다 감소하였다. 또한, IL-1β는 72시간에 LPS+
CD-B(21.81±5.76 pg/mL)가 LPS+CD(75.20±16.88 pg/
mL)보다 유의적으로 감소하였고(P<0.05), IL-6는 마찬가 지로 72시간에 LPS+CD-B(303.96±195.50 pg/mL)가 LPS +CD(837.27±94.92 pg/mL)보다 유의적으로 감소하여(P<
0.05), 소리쟁이를 법제하였을 때가 법제하지 않을 때보다 72시간에 pro-inflammatory cytokine을 현저히 감소시킨 것으로 나타났다.
T helper cell(Th)은 B cell을 조절하여 염증반응과 im- mune response를 조절한다(24). Th는 그 역할과 종류에 따라 Th1, Th2, Th17, Treg로 분류된다(25). 각각의 Th는 각기 다른 cytokine이 Naive T cell을 자극하여 Th로 분류 되는데, 이때 역할을 담당하는 cytokine을 pro-inflamma- tory cytokine으로 부른다(24). Th1 cytokine의 주요 인자 는 TNF-α와 IFN-γ이고 Th2 cytokine의 주요 인자는 IL- 1β와 IL-6가 있으며(26,27), 이러한 pro-inflammatory cytokine은 전사인자인 NF-κB에 의해 활성화되고(16), 인 체의 염증반응에 관여한다(15).
면역세포에 LPS를 처리하면 염증관련 인자(TNF-α, IFN- γ, IL-1β, IL-6, iNOS, COX-2)가 급격히 증가하는데(21), 이는 LPS 자극으로 염증반응이 활발히 일어나 전반적인 염 증관련 인자가 증가하는 것이다. Ahn 등(28)의 보고에 의하 면 소리쟁이가 dinitrochlorobenzene(DNCB)으로 아토피 를 유도한 Balb/c 마우스에서 아토피 증상이 억제되고 비장 의 무게가 감소하였다. 아토피가 발생하면 Th2 사이토카인 인 IL-1β, IL-6가 증가하게 되는데(27), 본 연구에서도 LPS 로 유도된 염증반응으로 증가한 IL-1β, IL-6가 소리쟁이를 처리하면 감소하는 것으로 나타났다. 그리고 소리쟁이의 항 산화(23) 작용으로 LPS로 유도된 Th1, 2 cytokine을 감소 시키는 것으로도 생각된다. 또한, 법제된 소리쟁이(CD-B) 는 법제되지 않은 것(CD)보다 pro-inflammatory cytokine 을 감소시키는 효과가 컸었는데, Kim 등(10)의 연구에 의하 면 dextran sulfate sodium으로 대장염이 유도된 마우스에 게 법제된 생강과 일반 생강을 섭취시켰을 때 법제된 생강이 TNF-α, IFN-γ, IL-6의 수치를 감소시키는 효과가 컸었다.
이는 법제를 통해 anti-nutritional 한 요인은 제거하고(8),
Fig. 1. Pro-inflammatory cytokines of TNF-α, IFN-γ, IL-1β, and IL-6 in mice splenocyte by the treatment of LPS, curly dock, and curly dock with Beopje. Abbreviations are same as shown in Table 1. Means±SD (n=3). Means with the different letters (a-e) above the bars are significantly different (P<0.05) by Duncan’s multiple range test.
인체에 유용한 물질을 증진시켜 더욱 높은 염증 억제 효과를 나타낸 것으로 생각된다.
