240 서 론
쥐방울덩굴과에 속하는 다년생 초본식물인 족도리풀 (Asarum sieboldii Miq)은 우리나라 전역에 분포하고 있으나
그 서식 환경이 조도는 낮고 고도는 높은 북향 내지 북 동향에 치우쳐 있으며 토양의 수분 함량이 많고 부엽층 이 두꺼운 아주 오래된 활엽수림 아래에서 주로 자생하 는 것으로 보고된 바 있다.1) 형태학적으로는 봄에 2개의 잎이 마주 나오고 거의 동시에 족도리 모양의 자주색 꽃
인체 암세포의 증식에 미치는 세신 열수 추출물의 영향
1경성대학교 교육대학원 및 자연과학대학 생물학과,
2동의대학교 한의과대학 생화학교실 및 대학원 바이오물질제어학과
김옥숙1․박 철2․문성기1․최영현2
Anti-proliferative Effects of Water Extract of Asarum sieboldii in Human Cancer Cells
Ok-Suk Kim1, Cheol Park2, Sung-Gi Moon1 and Yung Hyun Choi2
1Department of Biology, College of Science and Graduate School of Education, Kyungsung University, Busan 608-736,
2Department of Biochemistry, Dongeui University College of Oriental Medicine and
Department of Biomaterial Control (BK21 Program), Dongeui University Graduate School, Busan 614-052, Korea
Asarum sieboldii, as a medicinal and food plant, has been used to treat pain and inflammation in traditional Korean medicine. In the present study, it was analyzed the effects of water extracts of A.
sieboldii (WEAS) on the growth and apoptosis of human lung carcinoma (A549 and NCI-H460) and colon cancer (HT29) cell lines. Treatment with WEAS in A549 cells inhibited the cell growth and induced apoptosis in a dose-dependent manner as measured by MTT assay, however NCI-H460 and HT29 cells exhibited relatively weak anti- proliferative activities to WEAS. RT-PCR and Western blot data revealed that the anti-proliferative effect of WEAS in A549 cells was associated with a dose-dependent up-regulation of cyclin-dependent kinase inhibitor p21 (WAF1/CIP1) protein in a tumor suppressor p53-independent fashion. However the levels of proliferating cell nuclear antigen (PCNA) mRNA and protein expression were markedly down-regulated by WEAS treatment. It was also found WEAS treatment decreased the levels of cyclooxygenase (COX)-2 and inducible nitric oxide synthease (iNOS) expression without significant changes in the expression of COX-1. Though further studies will be needed to identify the active compounds that confer the anti-cancer activity of WEAS, the present findings provide important new insights into the possible molecular mechanisms of the anti-cancer activity of A. sieboldii. (Cancer Prev Res 11, 240-247, 2006)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Asarum sieboldii, p21, PCNA, COX-2, iNOS
책임저자:최영현, ꂕ 614-052, 부산광역시 부산진구 양정동 산 45 동의대학교 한의과대학 생화학교실
Tel: 051-850-7413, Fax: 051-853-4036 E-mail: [email protected]
접수일:2006년 8월 9일, 게재승인일:2006년 9월 5일
Correspondence to:Yung Hyun Choi
Department of Biochemistry, Dongeui University College of Oriental Medicine, San 45, Yangjeong-dong, Busanjin-gu, Busan 614-052, Korea Tel: +82-51-850-7413, Fax: +82-51-853-4036
E-mail: [email protected]
이 피며, 줄기는 땅속줄기이고 마디가 있으며 마디마다 뿌리가 나온다. 족도리풀의 뿌리는 가늘고 길며, 맛이 맵 다고 하여 세신(細辛)이라고 부르며 예로부터 한약 재료 로서 널리 사용되고 있다.2)
세신에는 2∼3%의 정유성분이 함유되어 있고, methy- leugenol, safrol, eucarvone, elemicin, β-pinene, higenamine, asarinin 및 γ-asarone 등이 주요 정유 성분인 것으로 알려 져 있다.3) 이 중 methyleugenol과 safrol은 세신의 주요 지 표성분으로서 methyleugenol은 xanthoxylol과 마찬가지로 기침억제 작용 및 항알레르기에 효과가 있으며,4) safrol은 asaricin과 함께 살충작용 및 항균활성이 있는 것으로 보 고된 바가 있고, 뇌세포 보호기능과 치매 예방 효과 등도 보고된 바 있다.5) 이러한 선행연구의 내용으로 미루어 세신에 함유되어 있는 다양한 성분들이 암의 발생이나 암세포의 증식 억제 가능성이 매우 높을 것으로 예상되 지만, 세신 추출물을 대상으로 항암작용에 관한 연구는 현재까지 이루어진 바가 없는 실정이다.
