생체시계 단백질의 세포 내 위치를 조절하는
인산화 효소 발굴
아 주 대 학 교 대 학 원
의생명과학과/신경과학전공
인산화 효소 발굴
지도교수 김 은 영
이 논문을 이학 석사학위 논문으로 제출함.
2015년 2월
아 주 대 학 교 대 학 원
의생명과학과/신경과학전공
박 나 영
박나영의 이학 석사학위 논문을 인준함.
심사위원장 김 은 영 인
심 사 위 원 백 은 주 인
심 사 위 원 조 은 혜 인
아 주 대 학 교 대 학 원
2014년 12월 04일
i
생체시계의 분자 기전은 전사인자인 CLOCK과 BMAL1 단백질이 전사를 촉 진하고 그 결과 발현된 PERIOD와 CRYPTOCHROME이 CLOCK/BMAL1의 활성을 억제함으로써 생체시계 유전자 및 생체시계의 조절을 받는 유전자의 발현을 하 루를 주기로 oscillation 하도록 하는 것이며, 생체시계는 이를 통해 생명체의 행 동 및 생리작용들이 생체리듬을 나타내도록 관장한다. 그런데 생체시계가 24시간 의 주기를 가지고 정상적으로 작동하기 위해서는 전사 억제기능을 하는 PERIOD (PER)와 CRYPTOCHROME (CRY) 단백질의 양 및 핵으로 이동하는 시간을 조절
하는 것이 필수적이다. 이들 단백질의 양, 활성 및 세포 내 위치가 번역 후 조절 (post-translational regulation), 특히 인산화에 의해 조절될 수 있음이 보고되어 있다. 본 연구에서는 mCRY1과 mPER2의 세포 내 위치를 조절하는 인산화 효소를 발 굴하고자 유전체 수준에서 스크리닝을 진행하였다. 먼저 스크리닝에 사용할 수 있도록 인간 인산화 효소 유전체 라이브러리를 구축하였다. 이를 위하여 인간 인산화 효소 533개에 대하여 포유류 발현 벡터에 형광 단백질 (red fluorescence protein)로 표지하는 클로닝을 진행하였고, 궁극적으 로 385개의 인산화 효소를 포함하는 RFP 표지 인간 인산화 효소 유전체 라이브 러리를 구축하였다.
ii
스크리닝했다. 그 중 hVRK3 (vaccinia related kinase 3)와 hUCKL1 (uridine-cytidine kinase 1-like 1)이 mCRY1 단백질의 세포 내 위치를 변화시켰다. mCRY1은 NIH
3T3과 HEK 293T 세포에서 핵에 위치한다. 그러나 hVRK3와 함께 발현함으로써
핵과 세포질에 위치했다. 또한 hVRK3는 mCRY1뿐 아니라 mCRY2, mPER2, BMAL1과 같은 다른 생체시계 단백질의 세포 내 위치에도 변화를 주었다.
hVRK3와 달리, hUCKL1은 mCRY1에만 특이적으로 핵이 아닌 세포질에 머물게
했다.
EGFP가 표지된 mPER2 단백질은 HEK 293T 세포에서 스크리닝을 진행하였
다. 현재 82개의 인산화 효소를 스크리닝 하였으며 hPRKRA (interferon-inducible double stranded RNA-dependent protein kinase activator A)가 mPER2 단백질 특이적으
로 세포 내 위치를 변화시켰다. mPER2는 NIH 3T3와 HEK 293T 세포에서 핵에 약 75% 존재한다. 그러나 hPRKRA를 함께 발현시키면 mPER2의 위치가 세포질에 많이 있는 것을 보았다. 흥미롭게도 HEK 293T 세포에서 발현시켰을 때 hPRKRA 는 mPER2 단백질의 인산화를 증가시켰고 mPER2 단백질의 양도 증가시켰다. hPRKRA 이 외에 mPER2의 세포 내 위치를 변화 시키는 새로운 인산화 효소
hNEK2 (NIMA -never in mitosis gene A- related kinase 2), hCKMT1B (creatine kinase, mitochondrial 1B), hETNK1 (ethanolamine kinase 1), hMST1R (macrophage stimulating 1 receptor)을 발굴했다.