마우스 비장세포에서 iNOS, COX-2의 mRNA 발현 iNOS와 COX-2는 pro-inflammatory cytokine과 마찬 가지로 핵에서의 전사인자인 NF-κB에 의해 활성화되며
(16,17), 다양한 염증 지표 중 중요한 인자이다. 마우스의 비 장세포에 LPS를 처리하면 iNOS와 COX-2의 mRNA 발현 이 미처리군에 비해 급격히 증가하였다(P<0.05)(Fig. 2). 하 지만 LPS+CD, LPS+CD-B는 미처리군과 유사한 mRNA 발현 수준을 나타내어 소리쟁이와 법제된 소리쟁이가 LPS 로 과발현된 iNOS와 COX-2의 mRNA 발현을 감소시킨 것
Fig. 2. mRNA expression of inflammation related genes (iNOS and COX-2) in mice splenocyte by the treatment of LPS, curly dock, and curly dock with Beopje (48 h). Abbreviations are same as shown in Table 1. The mRNA expression levels were calculated based on 18S rRNA, which was used as a control (no treatment fold ratio=1). Student t-test (**P<0.01). Means±SE (n=4). Means with the different letters (a,b) above the bars are significantly different (P<0.05) by Duncan’s multiple range test.
으로 나타났다(P<0.05). 특히 COX-2의 mRNA 발현의 경우 LPS+CD에 비해 LPS+CD-B가 유의적으로 감소한 것으로 나타나(P<0.05), 법제된 소리쟁이가 COX-2의 발현을 더욱 효과적으로 감소시킨 것으로 확인되었다.
Raw 264.7 cells에 LPS로 염증반응을 유도한 후 소리쟁 이를 처리하면 염증반응에 관련된 β-hexosaminidase 효소 를 감소시켜(29) 염증 반응을 감소시킨다. 또한, Lee 등(3) 의 연구에 의하면 arachidonic acid(AA)로 ICR 마우스에 염 증반응을 일으킨 후 소리쟁이를 섭취하면 소리쟁이를 섭취 하지 않은 군에 비해 부종을 감소시켰다. AA는 COX path- way를 거쳐 prostglandins과 leukotrienes이 염증반응을 유도하는데(3), 본 연구에서도 소리쟁이가 LPS로 유도된 COX-2의 mRNA 발현을 억제시켜 염증반응을 억제하였고, iNOS까지 감소시켜 이전 연구와 유사한 염증억제 효과를 나타내었다. 특히 법제한 소리쟁이는 법제하지 않는 것보다 COX-2의 발현을 더욱 효과적으로 감소시켰는데, 이는 Kim 등(10)의 연구에서 법제된 생강이 일반 생강보다 대장염이 유도된 마우스에서 iNOS와 COX-2의 발현이 더욱 효과적 으로 감소한 것과 유사하다.
결론적으로 법제는 우리나라의 전통적인 방법으로, 독성은 제거하고(8) 유용한 물질을 높인다(7). 콩을 법제하게 되면 anti-nutritional 요인인 tannin, phytic acid, trypsin in- hibitor를 감소시키고(30), 생강을 법제하게 되면 6-gin- gerol이 유의적으로 증가하게 된다(7). 소리쟁이의 유용성분 은 폴리페놀, 플라보노이드, 사포닌, chrysophanol, emo- din, anthraquinone 등이 알려져 있는데(1), 법제를 하면 이 러한 유용한 성분이 증가하게 되며, 본 연구실의 연구에 의하 면 소리쟁이를 법제하면 chrysophanol이 1.7배 증가하는 것으로 나타나(data not shown), 이로 인해 더욱 높은 항염 증 효과를 나타낸 것으로 생각된다(31). 따라서 이러한 부분 에 대한 추가적인 연구가 필요할 것으로 생각된다.
