Introduction
Recently, artificial methods of controlling repro- duction have been developed to enable the collec- tion of fish sperm at any time. However, aqua- culturists have had difficulties in obtaining sperm and eggs simultaneously in adequate ratios for insemination and management of broodstock.
Hence, many researchers have felt keenly the necessity of the development of effective tech- niques for sperm preservation, which is suitable for preservation compared to eggs.
In seedling production, artificial insemination requires a large quantity of good quality semen available. Collection and storage of good quality semen for future use may improve the conve- nience of artificial insemination and reduce stress to male broodstock caused by repeated semen sampling, which reduces semen quality (Yao et al., 2000). Cryopreservation is an effective meth- od for long-term storage of viable sperm. It has
long been used in breeding many species of animals since Polge et al. (1949) found that the addition of glycerol allowed survival of human and fowl sperm after thawing.
Since the first successful trials of cryopreser- vation of spermatozoa from marine fish were reported by Blaxter (1953) on Cluper harengus, the techniques of preservation of fish sperm have been steadily developed to obtain a fertility rate of over 80% in salmonid fish from the latter half of 1970 (Kurokura, 1983). But there is little agree- ment to results depending on the kinds and con- centrations of diluents and cryoprotectants used, and considerable discrepancy of results among many researchers.
At the present study, researcher made to opti- mize methods of cryopreservation of sperm in pointhead flounder for the cryoprotectants and freezing rates, in terms of the motility and sur- vival rate of post-thawing sperm, and observe the ultrastructural changes of sperm after freez- ing and thawing.
─
─ 213 ──
Cryopreservation of Pointhead Flounder (Cleisthenes pinetorum herzensteini ) Sperm
Kang Hee Kho and Kyoung Ho Kang
Division of Aqua Life Science, Yosu National University, Yeosu, 550-749, Korea
In the present study, attempts were made to cryopreserve sperm of the pointhead flounder (Cleisthenes pinetorum herzensteini) in liquid nitrogen to evaluate effects of various cryoprotectants and freezing rates on the motility and survival rate, and to identify the ultrastructural changes of sperm after cryopreservation. Sperm motility was significantly inhibited in 30% artificial seawater (ASW), but was restored entire- ly after 100% ASW was added again. Two cryoprotectants, dimethyl sulfoxide (DMSO) and glycerol, were added to 30% ASW to formulate the extenders at concentrations between 5% and 20% by volume for freezing. The highest post-thawed sperm motility and survival rate were obtained with 10% glycerol. Afterward, the effect of different freezing rates was examined using 10% glycerol as a cryoprotectant, and the rate of 20�C/min to -80�C showed the best result. Ultrastructural changes of the detachment of plasma membranes were observed after cryopreservation.
Key words : Pointhead flounder, Cleisthenes pinetorum herzensteini, sperm, cryop- reservation.
Materials and Methods
Fish and sperm collection
Adults pointhead flounder (Cleisthenes pineto- rum herzensteini ) were captured in the shallow sea near by Yosu city of Jeonnam province, Korea from December 2002 and transported to labora- tory. The body length, body weight and number of fish used for the experiment were 25.2±3.5 cm, 372.8±75.3 g and 7, respectively.
The semen was collected by abdominal stripp- ing and stored in polyethylene tubes on crushed ice until use.
Motility estimation and choice of diluent The motility of sperm from each male was determined immediately after collection. The percentage of sperm exhibiting rapid, vigorous and forward movement was classified under light microscope by diluting the sperm in ASW (423.0 mM NaCl, 9.0 mM KCl, 9.27 mM CaCl
2, 22.94 mM MgCl
2, 2.11 mM NaHCO
3, 10 mM HEPES- pH 7.8) in the ratio of 1:1000 (Table 1). The sam- ples with high motility were used in the follow- ing experiments.
ASW of 20, 30 and 40% were examined for selec- tion of an appropriate immobilizing solution. The fresh sperm was suspended in diluted ASW at a ratio of 1 : 10, and then the sperm motility was tested after activation with ASW.
Cryopreservation of sperm
According to results of above trials, 30% ASW was selected as the diluent for cryopreservation.