iii
iv 차례 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ ⅳ 그림 차례 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ ⅶ 표 차례 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ ⅸ Ⅰ. 서론 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 1 A. 생체시계의 분자 생물학적 기전 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 1 B. 인산화에 의한 생체시계 조절 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 2 C. 핵 이동 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 5 D. 연구의 필요성 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 5 Ⅱ. 재료 및 방법 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 6
A. Plasmids and cloning ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 6
B. Establishment of human kinome expression library ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 9
C. Cell culture and transfection ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 9
D. Fluorescence microscopy ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 11
E. Quantitative real-time RT-PCR ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 12
F. Cell lysis ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 12
G. Western blotting ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 13
v
Ⅲ. 결과 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 17
A. 인간 인산화 효소 라이브러리 구축 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 17
B. 인간 인산화 효소 스크리닝 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 20
C. VRK3 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 27
1. hVRK3는 mCRY1/2, mPER2, BMAL1 단백질의 세포 내 위치를 조절한
다 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 27 2. hVRK1과 hVRK2는 mCRY1의 세포 내 위치를 바꾸지 않는다 ∙∙∙∙∙∙∙∙∙∙ 30 3. hVRK3는 mSTAT3의 세포 내 위치에는 영향을 미치지 않는다 ∙∙∙∙∙∙∙∙∙ 32 4. hVRK3는 핵 안에서 작용하여 생체시계 단백질 -mCRY1, mPER2, BMAL1- 을 세포질에 머물게 한다 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 34 D. PRKRA ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 38
1. hPRKRA는 mPER2 특이적으로 세포 내 위치를 변화시키고, mCRY1과
mPER2, BMAL1의 양을 증가시킨다 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 38
2. mPER2와 hPRKRA는 결합하며 hPRKRA는 mPER2 단백질의 인산화
를 증가시킨다 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 41 3. CG6866 -prkra Drosophila homolog- knockdown 초파리는 rhythmicity가
vi
Ⅴ. 결론 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 53
참고문헌 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 55 ABSTRACT ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 59
vii
Figure 1 Schematic diagram of establishment of fluorescent peptide tagged human kinome expression library ··· 19
Figure 2 Kinases altering subcellular localization of mCRY1 in NIH 3T3 cells ·· 23
Figure 3 Quantification of subcellular localization of mPER2 in HEK 293T cells 26
Figure 4 Co-transfection of hvrk3 changes the subcellular localization of mCRY1/2, mPER2 or BMAL1 in HEK 293T cells ··· 28
Figure 5 hVRK3 increases subcellular localization of nucleus and cytoplasm more than nucleus in co-expression with mCRY1/2, mPER2, BMAL1 ··· 29
