328
서 론
호중구는 체내로 침입하는 미생물을 탐식하고 사멸시키 는 역할을 가지는 세포로서 생체방어 작용을 하는데 우선 적으로 동원된다. 호중구, 단핵구, 및 호산구는 모두 Fas의 발현이 있으나 이들 세포 중에서 호중구만이 Fas 리간드 (FasL)를 발현시킴으로써 가장 짧은 생존기간을 가지고 있 다.(1) 정상 호중구는 자가적으로 세포사멸에 들어가거나 주위 대식세포 등의 염증 세포들에서 분비되는 FasL에 의 하여 세포사멸이 되어 대식세포에 의하여 탐식되어 소멸되 기도 한다.(2) 한편으로 FasL에 의하여 호중구의 화학주성 이 일어난다는 것은 호중구의 생존과 기능이 서로 밀접히 관계되어 있다는 것을 시사한다.(3) 호중구의 세포사멸은 세포 기능을 조절할 뿐만 아니라 단백질 분해 물질의 분비 를 차단하므로 주위 조직의 손상을 막게 된다. 그러나 말초 혈액이나 조직에서 호중구의 생존 기간은 자극제들에 의하 여 다양하게 나타난다. 이러한 호중구의 생존 기간의 조절 은 조직 손상을 유발하거나 만성적 염증 상태를 지속하는 가에 대한 중요한 결정 인자라는 것이 보고되었다.(4) 특히, 활성 산소종을 생성하는 세포의 자연 세포사멸이 조절이 되지 않을 경우 퇴행성 질환, 자가 면역성 질환, 및 종양을 일으킨다.(4)
혈관 내피세포 성장 인자(Vascular endothelial growth factor, VEGF)는 여러 종류의 암 세포들에서 혈관 생성 능력 을 가지는 헤파린 결합 단백질로 발견되었다.(5) VEGF는 혈관 내피 세포의 증식이나 이동, 혈관 평활근 세포의 이동, 및 혈관 투과도의 증가 등 여러 생리적 작용을 나타낸 다.(5,6) 홍 등(7)은 암 환자의 혈청에서 VEGF의 농도가 증 가한다고 보고하였다. VEGF의 작용은 주로 두 가지 수용체 를 통하여 일어나는데 VEGFR-1 (flt-1, fms-like tyrosine kinase 1)와 VEGFR-2(KDR, kinase domain region/flk-1, fetal liver kinase 1)가 밝혀져 있으며, 이들 중 호중구에는 VEGFR-1이 발현되어 있다.(8) 특히, VEGFR-1이 활성화된 호중구에서 세포의 화학주성이 증가되는 것으로 보고되었다.(9) 종양세 포의 배양액으로부터 분리한 VEGF를 단핵구에 처리하면 단핵구의 활성과 이동이 유도되며 VEGF에 의한 화학주성 은 VEGF-1형 수용체인 flt-1에 의해 조절된다고 하였다.(8) 한편, KDR/flt-1을 혈관 내피 세포나 다른 세포에 과발현시
혈관 내피세포 성장 인자에 의한 호중구 자연 세포사멸의 지연효과
성분도병원 외과, 1동아대학교 의과대학 생화학교실, 2서울대학교 의과대학 외과학교실
전창원․박해영1․곽종영1․김형호2
책임저자:김형호, 경기도 성남시 분당구 구미동 300 ꂕ 463-802, 분당서울대학교병원 외과 Tel: 031-787-7095, Fax: 031-787-4055 E-mail: [email protected]
접수일:2005년 6월 7일, 게재승인일:2005년 8월 1일
Delay of Spontaneous Neutrophil Apoptosis by Vascular Endothelial Growth Factor
Chang-Won Jeon, M.D., Hae-Young Park, M.S.1, Jong- Young Kwak, M.D.1 and Hyung-Ho Kim, M.D.2
Purpose: Vascular endothelial growth factor (VEGF) stimu- lates angiogenesis and vascular permeability. Tissue damage is related to angiogenesis, and induced by a delay in neutrophil apoptosis. This study was performed to inves- tigate the effect of VEGF on the spontaneous neutrophil apoptosis via the activation of VEGFR-1 and phosphorylation of the p38-MAPK pathway.