한편 세포의 죽음은 necrosis와 apoptosis로 구분되며 이 것은 세포의 형태적 및 생화학적인 특성에 의하여 구분 될 수 있다. Necrosis는 생리적, 화학적인 외상에 의한 세 포의 죽음이고 apoptosis는 개체의 정상적인 발달과 분화 에 관여하며, DNA 손상, 바이러스 감염 등에 의한 유전 적 조절 하에서 일어나는 정교한 방어기전이라는 점에 서 생리적이거나 화학적인 외상에 의한 세포의 죽음인 necrosis와는 구별된다.6) 또한 apoptosis는 개체보존의 수 준에서 손상된 세포들의 제거를 위한 중요한 수단이며, apoptosis의 유발에 종양억제유전자 p53, Bcl-2 family 및 caspase member 등과 같은 유전자가 관여한다는 사실이 알려지고 apoptosis와 연관된 분자적 기전이 최근 많이 밝혀지면서 apoptosis의 유발은 암세포의 성장, 증식의 억제와 암세포 제거의 한 방법으로써 널리 연구되고 있 다.7)
세포의 성장과 생존에서 세포주기에 관한 개념은 20 세기 이전에 광학 현미경의 관찰을 통한 간기세포와 유 사 분열기 세포들의 구별이 가능해진 후, 1950년대에 Swift 및 Howard에 의해 G1, S, G2 및 M기에 대한 정의가 내려지면서 성립되었다.8) 그 후 생화학 및 분자생물학적 실험방법들의 폭발적인 성장과 더불어 1980년대에 세포 주기조절의 양성인자라 할 수 있는 cyclin들이 동정되었 으며, cyclins의 합성, 인산화 및 분해 과정 등의 연구를 통해서 이들이 전 세포주기에 걸쳐 매우 다양하게 존재 한다는 것이 밝혀졌다.9) 먼저 D-type cyclin은 세포가 G1 기로 접어들면서 발현이 증가되기 시작하여 cyclin-de- pendent kinase (Cdk) 4 및 Cdk6와 복합체를 형성하면서
G1기 조절을 담당하고 있다.10) 또한 cyclin E는 Cdk2와 복 합체를 이루면서 S기로의 전이에 중요한 역할을 담당하 고 있는 것으로 알려져 있다.11) Cyclin A는 G1 후기에 발 현을 시작하여 S기와 G2기 동안 발현이 증가되고 cyclin B1은 G2기에서 M기로의 전이를 조절하며, 각각은 Cdk2 및 Cdc2의 kinase 활성을 위한 양성 조절자로서 중요한 역할을 한다.12) 한편 Cdks는 다양한 세포증식 억제 신호 들에 의해 유도되는 Cdk inhibitor에 의하여 그 활성이 억 제되는데, 크게 INK4 family 및 CIP/KIP family 두 가지로 분류되고 있다.13,14) 그중 Cdk2와 결합하면서 그들의 활 성을 저해하는 것으로 알려진 CIP/KIP family에 속하는 p21 및 p27은 DNA 손상에 의한 종양 억제인자인 p53에 의해 조절을 받는다고 알려져 있다.15) 이런 세포주기 조 절 인자와 그 외의 분자들의 상호 작용에 의해 그 주기 가 조절되며 또한 그 인자들의 변화에 의해 주기가 억제 또는 증폭되는 것이다. 따라서 세포주기에 대한 연구는 암세포의 과다한 증식과 성장억제에 대한 연구에 필연 적인 것으로 사료된다.
본 연구에서는 2종류의 인체 폐암세포와 대장암세포 의 증식에 미치는 세신 추출물의 영향을 조사하였으며, 그중에서 가장 감수성이 뛰어난 A549 인체 폐암세포의 세포주기에 미치는 영향, 그리고 apoptosis의 유발에 관한 연구를 실시하였다.
재료 및 방법 1. 실험 재료 및 세포배양
본 실험에 사용된 족도리풀은 2005년 부산의 금정산 과 금련산, 장산에서 채집한 것을 말려 그 뿌리(세신)만 을 사용하였으며, 100 g을 1,000 ml의 증류수에 3시간 이 상 끓인 후, 3,000 rpm으로 20분간 원심 분리시켜 침전물 을 제거하였다. 이를 다시 0.45μm의 여과지를 이용하여 부유 성분을 걸러낸 후 수용성분을 동결 건조하여 사용 하였다. 준비된 세신 열수 추출물(water extract of A.
sieboldii, WEAS)은 3차 증류수에 용해하여 멸균과 여과 과 정을 거쳐 배지에 적정 농도로 희석하여 처리하였다.