Figure 6 Co-transfection of hvrk1/2 did not alter the nuclear localization of mCRY1 ··· 31
Figure 7 Co-transfection of hvrk3 did not alter the subcellular localization of mSTAT3 ··· 33
Figure 8 hVRK3-NLS mutant is present predominantly in the cytoplasm in HEK 293T cells ··· 36
Figure 9 The extent of alterations in subcellular localization of mCRY1, mPER2, and BMAL1 was attenuated in the case of co-expressing NLS mutant version of hVRK3 in HEK 293T cells ··· 37
Figure 10 Co-expression of hPRKRA altered subcellular localization of mPER2 in HEK 293T cells ··· 39
viii
flies ··· 45
Figure 14 hUCKL1 alters the subcellular localization of mCRY1 ··· 47
Figure 15 Co-expression of hNEK2, hCKMT1B, hETNK1, or hMST1R altered the subcellular localization of mPER2 in HEK 293T cells ··· 48
ix
Table 1 Kinases which regulate circadian clock in mammals ··· 4
Table 2 Primer list for cloning ··· 8
Table 3 List of screened kinases ··· 22
1
Ⅰ
서론
A. 생체시계의 분자 생물학적 기전 살아있는 모든 생명체들은 하루 동안의 내제되어있는 정교한 생체시계 (circadian clock)를 지닌다. 이로 인해, 식물에서의 광합성부터 인간에서의 수면-각 성까지 거의 모든 생리작용들이 24시간 주기를 갖는 생체리듬을 나타낸다. 이러 한 생체시계는 환경의 변화로부터 적응되고 보존되었다 (Mohawk et al., 2012). 포유류에서 생체시계를 조절하는 핵심 중추는 시상하부의 SCN (suprachiasmatic nucleus)이다 (Okamura, 2007; Cuninkova and Brown, 2008). 빛이나 온도, 음식과 같은 자극 들이 SCN을 통해 인지되면 SCN은 여러 기관과 조직으로 신호를 전달하고 여러 조직과 기관의 일주기 리듬을 관장한다. 포유류에서 생체 시계의 분자적 기전은 세포의 자율성을 가지며 자기조절의 음성 되먹임 전사활 동 (auto-regulatory negative feedback loop)에서 유발된다 (Lowery and Takahashi 2004, Takahachi et al., 2008). 일반적으로 두 개의 되먹임 고리 (feedback loop)가 서로 맞
물려 유전자의 발현을 조절하며, 핵심 생체시계 유전자는 clock, bmal1, period (per1, per2, per3), cryptochrome (cry1, cry2)으로 구성된다. CLOCK 단백질은 BAML1 단백질과 상호작용하여 period와 cryptochome과 같은 여러 유전자들의 전사를 유 도한다 (Reppert and Weaver, 2002). 이로 인해 축적된 PERIOD와 CRYPTOCHROME
2
단백질은 인산화 (phosphorylation)가 일어나고 이량체를 이루어 핵 안으로 이동 (nuclear translocation)한다. 이들은 CLOCK과 BMAL1에 작용하여 전사를 억제시키
며 이러한 되먹임 고리가 약 24시간을 주기로 일어난다.
생체시계가 제대로 작동되지 않으면 암, 수면장애, 우울증, 노화, 신경퇴행 성 질환, 대사 증후군, 약물중독과 같은 많은 질병을 유발한다는 보고가 있다 (Fu et al., 2002; Morikawa et al., 2005; Roybal et al., 2007; Liu Z and Chu G, 2013). 따라
서 생체시계의 작동 기전을 연구하는 것은 다양한 생리작용의 시간에 따른 조절 기전을 이해하고 나아가 위와 같은 여러 가지 질병을 예방하고 치료할 수 있다. B. 인산화에 의한 생체시계 조절 생체시계가 24시간으로 유지되기 위해서는, 생체시계 단백질들의 양과 활성, 위치 변화뿐 아니라 인산화와 같은 번역 후 조절 (post-translational regulation)이 중요하다. 이전 연구에서 생체시계 단백질의 세포 내 분포를 조절하는 인산화 효 소들이 알려져 있다. 예를 들어 PER1은 casein kinase Ⅰε에 의해 인산화되고 이어 ubiquitin-proteasome pathway를 통해 분해된다 (Takano et al., 2004; Virshup et al., 2007). 또한 casein kinase Ⅱ와 GSK3β는 PER2를 인산화시키고 세포 내 위치를 조절한다 (Iitaka et al., 2005; Maier et al., 2009). 현재까지 포유류에서 생체시계의 작동에 필수
적인 것으로 알려진 인산화 효소는 CKⅠ α/ε/δ, CKⅡ, GSK3β, AMPK 그리고 CDK5 5개다 (Hirota et al., 2010; Reischl and Kramer, 2011; Kwak et al., 2013) (Table 1).
3
에도 생체시계 단백질 조절에 관여하는 더 많은 인산화 효소가 존재할 것이라 본다. 본 연구에서는 인간 인산화 효소 라이브러리를 구축하고 생체시계 단백질 의 세포 내 위치를 변화 시키는 인산화 효소를 발굴 함으로써 생체시계의 작동 에 관여하는 새로운 인산화 효소를 발굴하고자 하였다.