Methods: Neutrophils were prepared from 10 healthy young donors, cultured for 20 h, and the apoptosis measured by the morphological changes and flow cytometry. The VEGF receptor expression and phosphorylation of mitogen acti- vated protein kinase (MAPK) were measured using a Western blotting method.
Results: VEGF dose-dependently delayed the spontaneous neutrophil apoptosis, but this effect was blocked by pre-treatment of the cells with a VEGF receptor antagonist.
VEGF increased the phosphorylated forms of the extra- cellular stress related kinase (Erk) and p38-MAPK. However, the VEGF-induced delay in apoptosis was not affected by the Erk inhibitor, PD98059 but was affected by the p38- MAPK inhibitor, SB203580. The VEGF receptor-1, but not the VEGF receptor-2, was detected in neutrophils, but its level was reduced in cultured neutrophils.
Conclusion: VEGF delays neutrophil apoptosis through p38- MAPK activation. (J Korean Surg Soc 2005;69:328-334) Key Words: VEGF, Neutrophils, Apoptosis
중심 단어: 혈관 내피세포 성장 인자, 호중구, 세포 사멸
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Department of Surgery, St. Benedict Hospital, Busan, 1De- partment of Biochemistry, Dong-A University College of Medicine, Busan, 2Department of Surgery, Seoul National University College of Medicine, Seoul, Korea
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ 켰을 때는 세포의 증식능력이 조절된다고 하였다.(8)
호중구는 사이토카인이나 성장 인자 등에 의하여 세포의 생존 기간이 변하고 이들에 의하여 자연 세포사멸이 영향 을 받는다.(10) 호중구는 VEGF를 분비하고 혈관 생성에도 관여하는 것으로 알려져 있다.(11,12) 그러나 VEGF에 의한 호중구의 활성에 대한 연구결과는 보고된 바 있으나(8) 호 중구의 생존에 대한 VEGF의 영향은 아직 보고된 바가 없 다. VEGF 수용체를 통한 세포활성은 tyrosine kinase의 경로 를 통하여 일어나는데 호중구에서 lipopolysaccharide에 의 한 세포사멸의 지연은 tyrosine kinase의 활성을 통하여 일어 난다고 하였다.(13) 따라서 본 연구에서는 염증 및 암 조직 에서 증가하는 VEGF가 호중구의 세포사멸에 어떠한 영향 을 미치는가를 조사하고 그에 관련된 신호전달 경로를 밝 히고자 하였다.
방 법
1) 시료
Giemsa 시약은 Fluka사(Bushes, Switzerland) 제품을 사용 하였으며. 덱스트란은 Amersham Pharmacia Biotech사(Upp- sala, Sweden) 제품을 사용하였다. VEGF165는 R&D Systems Inc 회사(Minneapolis, MN, USA)로부터 구입하였다. Extra- cellular stress related kinase (Erk), phospho-ERK (p-Erk), 및 phospho-p38-mitogen activated protein kinase (p- p38-MAPK) 에 대한 특이 항체는 Cell signaling Technology사(Beverley, MA, USA)에서 구입하였고 Flt-1 항체는 Flt-1의 카르복실 발단에 대한 다클론 항체로서 Santa Cruz Biotechnology사 (Santa Cruz, CA, USA)에서 구입하였다. MEK의 억제제인 PD98059, p38MAPK 억제제인 SB203580, phosphoinositide-3 kinase (PI-3 kinase)의 억제제인 LY294002와 VEGF 수용체 의 인산화를 억제하여 VEGF작용을 봉쇄하는 길항제와 tyrosine kinase를 봉쇄하는 genistein은 Calbiochem사(San Diego, CA, USA)로부터 구입하였다. RPMI-1640 배양액은 Gibco-BRL사(Rockville, MD, USA)로부터 구입하였으며 Histopaque 및 propidium iodide (PI)와 다른 기초 시약은 Sigma사(St. Louis, MO, USA)로부터 구입하였다. VEGF 결 합 길항 펩타이드는 포항공대 채치범 교수로부터 제공받았 다.(14)
2) 호중구의 분리 및 배양
혈액(100 ml)은 0.1 M EDTA, pH 7.4와 0.9% NaCl이 들어 있는 수혈용 팩에 채혈한 후 이 혈액에 덱스트란을 넣고 조심스럽게 섞은 다음 플라스틱 실린더에서 1시간 동안 실 온 상태에서 방치하여 적혈구를 가라앉혔다. 호중구를 포 함하는 혈장 성분층을 분리하여 4oC 상태에서 850×g로 15 분 동안 원심분리하고 침전된 세포를 20 ml의 완충액 A (8.0 mM Na2PO4, 1.5 mM KH2PO4, 136 mM NaCl, 2.6 mM
KCl, 0.5 mM MgCl2, 0.6 mM CaCl2, pH 7.4)에 부유시킨 다 음 증류수를 넣어 혼재된 적혈구를 제거하였다. 임파구 분 리 용액을 세포층 밑에 넣고 850×g에서 30분간 원심분리하 여 중간층에 존재하는 임파구는 버리고 침전된 과립구 층 은 11 mM 글루코즈와 1 mg/ml 소 혈청 알부민을 첨가한 완충액 B (25 mM Hepes pH 7.4, 125 mM NaCl, 0.7 mM MgCl2, 0.5 mM EDTA)에 부유시켜 사용하였다.