실험에 사용한 A549 인체 폐암세포, NCI-H460 인체 폐암세포, HT29 인체 대장암세포는 생명공학연구소 (KRIBB, Taejeon, Korea)에서 분양 받았으며, 세포의 배양 과 시약처리를 위해 10%의 우태아혈청(fetal bovine serum, FBS, Gibco BRL, Grand Island, NY, USA)과 1%의 peni- cillin-streptomycin (Gibco BRL) 등이 포함된 RPMI-1640 배 지(Gibco BRL)를 사용하였다. 세포는 37oC, 5% CO2 조건 하의 CO2 incubator에서 배양하였고, 세포수의 증식에 따
른 과밀도 현상에 의한 세포의 성장억제나 각종 오염을 방지하기 위해 0.05% trypsin-ethylenediamine tetraacetic acid (EDTA, Gibco BRL)를 처리하여 세포를 부유시킨 다음 적 정한 수의 세포를 분주하여 재배양하였다.
2. MTT assay를 이용한 세포 성장률의 측정 및 형태 변화 관찰
세포배양용 6 well plate에 각각의 암세포들을 well당 3×104의 개수로 분주하고 24시간 동안 안정화시킨 후 세신 추출물을 배지에 적정농도로 처리한 후 배양하였 다. 72시간 후 배지를 제거하고 tetrazolium bromide salt (MTT, Sigma, St. Louis, MO, USA)를 0.5 mg/ml 농도로 희 석하여 2 ml씩 분주하고 3시간 동안 처리하였다. 3시간 후 MTT 시약을 제거하고 DMSO를 1 ml씩 분주하여 well 에 생성된 formazan을 모두 녹인 후 96 well에 200μl씩 옮 겨서 ELISA reader (Molecular Devices, Sunnyvale, CA, USA) 를 이용하여 540 nm에서 흡광도를 측정하였다. 3번의 측 정값과 평균값을 표준 오차를 Microsoft EXCEL program을 이용하여 분석하여 그래프로 나타내었다. 세신 추출물 처리에 따른 암세포의 형태 변화를 관찰하기 위하여 동 일한 조건에서 72시간 동안 세신 추출물을 적정 농도로 처리한 후 위상차 현미경을 이용하여 200배의 배율 하에 서 정상 및 세신 추출물이 처리된 배지에서 배양된 세포 의 형태를 비교하였다.
3. DNA flow cytometry에 의한 세포 주기의 분석
정상 및 세신 추출물을 처리한 배지에서 72시간 동안 배양시킨 암세포를 PBS로 씻어 내고 0.05% trypsin-EDTA 를 처리하여 부유시킨 다음 1,000 rpm으로 10분간 원심 분리하여 상층액을 버리고 세포들만 모았다. 여기에 다 시 PBS를 첨가하여 충분히 씻은 다음 1,000 rpm으로 10 분간 원심분리 한 후 상층액만 버리고 남은 세포에 0.5 ml의 PBS로 잘 부유시키고, 차가운 ethanol 0.5 ml을 첨가 하여 4oC에서 한 시간 동안 고정시켰다. 5×106개의 고정 된 세포들을 원추형 vial에 넣어서 1,000 rpm으로 수분간 원심 분리하여 상층액을 제거하고, 1% bovine serum albumin (BSA, Sigma)이 함유된 PBS로 2∼3회 washing 과 정을 거친 후 다시 수분간 원심 분리하였다. 세포 침전물 을 1% BSA를 함유한 PBS 0.8 ml로 부유시키고 핵산에 특이적으로 결합하는 형광물질인 DNA intercalating dye propidium iodide (PI, concentration, 50μg/ml; Sigma)와 0.1 mg/ml의 RNase (Sigma)를 처리하여 암실(4oC)에서 1시간 동안 염색과정을 거쳤다. PBS로 두 번 washing 과정을 거 친 후, 부유액을 만들고, 35μm pore size의 nylon mesh에
부유액을 pipette으로 통과시켜 단일 세포로 분리시킨 후 DNA flow cytometry (Becton Dickinson, San Jose, CA, USA) 에 적용시켜 형광반응에 따른 histogram을 ModiFit LT (Becton Dickinson) program을 사용하여 분석하였다.