Two cryoprotectants, DMSO and glycerol, were added to 30% ASW to formulate the extenders at the concentrations between 5 and 20% by volume for freezing. The sperm was diluted 1 : 6 with the extenders. The diluted sperm was inserted into 0.5 ml plastic straws and frozen at a freezing rate of 40� C/min to -80� C in a programmable freezer (Kryosave integra, Rovers Polska, UK) after equilibration for 10 min at room tempera- ture, and subsequently plunged into liquid nitro-
gen. Twenty-four hours later, the straws were thawed in a 30� C water bath for 15 sec. On thaw- ing, the motility of frozen-thawing sperm was evaluated and subsequently the sperm was fixed by 5% glutaraldehyde in phosphate buffer (pH 7.2) for ultrastructural examination following the method described bellow. The survival rate was also estimated by an eosin-nigrosin staining technique (Bllom, 1950; Fribourgh, 1966 ). The frozen-thawing sperm was stained with a drop of 5% eosin and two drops of 10% nigrosin. After mixture smears were made, and the survival rate was determined under light microscope in terms of the percentage of unstained sperm.
To evaluate the effect of different freezing rates, 10% glycerol was used as a cryoprotectant accord- ing to the results of above experiments. Four freezing gradients were tested: (a) 40� C/min to - 80� C (b) 30� C/min to -80� C (c) 20� C/min to -80
� C (d) 10� C/min to -80� C, as above methods.
Ultrastructural changes
The fresh and frozen-thawed sperm were fixed for 2 hr at 4� C in 5% glutaraldehyde, buffered to pH 7.2 with phosphate. Samples were then washed with the same buffer. They were post- fixed for 2 hr in 1% osmium tetroxide. After dehydration through an ascending ethanol series, the samples were embedded in Spurr resin for TEM observation. Ultrathin sections were con- trasted with alcoholic uranyl acetate and lead citrate. They were then examined with TEM (JEM 1200 EX-II, 60~80 Kv, JEOL). Changes in the plasma membrane of frozen-thawed spermato- zoa were compared with the fresh sprematozoa.
Statistical analysis
Each trial of this study was carried out in trip- licate, and the results were analyzed using the Student’s t-test.
Results
Motility characteristics
On contact with artificial seawater, the sperm
Table 1. Motile index in relation to percentage of sperm with rapid, vigorous and forward movement Motility index
5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 0.5 0
Forward sperm (%) 100 90 80 65 50 30 20 15 10 5 0
were activated immediately, and reach their maximum motility at the same time. The vigor- ous forward movement of sperm is constant for about 10 sec, after which the sperm lost motility entirely.
After dilution with ASW of 20, 30 and 40%, the sperm motility was significantly inhibited, but once again restored entirely only in 30% ASW, and in 20 and 40% ASW some sperm became immotile after 100% ASW was added.
Cryoprotectant trial
The motility and survival rate of frozen-thaw- ing sperm using different crypoprotectants were showed in Fig. 1. They all declined significantly after cryopreservation. The highest post-thawed sperm motility (class 3.2±0.5) and survival rate
(70±5.4%) were obtained with 10% glycerol, and those of sperm diluted in DMSO only were less than class 1 and about 15%.
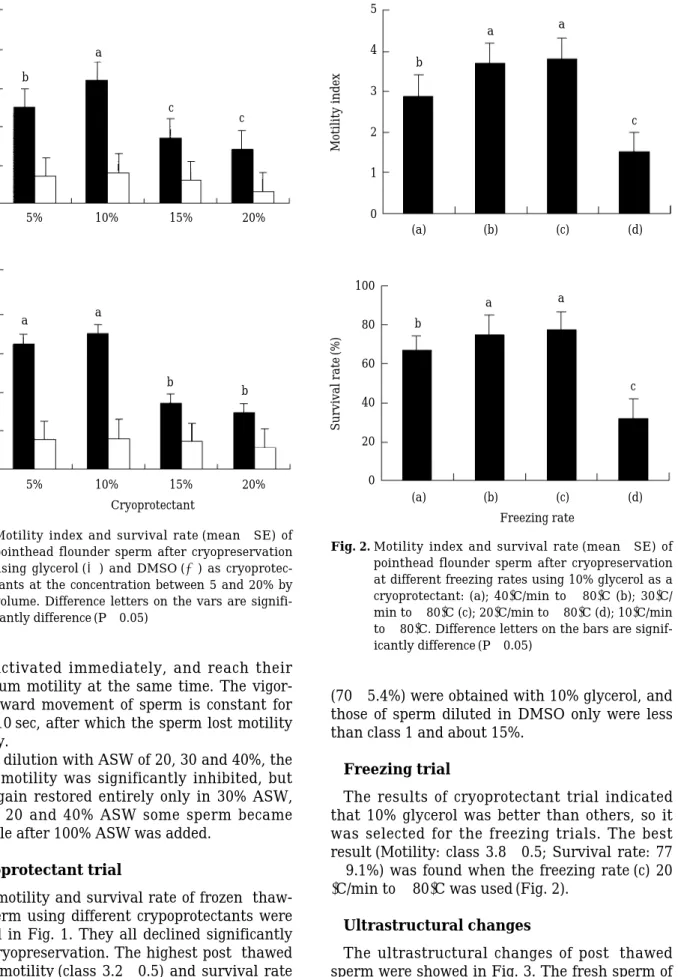
Freezing trial
The results of cryoprotectant trial indicated that 10% glycerol was better than others, so it was selected for the freezing trials. The best result (Motility: class 3.8±0.5; Survival rate: 77
±9.1%) was found when the freezing rate (c) 20
� C/min to -80� C was used (Fig. 2).