3) 세포사멸의 형태적 관찰
호중구는 여러 가지 형태의 핵을 가진 세포로서 세포사 멸이 일어날 경우 핵 모양의 변화가 일어난 세포의 수로서 그 정도를 측정하였다.(15) Cytospin에서 500×rpm으로 1분 간 원심분리한 다음 메탄올로 고정시키고 May-Grunwald Giemsa 염색액으로 염색하였다. 호중구의 세포사멸 정도는 400배 현미경 시야에서 최소한 300개의 세포 중에서 특이 적인 변화가 나타난 세포를 확인하여 %로 표시하였다.
4) Flow cytometry 측정
세포사멸의 양적 측정은 phosphatidylserine의 세포막으로 의 노출과 PI 염색 정도를 apoptosis detection kit (Oncogene Research Products, Boston, MA, USA)를 사용하여 annexin V-fluorescein isothiocyanate (FITC)결합으로 측정하였다. 호 중구(1×106)를 채취하여 인산완충액으로 1회 세척한 후, annexin V-FITC와 PI로 15분간 배양하였다. 반응시킨 세포 의 일부(1×104)를 flow cytometry를 이용하여 측정하였 다.(15)
5) Western blot
전기영동 후 Western blot법을 사용하여 각각의 항체를 이 용하여 발현 정도를 측정하였다. 호중구를 2.5% mercaptoe- thanol과 1.2% sodium dodecyl sulfate를 넣고 100oC에서 5분 간 가열하여 처리한 다음 7.5% 혹은 10% polyacrylamide 겔 에서 전기영동한 후 겔의 단백질을 nitrocellulose 막으로 전 이시켰다. Nitrocellulose 막을 10 mM Tris HCl, 0.15 M NaCl, 0.1% NaN3로 구성된 완충액에 5% 탈지분유를 포함하는 완 충액으로서 2시간 처리하며 1차 항체는 1:1000으로 희석 하여 실온에서 하룻밤 동안 반응시켰다. Horse-radish perox- idase가 결합된 이차 항체를 실온에서 1시간 동안 반응시켰 다. Triton X-100이 들어 있지 않는 Tris 용액으로 세척하고 ECL chemiluminescence 발색 방법으로 전개시켰다.
6) 통계 처리
세포사멸의 결과는 평균±표준편차(mean±SD)로 나타내 었다. 통계학적 유의성은 자료들의 분포를 확인하고 Stu- dent's t-test를 이용하여 검정하였고 P값이 0.05 이하이면 유 의성이 있는 것으로 판정하였다.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
결 과
1) 호중구의 자연 세포사멸에 대한 VEGF의 영향 호중구의 형태적 관찰은 Fig. 1의 A에서 나타낸 바와 같 이 호중구 세포사멸의 현상을 가장 특징적이고 간단하게 확증할 수 있는 방법이다. 이 방법은 호중구의 세포 특성상 다른 어떠한 세포보다도 세포사멸 동안 핵이 농축되고 그 후 분획으로 된 것이 잘 나타나고 세포질에 액포가 나타나 는 것을 쉽게 관찰할 수 있다. 호중구를 분리하여 배양 상태 에서 20시간을 방치하면 50∼60%의 세포는 핵의 응축 등 세포사멸의 형태를 보이나 20 ng/ml의 VEGF를 투여하였을 때 형태적으로 정상 세포가 대부분 유지되었다(20±6%).