4. RT-PCR에 의한 mRNA 발현의 분석
동일한 조건에서 준비된 암세포를 대상으로 TRIzol B (Invitrogen, Carlsbad, CA, USA)를 이용하여 total RNA를 분 리하였다. 분리된 RNA를 정량한 후, ONE-STEP RT-PCR PreMix Kit (Intron, Biotechnology, Korea)를 이용하여 2μg 의 RNA에서 cDNA를 합성하였다. 이 cDNA를 template로 사용하여 관찰 대상 유전자(Table 1)를 polymerase chain reaction (PCR) 방법으로 증폭하였다. 이때 housekeeping 유 전자인 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 유전자를 internal control로 사용하였다. 각 PCR 산물들을 1% agarose gel을 이용하여 전기영동하고 ethidium bro-
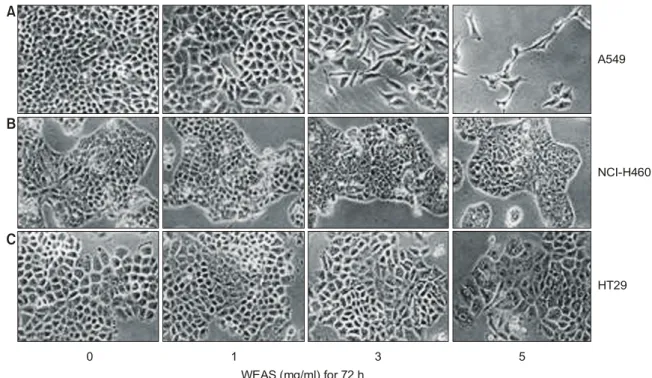
Table 1. Gene-specific primers for RT-PCR
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Gene name Sequence
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ GAPDH Sence 5'-CGG-AGT-CAA-CGG-ATT-
TGG-TCG-TAT-3'
Antisence 5'-AGC-CTT-CTC-CAT-GGT- GGT-GAA-GAC-3'
COX-1 Sence 5'-TGC-CCA-GCT-CCT-GGC- CCG-CCG-CTT-3'
Antisence 5'-GTG-CAT-CAA-CAC-AGG- CGC-CTC-TTC-3'
COX-2 Sence 5'-TTC-AAA-TGA-GAT-TGT- GGG-AAA-AT-3'
Antisence 5'-AGA-TCA-TCT-CTG-CCT- GAG-TAT-CTT-3'
iNOS Sense 5'-AGA-GAG-ATC-CGG-TTC- ACA-3'
Antisense 5'-CAC-AGA-GCT-GAG-GGT- ACA-3'
p53 Sense 5'-GCT-CTG-ACT-GTA-CCA- CCA-TCC-3'
Antisense 5'-CTC-TCG-GAA-CAT-CTC- GAA-GCG-3'
p21 Sense 5'-CTC-AGA-GGA-GGC-GCC- ATG-3'
Antisense 5'-GGG-CGG-ATT-AGG-GCT- TCC-3'
PCNA Sense 3'-GCT-GAC-ATG-GGA-CAC- TTA-5'
Antisense 3'-CTC-AGG-TAC-AAA-CTT- GGT-G-5'
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
mide (EtBr, Sigma)로 염색한 후 ultra violet (UV)하에서 확 인하였다.
5. Western blot analysis에 의한 단백질 발현의 분석
정상 및 세신 추출물이 처리된 배지에서 자란 암세포 들을 lysis buffer로 용해한 후, 고속원심분리로 세포 내 잔 사물을 분리시킨 후 동량의 단백질을 SDS-polyacrylamide gel 전기영동으로 분리하였다. 분리된 단백질을 함유한 acrylamide gel을 nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electroblotting에 의해 전이 시킨 후, 10% skim milk를 함유한 PBS-T (0.1% Tween 20 in PBS)에 4oC에서 1시간 이상 배양하면서 비특이적인 단 백질들에 대한 blocking을 실시하였다. 그리고 특정 단백 질에 대한 항체를 membrane에 적용시켜 항원 항체 반응 을 일으킨 후, PBS-T로 씻어내고 특정 항체에 대한 이차 항체 반응을 실시한 후 ehanced chemiluminoesence (ECL) 용액(Amersham Life Science Corp., Arlington Heights, IL, USA)을 적용시킨 다음 X-ray film에 감광시켜 특정 단백 질의 양을 분석하였다. 본 실험에 사용된 항체들은 Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA) 및 Calbiochem (Cambridge, MA, USA)에서 구입하였으며, 이차 항체로 사 용된 horseradish peroxidase-labeled donkey anti-rabbit immu- noglobulin 및 peroxidase-labeled sheep anti-mouse immuno- globulin은 Amersham Corp.에서 구입하였다.
결과 및 고찰
1. 암세포의 증식에 미치는 세신 추출물의 영향
3가지 종류의 인체 암세포 증식에 세신 추출물의 각 처리 농도별로 어떤 영향을 미치는지에 대해 MTT assay 를 이용하여 분석하였다. MTT assay는 세포의 생존정도 를 간접적으로 측정하는 방법으로 세포내 소기관인 미 토콘드리아의 탈수소 효소 작용에 의하여 노란색의 수 용성 기질인 MTT tetrazolium을 청자색을 띠는 비수용성 의 MTT formazan으로 환원시키는 미토콘드리아의 능력 을 이용하는 검사법이다. 준비된 암세포에 적정 농도의 세신 추출물을 72시간 처리 후 540 nm 파장에서 흡광도 를 조사한 결과는 Fig. 1에 나타낸 바와 같다. 결과에서 알 수 있듯이 A549 폐암세포의 경우 3 mg/ml 처리군에서 약 20% 이상의 증식 억제를 보였으며, 4 mg/ml 및 5 mg/ml 처리군에서는 약 58% 및 72%의 증식 억제 효과 가 나타나 세신 추출물 처리 농도 의존적으로 암세포의 증식이 매우 억제되었음을 알 수 있었다. 그러나 동일 조건에서 NCI-H460 폐암세포에서는 최고 20% 미만의
증식 억제 효과가 있었으며, HT29 대장암세포에서는 5 mg/ml 처리군에서도 거의 증식 억제 효과가 나타나지 않았다. 이러한 결과는 trypan blue 염색에 의한 hemocy- tometer count 분석에서도 유사하게 나타났었다(data not shown). 즉 세신 추출물은 조사된 3가지 암세포주 중, A549 세포에서만 특이적으로 높은 항암활성을 보여주 었다.