Ultrastructural changes
The ultrastructural changes of post-thawed sperm were showed in Fig. 3. The fresh sperm of
Fig. 2. Motility index and survival rate (mean±SE) of pointhead flounder sperm after cryopreservation at different freezing rates using 10% glycerol as a cryoprotectant: (a); 40�C/min to -80�C (b); 30�C/
min to -80�C (c); 20�C/min to -80�C (d); 10�C/min to -80�C. Difference letters on the bars are signif- icantly difference (P⁄0.05)
(a) (b) (c) (d)
Freezing rate 5
4
3
2
1
0 b
a
c c
a a
b b
5% 10% 15% 20%
5% 10% 15% 20%
Cryoprotectant 100
80
60
40
20
0
Motility indexSurvival rate(%)
Fig. 1. Motility index and survival rate (mean±SE) of pointhead flounder sperm after cryopreservation using glycerol (■) and DMSO (□) as cryoprotec- tants at the concentration between 5 and 20% by volume. Difference letters on the vars are signifi- cantly difference (P⁄0.05)
5
4
3
2
1
0
(a) (b) (c) (d)
b
b
a
a
a
a
c
c 100
80
60
40
20
0
Motility indexSurvival rate(%)
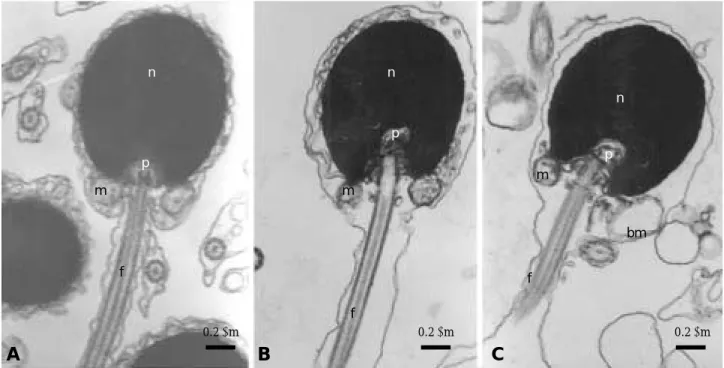
pointhead flounder had a round-shaped head, a midpiece that contained mitochondria. A plasma membrane tightly covered the head, midpiece, and tail. The chromatin was compact and homo- geneous. After cryopreservation, the chromatin did not alter, while the membrane showed some ultrastructural changes, the detachment of plasma membranes, as well as the mitochondria.
Morphological normality of the sperm in 10%
glycerol was better than that of others.
Discussion
In marine teleosts, puffers, flounders, cod, etc., osmolality isotonic to the seminal plasma sup- presses sperm motility and motility is initiated when osmolality increase (Morisawa and Mori- sawa, 1990). The immobilizing solutions, such as 1% NaCl, 0.3 M glucose, diluted seawater, and other solutions whose compositions were similar with seminal plasma, were often used as a dilu- ent in the short-term preservation and cryop- reservation of fish sperm (Blaxter, 1953; Chang et al., 1997; Fabbrocini et al., 2000; Yao et al.,
2000). The motility of pointhead flounder sperm showed short-lived, only was constant for about 10 sec after activation with ASW in this study.