Fig. 1의 B에서는 이러한 세포사멸을 annexin V와 PI염색으 로 관찰하였을 때도 annexin V와 PI에 염색되는 세포의 비 율이 현저히 감소하는 것으로 나타났다(65±5% vs 29±6%, P<0.01). 이러한 결과는 VEGF에 의하여 호중구의 자연 세 포사멸이 억제된다는 것을 알 수 있다. VEGF의 세포사멸에 대한 효능을 농도별로 측정하였을 때 20 ng/ml의 농도에서 현저히 세포사멸이 억제되는 것으로 나타났다(Fig. 2).
Fig. 1. Delay of spontaneous apop- tosis of neutrophils by VEGF. (A) Cytospin prepa- rations of neutrophils after incubation in vitro for 20 h were stained with Giemsa solution. Solid arrows indi- cate apoptotic cells. Giemsa stain, ×400. (B) Neutrophils were cultured for 20 h and cell death was assessed by annexin V-FITC and PI stai- ning. Results are represent- atives of three independent experiments.
PI FL3-H FL3-H
Annexin V-FITC
20 h/None 20 h/NEGF
104 104
103 103
102 102
101 101
100 100
100 101 102 100 101
FL1-H FL1-H
102
103 103
6.98% 10.20%
58.31% 19.41%
1.01% 0.70%
104 104
A
B
Fig. 2. Dose-dependent anti-apoptotic effect of VEGF on spon- taneous apoptosis of neutrophils. Neutrophils were incu- bated with indicated concentrations of VEGF for 20 h and apoptotic cells were identified by their morphology under light microscopy and counted. Results are representatives of three independent experiments. *P<0.001 vs untreated cells.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
2) VEGF 수용체 길항제의 영향
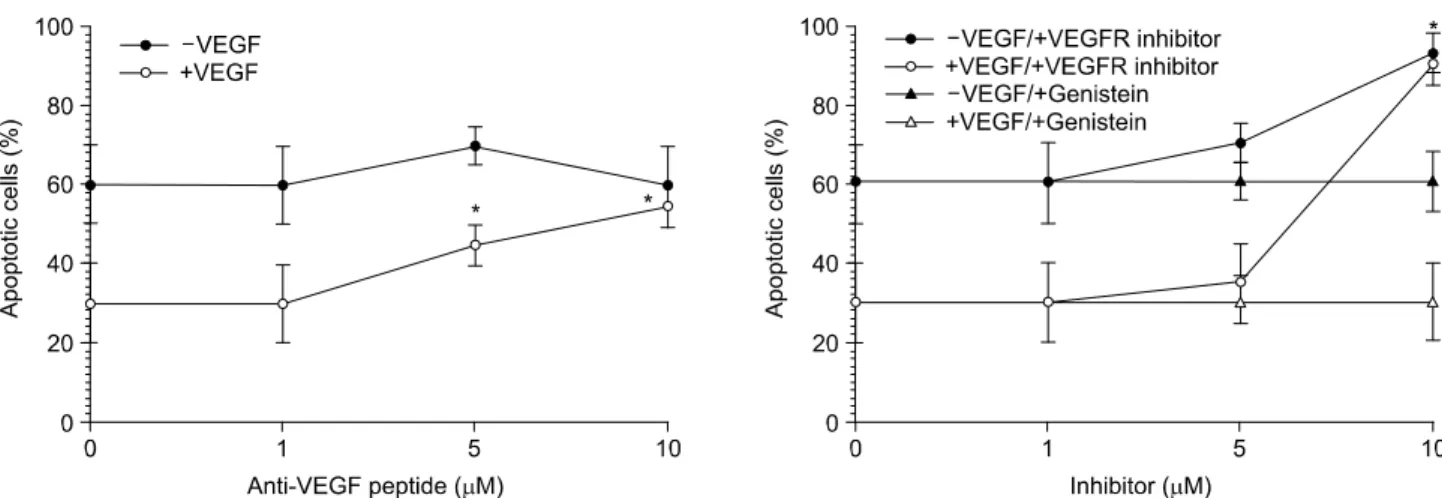
VEGF의 세포사멸 지연 효과가 호중구의 VEGF 수용체 특 이성으로 일어나는가를 보기 위하여 VEGF가 수용체에 결 합하는 것을 억제하는 펩타이드를 먼저 투여한 후 VEGF의 효능을 조사하였다. Fig. 3A에 나타낸 바와 같이 배 등(14) 이 개발한 VEGF 길항 헥사 펩타이드는 10μM 농도에서 VEGF의 세포사멸 지연 효과를 억제하였으며(56±5%, P<
0.001), Hennequin 등(16)이 개발한 VEGF 수용체의 길항제 도 10μM 농도에서 VEGF에 의한 호중구의 세포사멸 지연 을 방지하였다(92±5%, P<0.001)(Fig. 3B). 이러한 결과는 VEGF가 호중구의 VEGF 수용체를 통하여 일어난다는 것 을 제시한다. 한편, VEGF의 수용체의 길항제 단독은 자연 세포사멸을 촉진하는 것으로 보아 자연 세포사멸에 호중구 의 VEGF 수용체의 활성이 지연 효과를 가지는 것으로 보 인다. 그러나, VEGF에 대한 세포사멸 지연은 tyrosine kinase 억제제인 genistein에 의해서는 억제되지 않았으나 VEGF 수 용체의 tyrosine kinase 활성 억제제에 의하여 억제되었다 (Fig. 3B). 따라서, VEGF-1형의 수용체의 활성이 세포사멸 지연과 연관이 있으며 tyosine기의 인산화는 큰 영향이 없는 것으로 추측된다.