2. 세신 추출물이 암세포의 형태에 미치는 영향
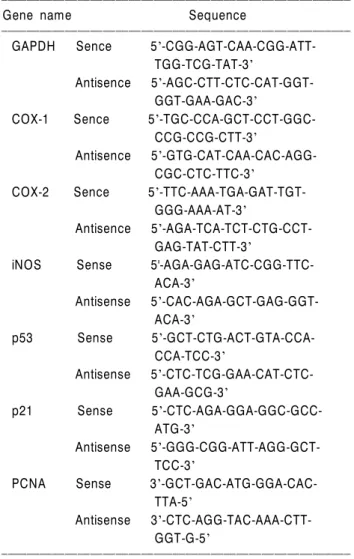
세신 추출물 처리에 따른 처리농도 의존적인 증식 억 제와 연관된 암세포의 전체적인 형태 변화를 알아보기 위해서 A549 폐암세포, NCI-H460 폐암세포, HT29 대장 암세포에 세신 추출물을 각 농도별로 처리하여 72시간 동안 배양한 후 위상차 현미경을 이용하여 관찰하였다.
Fig. 2에서 나타낸 바와 같이 A549 인체 폐암세포에서 세 신 추출물의 농도 증가에 따른 암세포의 심한 형태적 변 화가 관찰되었고 밀도가 감소하였다. 특히 세신 추출물 의 농도가 증가할수록 membrane shrinking 현상을 관찰할 수 있었고 길고 분지를 형성하는 듯한 dendrite-like한 구 조로 바뀌었으며, 부착력을 상실하고 배지 위로 부유되 는 경향성이 증가함을 알 수 있었다. 그러나 NCI-H460 및 HT29 세포주에서는 세신 추출물의 처리에 따른 큰 형태적인 차이를 관찰할 수 없었다. 따라서 A549 인체 폐암세포의 세신 추출물 농도 의존적인 형태적 변화와 밀도의 감소는 Fig. 1에서 관찰된 세신 추출물 처리에 따
Fig. 1. Anti-proliferative effects of water extract of Asarum sie- boldii (WEAS) in human lung carcinoma (A549 and NCI-H460) and colon cancer (HT29) cells. Cells were seeded as described in materials and methods, and treated with various concen- trations of WEAS. After 72 h incubation with WEAS, MTT assay was performed. Results are expressed as the means±S.E. of three independent experiments.
른 증식 억제 결과와 매우 일치되는 결과임을 알 수 있 었다.
3. A549 세포의 세포주기에 미치는 세신 추출물의 영향
다음은 세신 추출물 처리에 의하여 가장 항암활성이 높았던 A549 폐암세포를 대상으로 세신 추출물에 의한 증식 억제 현상과 연관된 세포주기 분포도 변화를 조사
하였다. 이를 위하여 각 농도별 세신 추출물을 처리한 후 PI을 이용하여 세포의 핵을 염색한 후 DNA flow cytometry를 이용하여 세포주기 각 기에 해당되는 세포의 빈도를 조사하였다. Fig. 3 및 Table 2에서 나타낸 바와 같이 정상 배지에서 자란 A549 세포의 경우 G1기에 해 당되는 세포의 빈도가 67.57%였으며, S기 및 G2/M기의 경우 각각 18.54% 및 13.89% 정도였다. 그러나 세신 추 출물이 처리된 배지에서 배양된 암세포의 경우에도 정 Fig. 2. Morphological changes in human lung carcinoma (A549 and NCI-H460) and colon cancer (HT29) cells following incubation with WEAS. Exponentially growing cells were incubated with either vehicle alone (0) or WEAS (1∼5 mg/ml) for 72 h. Cell morphology was visualized by light microscopy. Magnification, X200.
A
B
C
A549
NCI-H460
HT29
0 1 3 5
WEAS (mg/ml) for 72 h
Fig. 3. DNA-flourescence histogram of A549 cell nuclei after treatment with WEAS. Exponentially growing cells at 50% confluency were treated for 72 h with indicated concentrations of WEAS. Cells were trypsinized and pellets were collected. The cells were fixed and digested with RNase, and then cellular DNA was stained with PI, and analyzed by flow cytometry.