Therefore, an immobilizing solution is necessary for cyropreservation as the diluent in the phases prior to freezing, so as to avoid an excessive expenditure of energy by sperm movement (Gwo, 1994). The results of motility estimation showed that 30% ASW was enough better to inhibit the sperm movement and ensure the restoration of motility after activation again. The decrease of motility of sperm diluted with 20 and 40% may be caused by the osmotic shock.
Generally, the cryopreservation of sperm results in considerable damages to cellular structures such as plasma membrane, nucleus, mitochon- dria, and flagellum, due to the formation of stress caused by heat shock, ice crystals, and excessive dehydration during freezing (Lahnsteiner et al., 1996a; Watson, 1995). Of basic importance for sperm viability is the integrity of the cellular membrane. It is well known that the plasma membrane is the primary site of cooling and warming injury (Morris, 1981) and the resistance of spermatozoa to cooling is influenced by the
Fig. 3. The ultrastructural changes in pointhead flounder spermatozoa after cryopreservation. A: Sagittal section of fresh spermatozoa showing plasma membrane, centriole, compack chromatin, mitochondria and flagellum. B: Sagittal section of spermatozoa cryopreserved using 10% glycerol showing detachment of plasma membrane. C: Sagittal section of spermatozoa cryopreserved using freezing 20% glycerol showing detached plasma membrane and the broken mitochondrion. bm: broken mitochondrion, f: flagellum, m: mitochondria, n: nucleus, p: proximal centriole.
Arrow indicates plasma membrane.
0.2µm
n n
n
m
A B C
m
m
f
f
f
bm p
p
p
0.2µm 0.2µm
biochemical composition of their membranes (Darin-Bennet and White, 1977). Cooling and warming induce phase transitions of the lipids in the sperm cell membranes, influencing the spa- tial redistribution of their components (Darin- Bennet and White, 1977; De Leeuw et al., 1990;
Quinn, 1985). Consequent to these structural changes, membranes destabilize, modify their permeability, and impair the membrane trans- porting enzymes (Holt and North, 1984); thus, spermatozoa are more liable to suffer damages during freezing and thawing (De Leeuw et al., 1993). In addition to membrane destabilization, physical damage to the plasma membranes can result from the formation of ice crystals within the cell. Cellular osmoregulation could also be compromised, resulting in swelling of the head and tail, and secondary injuries to other intracel- lular organelles can determine displacement and loss of functionality of mitochondria and outflow of nuclear chromatin (Taddei et al., 2001).
The structural damages and functional loss of spermatozoa may lead to the decline of sperm motility and survival rate (Lahnsteiner et al., 1996a), which often used as the evaluation para- meters of sperm cryopreservation. In fact, many authors have linked the percentage of motile sperm and the quality of the movement to the fertilization ability (Lahnsteiner et al., 1996a;
Lahnsteiner et al., 1996b; Lahnsteiner et al., 1996c). In the present study, the sperm motilities and survival rates were all less than those of fresh sperm, and best frozen-thawing motility (3.8±0.5) and survival rate (77±9.1%) were ob- tained by using 10% glycerol as a cryoprotectant and freezing at the rate of 20� C/min to -80� C. At the same time, the constant time of rapid, vigor- ous, and forward movement of sperm, which is also important for successful insemination of fish eggs, did not decrease compared to that of fresh sperm.
The ultrastructural analysis revealed varying degrees of aberrations, such as the detachment of plasma membranes, as well as the fracture of mitochondria, which may result in the decrease of sperm motility after cryopreservation, whereas the chromatin and flagella were well preserved in this study. The relatively higher integration of plasma membrane, and the normality of mito- chondria in sperm preserved using 10% glycerol were accordant with its higher post-thawed motility. In addition, it should be noticed that the fresh sperm of pointhead flounder also showed
abnormalities in plasma membrane, e.g. the rup- ture of membrane in the region of flagellum may be due to a combination of vulnerable or ageing cells and fixation procedures for microscopic ob- servations (Taddei et al., 2001).
References
Blaxter, J.H.S. 1953. Sperm storage and cross fer- tilization of spring and autumn spawing herring.
Nature, 172 : 1189 ~1190.
Bllom, E. 1950. A one-minute live-dead sperm stain by means of eosin -nigrosin. Fertil. Steril., 1 : 176 ~177.