3) VEGF에 의한 MAPK의 활성과 세포사멸과의 관계 VEGF의 수용체를 활성화시킨 후 호중구 내 신호 전달 인자 중 MAPK의 활성을 측정하였다. Erk의 인산화는 일반 적으로 자극 후 30분까지의 빠른 시간에 일어나므로 0.5, 1, 2, 5, 및 10분 동안 호중구를 VEGF로 처리한 후 실험을 진 행하였다.(17) Fig. 4에 나타난 바와 같이 VEGF에 의한 Erk 의 인산화는 0.5분부터 증가하기 시작해서 1분에 최고에 이 르고 2분부터는 급격히 소실되었다. 또한 VEGF 첨가에 의 한 p38-MAPK의 인산화는 0.5, 1분에 최고에 이르나 2분부
터 감소하였다. 이는 VEGF 투여 1분 후 최고에 이른 Erk의 인산화에 의해 p38-MAPK의 활성이 봉쇄된 것이라 생각되 나 VEGF를 첨가하지 않은 대조군과 비슷한 수준의 인산화 가 유지되었으며 감소하지는 않았다. 이러한 사실은 VEGF 에 의하여 Erk와 p38-MAPK가 활성화되어 세포사멸 지연을 나타낼 것으로 추측할 수 있다. 그러나 Erk 활성의 억제제 인 PD98059로 전처리를 하였을 때 VEGF의 세포사멸 지연 효과는 억제되지 않았으나 p38-MAPK의 억제제인 SB203580 에 의하여 세포사멸의 비율이 자연 세포사멸 정도로 증가 하였다(Fig. 5). 따라서 이러한 결과로부터 VEGF첨가에 의 하여 Erk, p38-MAPK가 모두 인산화되지만 이들 kinase에 대한 억제제를 전처리하였을 때 p38-MAPK에 의하여 세포 사멸의 지연이 나타나는 것으로 생각된다. 호중구의 세포 사멸을 지연하는 데 중요한 역할을 하는 신호전달인자인 PI-3 kinase에 대한 억제제인 LY294002을 전처리한 경우 (18,19) VEGF에 의한 세포사멸의 지연은 봉쇄되지 못하였 Fig. 3. Blocking of VEGF effect on neutrophil apoptosis by VEGF receptor antagonist. Neutrophils were pretreated with or without
anti-VEGF peptide (A) or VEGFR antagonist or genistein (B) for 1 h and further incubated for 20 h in the presence of 20 ng/ml VEGF. Results are representatives of three independent experiments. *P<0.001 vs untreated cells.
Fig. 4. Erk and p38-MAPK activation by VEGF in neutrophils.
Neutrophils (1×107) were treated for indicated times with 20 ng/ml VEGF at 37oC. A Western blot of proteins from cell lysates was probed with an antibody that recognizes phosphorylated Erk or p38-MAPK. Results are represen- tatives of three independent experiments.