Cell number
G1
S
G2M Sub-G1
0 1 2 3 4 5
WEAS (mg/ml) for 72 h
DNA content
상 배지에서 배양된 세포와 비교하여 큰 차이는 나지 않 았지만, G1 및 G2/M기에 속하는 세포의 빈도가 다소 감 소하고 S기가 상대적으로 다소 증가하였다. 따라서 세신 추출물 처리에 의한 A549 인체 폐암세포의 증식억제는 거의 전 세포주기를 통하여 이루어지나 그 중 특히 S기 에서 다소 많이 교란 받을 수 있는 가능성을 보여준다.
그리고 apoptosis의 정량적 지표로 사용되는 sub-G1기에 해당되는 세포의 빈도는 Fig. 3에 나타낸 바와 같이 세신 추출물 처리 농도의 높아짐에 따라 점점 증가되어 정상 배지에서 자란 세포의 경우 3% 이하로 나타났으나, 3 mg/ml 및 5 mg/ml 처리군에서는 각각 약 15% 및 22%
정도였다. 따라서 세신 추출물 처리에 따른 A549 폐암세 포의 증식억제는 apoptosis 유발과 밀접한 연관성이 있음 을 알 수 있었다.
4. p53, p21WAF1/CIP1 및 PCNA의 발현에 미치는 세신 추출물의 영향
Cdk inhibitor는 cyclin/Cdk complex와 결합하여 그 활성 을 억제하는 것으로 알려져 있는데,11) 특히 CIP/KIP 군에 속하는 p21은 종양 억제유전자인 p53에 의하여 활성화 되어 G2/M기의 진행을 억제하기도 한다.16) Cdk inhibitor p21의 활성화에는 p53이 관여하는 것이 일반적이지만 세포주에 따라서 p53 비의존적인 경로를 통하여 p21이 활성화된다고도 알려져 있다.17,18) 세신 추출물처리에 의 한 A549 인체 폐암세포의 증식억제가 종양억제 유전자 또는 세포주기 조절억제자의 발현과 상관성이 있는지 조사하기 위하여 종양 억제 유전자 p53 및 Cdk inhibitor 인 p21의 발현을 RT-PCR 및 Western blot 분석법으로 조 사하였다. Fig. 4의 결과에서 알 수 있듯이 전사 및 번역 수준에서 세신추출물 처리 농도 의존적으로 p53의 발현 이 다소 감소되었으나 p21은 세신 추출물의 농도 의존적
으로 단백질 발현이 증가하였다. 그러나 세포주기 진행 전반에 걸쳐 세포의 증식을 촉진하는 것으로 알려진 proliferating cell nuclear antigen (PCNA)의 발현은19) WEAS 처리 농도가 증가할수록 전사 및 번역 수준에서 모두 감 소되었다. 따라서 세신 추출물을 처리했을 경우 p21이 p53 비의존적인 경로를 통하여 증가하게 하는 것을 알 수 있으며 p21의 증가가 암세포 주기 교란과 연관성을 가질 수 있음을 알 수 있다. 이는 매우 흥미로운 결과라 고 생각되며 이와 연관된 뚜렷한 증거를 제시하기 위해 서는 세신 추출물 처리에 의한 인체 암세포의 세포주기 교란 및 관련 유전자들의 kinase 활성 검사 등을 통한 후 Table 2. Fractions of each cell cycle phase of A549 human lung
carcinoma cells cultured in the presence or absence of various concentrations of WEAS
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
% of cell
Weas (mg/ml) ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
G1 S G2/M
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
0 67.57 18.54 13.89
1 69.22 21.24 9.54
2 66.45 23.64 9.91
3 63.39 26.11 10.50
4 67.03 22.59 10.38
5 65.59 23.42 10.99
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Fig. 4. Effects of WEAS treatment on the levels of tumor sup- pressor p53, Cdk inhibitor p21 and PCNA expression in A549 human lung carcinoma cell. (A) After 72 h incubation with WEAS, total RANs were isolated and reverse-transcribed. The resulting cDNAs were sujected to PCR with p53, p21 and PCNA primers, and reaction products were subjected to electro- phoresis in a 1% agarose gel and visualized by EtBr staining.
GAPDH was used as an internal control. (B) Cells were incubated with WEAS for 72 h, lysed and cellular proteins were separated by 12% SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with anti-p53, anti-p21 and anti-PCNA antibodies. Proteins were visualized using ECL detection system. Actin was used as a loading control.
A 0 1 2 3 4 5
WEAS (mg/ml)
p53
p21
PCNA GAPDH
B 0 1 2 3 4 5
WEAS (mg/ml)
p53
p21 PCNA
Actin
속연구가 진행되어야 할 것이다.