Chang, Y.J., K.H. Kho and H.K. Lim. 1997. Cold storage of sperm in marbled sole (Limanda yoko-
hamae), J. Aqua., 10(4) : 381~386.
Darin-Bennet, A. and I.G. White. 1977. Influence of the cholesterol content of mammalian sper- matozoa on susceptibility to cold shock. Cryobi- ology, 24 : 466~470.
De Leeuw, F.E., A.M. De Leeuw, J.H.G. Den Daas, B. Colenbrander and A.J. Varkleij. 1993. Effects of various cryoprotective agents and membrane -stabilizing compounds on bull sperm mem- brane integrity after cooling and freezing. Cry- obiology, 30 : 32 ~44.
De Leeuw, F.E., C. Hsiao -Ching, B. Colenbrander and A.J. Varkleij. 1990. Cold -induced ultra- structural changes in bull and boar sperm plas- ma membranes. Cryobiology, 27 : 171 ~183.
Fabbrocini, A., S. Lubrano Lavadera, S. Rispoli and G. Sansone. 2000. Cryopreservation of seabream (Sparus aurata) spermatozoa. Cryobiology, 40 : 46~53.
Fribourgh, J.H. 1966. The application of a differen- tial staining method to low -temperature studies on goldfish spermatozoa. Prog. Fish. Cult., 28 : 227 ~231.
Gwo, J.C. 1994. Cryopreservation of yellowfin sea - bream (Acanthopagrus latus) spermatozoa (tele- ost, perciformes, sparides). Theriogenology, 41 : 989 ~1004.
Holt, W.V. and R.D. North. 1984. Partially irrever- sible cold induced lipid phase transition in mammalian sperm plasma membrane domains;
Freeze -fracture study, J. Exp. Zool., 230 : 473~
483.
Kurokura, H. 1983. Review-Gyorui seiekino hozon.
Suisanikusyu, 8 : 42 ~53.
Lahnsteiner, F., B. Berger, T. Wiesmann and R.
Patzner. 1996a. Changes in morphology, physi- ology, metabolism, and fertilization capacity of rainbow trout semen following cryopreservation.
Prog. Fish. Cult., 58 : 149~159.
Lahnsteiner, F., B. Berger, T. Wiesmann and R.
Patzner. 1996b. The influence of various cry-
oprotectants on semen quality of the rainbow
trout (O. mykiss) before and after cryopreserva- tion. J. Appl. Ichthyol., 112 : 99 ~106.
Lahnsteiner, F., R. Patzner and T. Wiesmann.
1996c. Semen cryopreservation of salmonid fish:
Influence of handling parameters on the post - thawing fertilization rate. Aquacult. Res., 27 : 659~671.
Morisawa, M and S. Morisawa. 1990. Acquisition and initiation of sperm motility, in: M.D. Gagnon, (Ed.), Controls of Sperm Motility: Biological and Clinical Aspects, CRC, Boston, pp. 137 ~152..
Morris, G.J. 1981. Liposomes as a model system for investigating freezing injury, in: G.J. Morris and A. Clarke (eds.), Effects of Low Temperatures on Biological Membranes, Academic, New York, pp.
214 ~262.
Polge, C.A.U. Smithand A.S. Parks. 1949. Revival of spermatozoa after vitrification and dehydra- tion at low temperatures. Nature, 164 : 666.
Quinn, P.J. 1985. A lipid -phase separation model of low temperature damage to biological mem- branes. Cryobiology, 22 : 128 ~146.
Taddei, A.R., F. Barbato, L. Abelli, S. Canese, F.
Moretti, K.J. Rana, A.M. Fausto and M. Mazzini.
2001. Is cryopreservation a homogeneous pro- cess? Ultrastructure and motility of untreated, prefreezing, and postthawed spermatozoa of
Diplodus puntazzo (Cetti). Cryobiology, 42 : 244~255.
Watson, P.E. 1995. Recent developments and con- cepts in the cryopreservation of spermatozoa and the assessment of their post-thawing func- tion. Reprod. Fertil. Dev., 7 : 871 ~891.
Yao, Z., L.W. Crim, G.F. Richardson and C.J. Emer- son. 2000. Motility, fertility andultrastructural changes of ocean pout (Macrozoarces americanus L.) sperm after cryopreservation. Aquaculture, 181 : 361 ~375.
Received : October 6, 2003 Accepted : December 7, 2003
용가자미