0.5 1 2
-VEGF +VEGF
5 10 0.5 1 2 5 10 (min)
p-p38-MARK p-Erk Erk
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
는데 이러한 결과는 VEGF의 세포사멸 지연 효과는 PI-3 kinase와는 관계없이 일어난다는 것을 제시한다.
4) 호중구에서 VEGF 수용체의 발현
호중구에서 VEGF 수용체 중 어떠한 형이 발현되는가를 Western blot법으로 측정하였다. VEGF 수용체 1형인 flt-1은 혈액으로부터 분리한 호중구에서 발현되었으나 배양 후 그 발현 양은 현저히 감소하여 20시간 후에는 거의 측정되지 않았다. 이는 호중구가 시험과 내 배양 상태에서 24시간 후 약 50∼60%의 자연 세포사멸을 일으키는데 세포사멸된 호 중구에서 flt-1 단백질 양이 감소된 것으로 추측된다. 이에 비교하여 VEGF 수용체 2형인 flk-1은 호중구에서 측정되지 않았다(Fig. 6).
고 찰
호중구는 생체 내 염증이 일어나는 경우 가장 먼저 동원 되는 탐식 세포로서 다량의 활성 산소종을 생성하여 세균 등을 사멸시키지만 이러한 생성물이 조직의 손상을 동반하 기도 한다. 따라서 활성화된 호중구의 기능을 억제 조절하 는 것이 염증 반응을 종결시키기 위하여 매우 중요한데 최 근에는 활성화된 기능을 억제하는 것과 더불어 이들의 세 포사멸이 염증의 종결을 위하여 필수적이며 더욱 중요하다 고 제안되고 있다. 혈관 신생은 염증이나 상처의 치유에서 중요한 역할을 한다. 본 연구 결과에서와 같이 VEGF가 증 가하는 경우 호중구의 세포사멸이 지연되고 호중구의 기능 이 유지된다는 것은 종양 조직이나 염증 부위에서 조직에 대한 VEGF의 영향이 호중구를 통하여 일어날 가능성을 제 시하고 있다. 또한 호중구는 자극을 받는 경우 VEGF의 생 성 및 분비가 증가한다고 알려져 있다.(11,20)
호중구는 완전히 분화된 세포로 세포 분열이 일어나지 않으므로 세포 주기에 영향을 주어서 세포사멸을 결정짓는 조절 인자들보다는 세포 내 신호 전달 경로에 의하여 결정 되는 것임을 시사하고 있다. 아직 호중구의 세포사멸의 지 연을 결정한다고 알려진 인자로는 cAMP-dependent protein kinase (PKA), PI-3 kinase, 및 MAPK 등의 신호 전달 인자가 관여할 것으로 제시되고 있다.(18) 그러나 VEGF에 의한 세 포사멸의 지연은 Erk의 억제제에 의하여 영향을 받지 않았 으나 p38-MAPK의 억제제에 의하여 세포사멸이 증가한 것 으로 보아 p38-MAPK가 VEGF의 작용을 매개하는 것으로 보인다. VEGF는 호중구를 활성화하므로 VEGF의 자극을 받은 호중구가 세포사멸에 영향을 주는 여러 사이토카인이 나 인자들을 분비하여 호중구의 생존이 길어질 가능성도 배제할 수 없을 것이다.
VEGF 수용체에 결합하는 인자로서 phospholipase Cγ, PI-3 kinase, P120 GAP 뿐만 아니라 세포 골격체에 관여하는 focal adhesion kinase와 paxillin도 포함된다.(21,22) 하지만, 호중구에서 VEGF 수용체 자극 후 어떠한 경로로 활성화되 고 세포사멸이 지연되는가는 연구된 바가 없다. GM-CSF를 처리한 호중구는 세포의 생존이 길어졌는데 이에는 tyrosine 기의 인산화가 관여한다고 하였다.(23) 그러나 호중구에서 tyrosine 인산화는 세포사멸에 대하여 그 자극되는 정도에 따라 증가와 억제의 현상 두 가지를 다 나타낸다는 결과가 보고되었다.(23) VEGF에 자극을 받은 세포는 VEGF 수용체 가 가지고 있는 tyrosine kinase의 활성에 의하여 수용체의 자가 인산화가 일어나고 그 아래 단계의 결합 단백질과 결 합한다.(24) 본 연구에서 tyrosine kinase 억제제인 genistein 은 VEGF에 대한 세포사멸 지연을 억제하지 못하였으나 Fig. 5. Effects of kinase inhibitors on VEGF-induced delay of
neutrophil apoptosis. Neutrophils were preincubated with 10μM inhibitors for 1 h and further incubated with or without VEGF. Results are representatives of three independent experiments. *P<0.01 vs None.