5. Cyclooxygenase의 발현에 미치는 세신 추출물의 영향
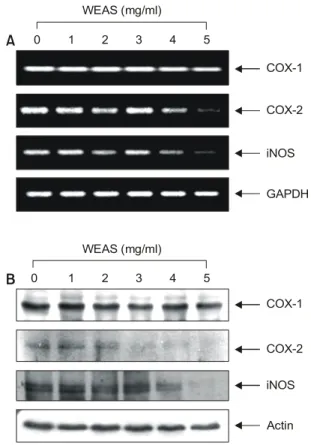
한편 염증을 포함한 다양한 생체 반응에서 prostaglandin 계열 물질들은 세포의 생존이나 증식에 영향을 줌으로 써 각종 인체 질병의 유발과 진행에 중요한 역할을 함이 최근 밝혀지고 있다.20) Prostaglandin의 합성에서 2가지의 cyclooxygenase (COX) isoform이 관여하고 있는데, 대부분 의 조직에서 일정한 수준으로 발현되는 COX-1의 경우 인체의 항상성 유지와 연관된 기능수행에 관여하며, COX-2는 성장인자, cytokines, 종양촉진인자 등의 자극에 의한 유도성 isoform으로서 암을 포함한 세포의 성장 및
분화와 연관된 각종 퇴행성 질환의 발병과 진행에 중요 한 역할을 한다.21) 역학적 조사와 여러 종류의 암 조직에 서 COX-2와 inducible nitric oxide synthease (iNOS)가 높은 수준으로 발현되고 있으며 이들 두 유전자의 발현을 증 대시킬 경우 apoptosis에 대한 저항성을 가지는 점으로 보 아 특히 염증반응과 함께 세포의 암화에 COX-2는 iNOS 와 밀접한 관계를 가진다고 볼 수 있다.22,23) 또한 COX-2 의 과발현에 의해 암조직에서의 혈관신생 및 전이능이 높아지고 apoptosis를 막는다는 점과 COX-2 specific in- hibitor에 의한 angiogenesis와 종양형성의 억제 등의 결과 에서 이 유전자의 선택적 조절에 의한 암 예방전략이 대 두되고 있다.24) 본 실험에서는 A549 인체 폐암세포에 대 한 세신 추출물의 처리가 이런 COXs 및 iNOS의 발현에 어떤 영향을 주는지를 조사하였다. Fig. 5에서 보여주듯 이 COX-1은 전사수준과 단백질 발현 양쪽에서 변화가 없었지만 COX-2와 iNOS는 전사수준과 단백질 발현 양 쪽 모두에서 세신 추출물의 처리농도 의존적으로 그 양 이 감소하였다. 따라서 세신 추출물 처리에 따른 A549 폐암세포의 증식억제는 COX-2 및 iNOS의 발현 저하와 연관된 apoptosis 유발과 관련이 있음을 알 수 있었으며, 세신 추출물은 암예방 및 염증성 질환 조절제로서의 연 구 가치가 높은 것으로 생각된다.
결 론
본 실험에서는 족도리풀(Asarum sieboldii Miq) 뿌리(세신) 의 열수 추출물(WEAS)에 의한 인체 암세포의 성장 및 apoptosis 유발에 관한 내용을 분석하였다. 세신 추출물의 처리에 의해 조사된 인체 암세포에서는 세신 추출물 농 도 의존적으로 세포의 생존율 감소와 증식 억제가 관찰 되었으며, 이러한 세신 추출물의 암세포의 성장 억제 효 과는 apoptosis의 유발과 밀접한 관련이 있었다. 이는 특 히 A549 폐암세포에서 종양억제 유전자 p53 비존적인 Cdk inhibitor p21의 발현 증가 및 PCNA 발현의 감소와 연관성이 있었다. 아울러 세신 추출물 처리에 의하여 COX-1의 발현에는 큰 변화가 없지만 COX-2는 세신 추 출물의 처리농도 의존적으로 전사 및 번역 수준에서 발 현이 모두 감소되었다. 이상의 결과에서 세신 추출물은 효과적인 암예방제로 사용될 수 있을 가능성을 제시할 수 있었으나, 후속 연구를 통하여 세신 추출물 처리에 의한 항암 효과에서의 주 성분물질에 대한 분석이 필요 할 것이다. 또한 그러한 성분물질들이 어떤 기전들을 통 하여 항암 효과를 나타내는지에 대해서도 밝혀져야 할 것이다.
Fig. 5. Down-regulation of COX-2 and iNOS expression by WEAS in A549 human lung carcinoma cells. (A) After 72 h incubation with WEAS, total RANs were isolated and reverse- transcribed. The resulting cDNAs were sujected to PCR with indicated primers, and reaction products were subjected to electrophoresis in a 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) Cells were lysed, and then cellular proteins were separated by 10%
SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with indicated antibodies. Proteins were visualized using ECL detection sys- tem. Actin was used as a loading control.