4
Table 1 Kinases which regulate circadian clock in mammals
This table is known for circadian kinase. kinases which is discovered until now are CKⅠ, CKⅡ, GSK3β, AMPK, CDK5 just five kinases.
5
C. 핵 이동
핵심 생체시계 단백질들이 핵 안으로 들어가거나 세포질에 머무는 시기는 생체시계 기전을 조절하고 유지하는데 필수적이다. SCN 뉴런들이 동기화 (synchrony)되면 CRY1, CRY2, PER1, PER2 단백질들이 핵 안에 많이 존재한다고 보고된 바 있다 (Field et al., 2000). 게다가 많은 생체시계 단백질들은 nucleo-cytoplasmic shuttling을 조절할 수 있는 nuclear entry와 export signal을 가지고 있다
(Miyazaki et al., 2001; Yagita et al., 2002; Tamanini et al., 2005). 생체시계 조절 기작에
서 24시간이라는 정확한 생체리듬을 갖기 위해서는 생체시계 단백질의 핵으로의 이동이 시간에 따라 정교하게 조절되는 것이 매우 중요하다. 여전히 시간에 따른 생체시계 단백질의 세포 내 위치 조절은 밝혀나가야 할 부분이 많지만, 이전의 연구들로 보아 인산화와 같은 번역 후 조절 (post-translational modification)이 중요 하게 작용한다는 것은 분명하다 (Lee et al., 2001; Kim et al., 2002; Gallego and Virshup, 2007; Diernfellner and Schafmeier, 2011).
D. 연구의 필요성 생체시계를 이루는 핵심 단백질들은 하루를 주기로 양, 세포 내 위치 및 활 성 등이 인산화에 의해 조절되어 생체시계가 24시간의 주기를 갖고 다양한 생리 작용들이 적합한 시간에 일어날 수 있도록 한다. 본 연구에서는 시도된 적 없는 인간 인산화 효소 라이브러리를 이용한 새로운 스크리닝 방법으로 생체시계 단 백질을 인산화 시키는 인산화 효소를 찾고, 인산화로 인해 핵 안으로 이동하는 분자적 기작에 대해 연구하고자 한다.
6
Ⅱ 재료 및 방법
A. Plasmids and cloning1. Plamids
pUB6(A)-egfp 클로닝은 egfp primer를 가지고 PCR 하여 pUB6(A) vector를 제 한효소 XhoI과 NotI으로 잘라 삽입하여 제작했다. pUB6(A)-rfp 클로닝은 rfp primer 를 가지고 PCR하여 pUB6(A) vector에 제한효소 KpnI과 EcoRI으로 잘라 삽입하여 제작했다. pUB6(A)-mcry1-egfp과 pUB6(A)-mper2-egfp는 동국대학교 약학대학, 고혁 완 교수님 실험실에서 받았다. pUB6(A)-mcry2-egfp 클로닝은 mcry2 primer로 PCR 을 한 후 앞에서 클로닝한 pUB6(A)-egfp vector를 제한효소자리 EcoRV와 AgeI을 이용하여 삽입하였다. pUB6(A)-clk-egfp, pUB6(A)-bmal1-egfp, pUB6(A)-mstat3-egfp 클 로닝은 각각 clk, bmal1, mstat3 primer로 PCR하여 제한효소 BamHI과 EcoRV를 사 용하여 pUB6(A)-egfp vector에 삽입하였다. pCMV-HA-mcry1, pCMV-HA-mcry2, pCMV-HA-mper2, pCMV-HA-clk, pCMV-HA-bmal1 클로닝은 각각 mcry1/2, mper2, clk,
bmal1 primer로 PCR한 후 제한효소 XhoI과 NotI을 사용하여 pCMV-HA vector에 삽 입하였다. pUB6(A)-rfp-V5, pUB6(A)-hprkra-V5는 rfp와 hprkra의 primer를 이용하여 pUB6(A)-V5 vector를 제한효소 EcoRV와 NotI으로 잘라 삽입하여 클로닝하였다.