Fig. 6. Expression of VEGF receptors on neutrophils. Neutrophils were cultured for 6 and 20 h. Cell lysates were electro- phoresed on 7.5% SDS-PAGE and VEGFR proteins were detected using anti-Flt-1 antibody. Results are represen- tatives of three independent experiments.
Fresh 6 h
1 Ab : Flt-1 20 h
205 kDa
97.4 kDa 116 kDa
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ VEGF 수용체의 tyrosine kinase 활성 억제제에 의하여 억제
되었다. 따라서, VEGF-1형의 수용체의 활성이 세포사멸지 연과 연관이 있으며 tyosine기의 인산화는 큰 영향이 없는 것으로 추측된다.
VEGF를 포함하는 사이토카인 등에 의하여 유도된 세포 활성화 과정을 억제하는 것과 함께 세포사멸을 유도하는 것은 더 이상의 조직 손상을 막는 새로운 치료 방법으로 제기되고 있다. 따라서 호중구의 세포사멸의 조절에 가장 큰 영향을 주는 사이토카인이 세포사멸에 어떠한 영향을 주는가를 밝히는 것은 앞으로 매우 중요한 과제일 것이다.
결 론
VEGF는 혈관의 생성이나 투과에 영향을 주는 성장 인자 이다. 혈관 생성은 조직 손상과 연관이 있으며 호중구의 세 포사멸의 지연은 조직 손상을 유발하는 것으로 알려져 있 다. VEGF를 투여한 호중구에서 농도 의존형으로 호중구 세 포사멸이 지연되었다. VEGF를 투여하기 전 VEGF 수용체 의 길항제를 처리하였을 때 VEGF의 세포사멸 지연 효능은 억제되었다. 호중구에서 VEGF 수용체의 발현을 조사하였 을 때 VEGF 수용체 1형이 발현되었으나 배양 후 발현이 감소하였으며 VEGF 수용체 2형은 검출되지 않았다.
이러한 결과들을 고려하면 혈관 신생에 관여하는 VEGF 는 호중구의 세포사멸을 지연시키며 이는 VEGF에 의해 세 포이동이 일어나는 단핵구와 같이 VEGF 수용체 1형인 flt-1 에 의해 조절된다. VEGF에 의한 세포사멸 지연이 p38- MAPK의 인산화경로를 통해 일어나며 PI-3 kinase 경로와는 독립적이라는 결과를 통해 호중구에서 VEGF 수용체의 자 극 후 어떠한 경로로 활성화되고 세포사멸이 지연되는가에 대한 세포 내 신호 전달 경로에 대한 이해의 토대를 마련하 였다. 또한 노화된 호중구에서 VEGF 수용체의 발현이 감소 되는 것으로 보아 혈관신생이 일어나는 부위에서 호중구의 생존이 어떠한 영향을 줄 것으로 생각된다.
REFERENCES
1) Liles WC, Kiener PA, Ledbetter JA, Aruffo A, Klebanoff SJ.
Differential expression of Fas (CD95) and Fas ligand on normal human phagocytes: implications for the regulation of apoptosis in neutrophils. J Exp Med 1996;184:429-40.
2) Brown SB, Savill J. Phagocytosis triggers macrophage release of Fas ligand and induces apoptosis of bystander leukocytes.
J Immunol 1999;162:480-5.
3) Seino K, Iwabuchi K, Kayagaki N, Miyata R, Nagaoka I, Matsuzawa A, et al. Chemotactic activity of soluble Fas ligand against phagocytes. J Immunol 1998;161:4484-8.
4) Ward C, Dransfield I, Chilvers ER, Haslett I, Rossi AG. Phar- macological manipulation of granulocyte apoptosis: potential
therapeutic targets. Trends Pharmacol Sci 1999;20: 503-9.
5) Leung DW, Cachianes G, Kuang WJ, Goeddel DV, Ferrara N. Vascular endothelial growth factor is a secreted angiogenic mitogen. Science 1989;246:1306-9.