A 0 1 2 3 4 5
WEAS (mg/ml)
COX-1 COX-2
iNOS GAPDH
B 0 1 2 3 4 5
WEAS (mg/ml)
COX-1
COX-2 iNOS Actin
참 고 문 헌
1) Kim YS, Park HD, Kim EC. Taxonomic study of the genus Asarum in Korea. J Plant Taxonomic Soc Kor 8, 19-31, 1978.
2) Yook CS, Kim JG. A new species and two new forma of Asiasarum. Kor J Pharmacol 27, 342-346, 1996
3) Zhang F, Wang LX, Luo Q, Xiao HB, Liang XM, Cai SQ.
Analysis of volatile constituents of root and rhizome of Asarum heterotropoides Fr. var. mandshuricum (Maxim.) Kitag. by gas chromatography-mass spectrometry. Se Pu 20, 467-470, 2002.
4) Hashimoto K, Yanagisawa T, Okui Y, Ikeya Y, Maruno M, Fujita T. Studies on anti-allergic components in the roots of Asiasarum sieboldi. Planta Med 60, 124-127, 1994.
5) Villegas M, Vargas D, Msonthi JD, Marston A, Hostettmann K. Isolation of the antifungal components Falcarindiol and Sarisn from Heteromorpha trifoliata. Planta Medica 54, 36-37, 1998.
6) Zimmermann KC, Bonzon C, Green DR. The machinery of programmed cell death. Pharmacol Ther 92, 57-70, 2001.
7) Schultz DR, Harrington Jr WJ. Apoptosis: programmed cell death at a molecular level. Semin Arthritis Rheum 32, 345-369, 2003.
8) Howard A, Pelc SR. Synthesis of deoxyribonucleic acid in normal and irradiated cells and its relation to chromosome breakage. Heredity 6, 261, 1953.
9) Minshull J, Pines J, Golsteyn R, Standart N, Mackie S, Colman A, Blow J, Ruderman JV, Wu M, Hunt T. The role of cyclin synthesis, modification and destruction in the control of cell division. J Cell Sci 12, 77s-97s, 1989.
10) Matsushime H, Quelle DE, Shurtleff SA, Shibuya M, Sherr CJ, Kato JY. D-type cyclin-dependent kinase activity in mammalian cells. Mol Cell Biol 14, 2066-2076, 1994.
11) Koff A, Giordano A, Desai D, Yamashita K, Harper JW, Elledg S, Nishimoto T, Morgan DO, Franza BR, Roberts JM.
Formation and activation of a cyclin E-cdk2 complex during the G1 phase of the human cell cycle. Science 257, 1689-1694, 1992.
12) Brandeis M, Hunt T. The proteolysis of mitotic cyclins in mammalian cells persists from the end of mitosis until the onset of S phase. EMBO J 15, 5280-5289, 1996.
13) Elledge SJ, Harper JW. CDK inhibitors: on the threshold of checkpoints and development. Curr Opin Cell Biol 6, 847-852, 1994.
14) Morgan DO. Principles of CDK regulation. Nature 374, 131-134, 1995.
15) Harper JW, Adami GR, Wei N, Keyomarsi K, Elledge SJ.
The p21 Cdk-interacting protein Cipl is a potent inhibitor of G1 cyclin-dependent kinases. Cell 75, 805-816, 1993.
16) Taylor WR, Stark GR. Regulation of the G2/M transition by p53. Oncogene 20, 1803-1815, 2001.
17) Zeng YX, el-Deiry WS. Regulation of p21WAF1/CIP1 ex- pression by p53-independent pathways. Oncogene 12, 1557- 1564, 1996.
18) Choi YH, Lee WH, Park KY, Zhang L. p53-independent induction of p21 (WAF1/CIP1), reduction of cyclin B1 and G2/M arrest by the isoflavone genistein in human prostate carcinoma cells. Jpn J Cancer Res 91, 164-173, 2000.
19) Maga G, Hubscher U. Proliferating cell nuclear antigen (PCNA): a dancer with many partners. J Cell Sci 116, 3051- 3060, 2003.
20) Turini ME, DuBois RN. Cyclooxygenase-2: a therapeutic target. Annu Rev Med 53, 35-57, 2002.
21) Vainio H. Is COX-2 inhibition a panacea for cancer prevention?
Int J Cancer 94, 613-614, 2001.
22) Surh YJ, Chun KS, Cha HH, Han SS, Keum YS, Park KK, Lee SS. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: down-regula- tion of COX-2 and iNOS through suppression of NF-κB activation. Mutat Res 480-481, 243-268, 2001.
23) Chiarugi V, Magnelli L, Gallo O. Cox-2, iNOS and p53 as play-makers of tumor angiogenesis. Int J Mol Med 2, 715-719, 1998.
24) Thun MJ, Henley SJ, Patrono C. Nonsteroidal anti-inflam- matory drugs as anticancer agents: mechanistic, pharmacolo- gic, and clinical issues. J Natl Cancer Inst 94, 252-266, 2002.