요 약
소리쟁이(CD)와 법제한 소리쟁이(CD-B)는 ex vivo 마우 스의 비장세포에 LPS를 처리하면 미처리군에 비해 NO의 생성이 증가하였으며, pro-inflammatory cytokine인 TNF -α, IFN-γ, IL-1β와 IL-6가 증가하였고, 염증관련 인자인 iNOS와 COX-2의 mRNA 발현이 증가하였다. 그러나 LPS 를 처리한 비장세포에 CD와 CD-B를 함께 처리하면 LPS를 단독으로 처리한 군에 비해 NO의 생성이 유의적으로 감소 하였고, 염증관련 인자들인 TNF-α, IFN-γ, IL-1β, IL-6, iNOS와 COX-2가 감소하였다. 또한, 48시간에 CD-B가 CD 보다 유의적으로 NO 생성과 COX-2의 mRNA 발현이 감소 하였고, 72시간에 IFN-γ, IL-1β와 IL-6가 감소하였다. 이 는 소리쟁이가 가지고 있는 활성성분들로 인해 항염증 효과 가 나타난 것으로 생각되며, 소리쟁이를 법제하게 되면 이러 한 유용한 물질이 증가되어 일반 소리쟁이보다 더욱 높은 항염증 효과가 나타난 것으로 생각된다.
감사의 글
본 결과물은 농림축산식품부의 재원으로 농림수산식품기술 기획평가원의 고부가가치식품기술개발사업의 지원을 받아 연구되었습니다(116169-3).
REFERENCES
1. Chang SW, Kim IH, Han TJ. 1999. Anthraquinone produc- tivity by the cultures of adventitious roots and hairy from curled dock (Rumex crispus). Korean J Plant Tissue Culture 26: 7-14.
2. Garodia P, Ichikawa H, Malani N, Sethi G, Aggarwal BB.
2007. From ancient medicine to modern medicine: ayurve- dic concepts of health and their role in inflammation and cancer. J Soc Integr Oncol 5: 25-37.
3. Lee S, Kim D, Yim D, Lee SY. 2007. Anti-inflammatory, analgesic and hepatoprotective effect of semen of Rumex
crispus. Kor J Pharmacon 38: 334-338.
4. Park DH, Jeong GT, Lee GM. 2006. Study of antimicrobial and antioxidant activities of Rumex crispus extract. Korean Chem Eng Res 44: 81-86.
5. Shiwani S, Singh NK, Wang MH. 2012. Carbohydrase in- hibition and anti-cancerous and free radical scavenging prop- erties along with DNA and protein protection ability of methanolic root extracts of Rumex crispus. Nutr Res Pract 6: 389-395.
6. Park SJ, Choi JH, Jung YS, Yu MH. 2013. Inhibitory effect of Rumex crispus L. fraction on adipocyte differentiation in 3T3-L1 cells. Korean J Food Sci Technol 45: 90-96.
7. Lee Y, Kim JS. 2003. Studies on the processing of herbal medicines (Ⅵ), HPLC analysis of standard compounds of unprocessed and processed herbal medicines. Korean J Orient Med 9: 69-72.
8. Khattab RY, Arntfield SD. 2009. Nutritional quality of le- gume seeds as affected by some physical treatments 2. Anti- nutritional factors. LWT-Food Sci Technol 42: 1113-1118.
9. Kim SJ, Kim YG, Park KY. 2012. Inhibitory effects of gin- ger and processed (Beopje) ginger extracts on HCl-ethanol induced gastritis in rats. J Korean Soc Food Sci Nutr 41:
1528-1533.
10. Kim SJ, Kim SH, Lim YI, Kim YG, Park KY. 2014. Inhibi- tory effects of ginger and Beopje ginger on DSS-induced colitis in mice. J Korean Soc Food Sci Nutr 43: 477-484.
11. Choi WC, Lee JH, Lee EO, Lee HJ, Yoon SW, Ahn KS, Kim SH. 2006. Study on antiangiogenic and antitumor activ- ities of processed Rhus verniciflua Stokes extract. Korean J Orient Physiol Pathol 20: 825-829.
12. Hotamisligil GS. 2017. Inflammation, metaflammation and immunometabolic disorders Nature 542: 177-185.
13. Gioannini TL, Weiss JP. 2007. Regulation of interactions of Gram-negative bacterial endotoxins with mammalian cells.
Immunol Res 39: 249-260.
14. Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X, Birdwell D, Alejos E, Silva M, Galanos C, Freudenberg M, Ricciardi-Castagnoli P, Layton B, Beutler B. 1998. De- fective LPS signaling in C3H/HeJ and C57BL/10ScCr mice:
mutations in Tlr4 gene. Science 282: 2085-2088.
15. Lu YC, Yeh WC, Ohashi PS. 2008. LPS/TLR4 signal trans- duction pathway. Cytokine 42: 145-151.
16. Xie QW, Kashiwabara Y, Nathan C. 1994. Role of tran- scription factor NF-kappa B/Rel in induction of nitric oxide synthase. J Biol Chem 269: 4705-4708.
17. Lee KM, Kang BS, Lee HL, Son SJ, Hwang SH, Kim DS, Park JS, Cho HJ. 2004. Spinal NF-kB activation induces COX-2 upregulation and contributes to inflammatory pain
hypersensitivity. Eur J Neurosci 19: 3375-3381.
18. Guzik TJ, Korbut R, Adamek-Guzik T. 2003. Nitric oxide and superoxide in inflammation and immune regulation. J Physiol Pharmacol 54: 469-487.
19. Kim SJ. 2012. Preventive effects of beopje ginger and tae- myeongcheong on in vivo gastritis and colitis. MS Thesis.
Pusan National University, Busan, Korea.
20. Amrouche T, Boutin Y, Prioult G, Fliss I. 2006. Effects of Bifidobacterial cytoplasm, cell wall and exopolysaccharide on mouse lymphocyte proliferation and cytokine production Int Dairy J 16: 70-80.
21. Song GH, Park ES, Lee SM, Kim TY, Park KY. 2017. An Atopic Preventive Drink (APD) reduces Th2 cytokines in LPS-treated RAW 264.7 cells. Tang 7: 15.1-15.6.
22. Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25: 402-408.
23. Kim HJ, Hwang EY, Im NK, Park SK, Lee IS. 2010. Anti- oxidant activities of Rumex crispus extracts and effects on quality characteristics of seasoned pork. Korean J Food Sci Technol 42: 445-451.
24. Vinuesa CG, Tangye SG, Moser B, Mackay CR. 2005. Fol- licular B helper T cells in antibody responses and auto- immunity. Nat Rev Immunol 5: 853-865.
25. Zou W, Restifo NP. 2010. TH17 cells in tumour immunity and immunotherapy. Nat Rev Immunol 10: 248-256.
26. Grewe M, Bruijnzeel-Koomen CA, Schöpf E, Thepen T, Langeveld-Wildschut AG, Ruzicka T, Krutmann J. 1998.
A role for Th1 and Th2 cells in the immunopathogenesis of atopic dermatitis. Immunol Today 19: 359-361.
27. Helmby H, Grencis RK. 2004. Interleukin 1 plays a major role in the development of Th2-mediated immunity. Eur J Immunol 34: 3674-3681.
28. Ahn JY, Im LR, Kim JH, Park JH, Kim DK, Lee YM. 2009.
Effects of Rumecis Radix water extract on development of atopic dermatitis in BALB/c mice. Kor J Pharmacogn 40:
218-223.
29. Park JA, Choi MO. 2011. Antimicrobial activity and anti-in- flammation effect to the human skin pathogens by the Rumex crispus L. root extracts. Kor J Aesthet Cosmetol 9: 9-16.
30. Sharma A, Kumari S, Wongputtisin P, Nout MJ, Sarkar PK.
2015. Optimization of soybean processing into kinema, a Bacillus-fermented alkaline food, with respect to a mini- mum level of antinutrients. J Appl Microbiol 119: 162-176.
31. Kim SJ, Kim MC, Lee BJ, Park DH, Hong SH, Um JY.
2010. Anti-inflammatory activity of chrysophanol through the suppression of NF-kappaB/caspase-1 activation in vitro and in vivo. Molecules 15: 6436-6451.