6) Keck PJ, Hauser SD, Krivi G, Sanzo K, Warren T, Feder J, et al. Vascular permeability factor, an endothelial cell mitogen related to PDGF. Science 1989;246:1309-12.
7) Hong SW, Whang DH, Pail IW, Lee HS. Serum vascular endothelial growth factor (VEGF) and microvessel tumor invasion in hepatocellular carcinoma. J Korean Surg Soc 2003;
64:224-8.
8) Barleon B, Sozzani S, Zhou D, Weich HA, Mantovani A, Marme D. Migration of human monocytes in response to vascular endothelial growth factor (VEGF) is mediated via the VEGF receptor flt-1. Blood 1996;87:3336-43.
9) Ancelin M, Chollet-Martin S, Herve MA, Legrand C, El Benna J, Perrot-Applanat M. Vascular endothelial growth factor VEGF189 induces human neutrophil chemotaxis in extra- vascular tissue via an autocrine amplification mechanism. Lab Invest 2004;84:502-12.
10) Maianski NA, Maianski AN, Kuijpers TW, Roos D. Apoptosis of neutrophils. Acta Haematol 2004;111:56-66.
11) Taichman NS, Young S, Cruchley AT, Taylor P, Paleolog E.
Human neutrophils secrete vascular endothelial growth factor.
J Leukoc Biol 1997;62:397-400.
12) Schruefer R, Lutze N, Schymeinsky J, Walzog B. Human neutrophils promote angiogenesis by a paracrine feedforward mechanism involving endothelial interleukin-8. Am J Physiol Heart Circ Physiol 2005;288:H1186-92.
13) Sweeney JF, Nguyen PK, Omann GM, Hinshaw DB.
Lipopolysaccharide protects polymorphonuclear leukocytes from apoptosis via tyrosine phosphorylation-dependent signal transduction pathways. J Surg Res 1998;74:64-70.
14) Bae DG, Gho YS, Yoon WH, Chae CB. Arginine-rich anti-vascular endothelial growth factor peptides inhibit tumor growth and metastasis by blocking angiogenesis. J Biol Chem 2000;275:13588-96.
15) Park MA, Lee MJ, Lee SH, Jung DK, Kwak JY.
Anti-apoptotic role of phospholipase D in spontaneous and delayed apoptosis of human neutrophils. FEBS Lett 2002;
519:45-9.
16) Hennequin LF, Thomas AP, Johnstone C, Stokes ES, Lon JJ, Ogilvie DJ, et al. Design and structure-activity relationship of a new class of potent VEGF receptor tyrosine kinase inhibi- tors. J Med Chem 1999;42:5369-89.
17) Cha MS, Lee MJ, Je GH, Kwak JY. Endogenous production of nitric oxide by vascular endothelial growth factor down- regulates proliferation of choriocarcinoma cells. Biochem Biophys Res Commun 2001;282:1061-6.
18) Akgul C, Moulding DA, Edwards SW. Molecular control of neutrophil apoptosis. FEBS Lett 2001;487:318-22.
19) Kotone-Miyahara Y, Yamashita K, Lee KK. Short-term delay
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ of Fas-stimulated apoptosis by GM-CSF as a result of tem-
porary suppression of FADD recuitment in neutrophils:
evidence implicating phosphatidylinositol 3-kinase and MEK1- ERK1/2 pathways downstream of classical protein kinase C.
J Leukoc Biol 2004;76:1047-56.
20) Gaudry M, Bregerie O, Andrieu V, El Benna J, Pocidalo MA, Hakim J. Intracellular pool of vascular endothelial growth factor in human neutrophils. Blood 1997;90:4153-61.
21) Ferrara N, Davis-Smyth T. The biology of vascular endothelial growth factor. Endocr Rev 1997;18:4-25.
22) Le Boeuf F, Houle F, Hout J. Regulation of vascular endothelial growth factor receptor 2-mediated phosphorylation of focal adhesion kinase by heat shock protein 90 and Src kinase activity. J Biol Chem 2004;279:39175-85.
23) Yousefi, S, Green, DR, Blaser, K, Simon, HU. Protein-tyrosine phosphorylation regulates apoptosis in human eosinophils and neutrophils. Pro Natl Acad Sci USA 1994;91:10868-72.
24) Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Vascular endothelial growth factor (VEGF) and its receptors. FASEB J 1999;13:9-22.