pUB6(A)-hvrk3 nls mut.-egfp는 hvrk3 NLS mutant를 primer를 이용하여 PCR한 후
pUB6(A)-V5 vector를 제한효소 자리 XhoI과 XbaI에 삽입하여 클로닝하였다. 자세
7
2. NLS mutagenesis
두 번의 overlapping PCR (Roche Applied Science, Rapid Translation System, Application Note No. 13/2003)을 이용하여 hvrk3의 NLS (49KRGLNSSFETSPKKVK64)
에 mutation을 일으켰다. 첫 번째 overlapping PCR은 49
KR49AA으로 치환시키기
위해 KpnI-hvrk3F primer와 hvrk3-166R primer (5’-ACT GGA GTT CAG CCC TGC CGC TGA GCC TTG GAA-3’)를 넣어 약 200bp정도의 hvrk3 앞부분을 PCR했으며,
hVrk3-130F primer (5’-TCC TTC CAA GGC TCA GCG GCA GGG CTG AAC TCC-3’)와
XhoI-hvrk3R primer를 이용하여 hvrk3의 뒷부분인 약 1200bp를 PCR하였다. 얻어진 PCR product를 각각 1ul씩 주형으로 삼아 KpnI-hvrk3F와 XhoI-hvrk3R primer로 전체
hvrk3 (KR49AA)를 얻었다. 두 번째 PCR은 61KKVK61AAVA로 치환시키기 위해 수행하였다. 첫 번째 PCR로 얻어진 hvrk3 (KR49AA)를 주형으로 KpnI-hvrk3F primer와 hvrk3-206R primer (5’-GAC GGT GCT GGA CCA TGC CAC TGC CGC AGG
AGA GGT TTC AAA-3’)로 hvrk3의 약 200bp PCR하였으며, hvrk3-166F primer
(5’-TTT GAA ACC TCT CCT GCG GCA GTG GCA TGG TCC AGC ACC GTC-3’)와
XhoI-hvrk3R primer를 이용하여 hvrk3의 약 1200bp를 PCR하였다. 얻어진 두 개의 product들을 각각 1ul씩 주형으로 삼아 KpnI-hvrk3F와 XhoI-hvrk3R primer로 전체
hvrk3 (KR49AA, KKVK61AAVA)를 얻었다. 모든 PCR은 pfuTurbo DNA polymerase (Str atagene Co., La Jolla, CA, U.S.A)를 이용했으며 PCR 반응조건은 pfuTurbo DNA
8
9
B. Establishment of human kinome expression library
HIP FLEXGene human kinase collection (Harvard Univ. DF/HCC DNA Resource Core, http://plasmid.med.harvard.edu /PLASMID/Home.jsp)에 RFP를 표지하기 위한 첫
번째 방법으로 creator system (New England Biolabs)을 사용하였다. Donor vector (pDNR-kinase)와 acceptor vector (pLP-RFP)를 Cre recombinase, 1X Cre buffer, 1X BSA,
1mM spermidine을 함께 넣어 RT에서 15분간 ligation시킨 후에 70℃에서 5분간
heat shock을 주었다. DH5α competent cell에 ligation된 시료를 넣고 30분 후에 42℃
에서 50초동안 heat shock을 가한 후 곧바로 얼음 위에 2분 이상 두었다. SOC을 첨가한 후 1시간 30분 동안 shaking incubator에서 배양하여 30ug/ml chrolampheniclol-7% sucrose LB plate에 transformation을 수행하였다. 두 번째 방법으
로 사용된 Gateway system은 Center for Cancer Systems Biology (CCSB)/Broad Institute Kinase Open Reading Frame Collection (Invegrogen, http://www.addgene.org/human_kinases)을 이용했다. Entry kinase 150ng, pDest vector
150ng, LR clonase Ⅱ (Invitrogen) 2ul, TE buffer pH 8.0을 총 양이 10ul가 되게 넣어 준 25℃에서 2시간 동안 LR reaction을 시켰다. 반응을 끝내기 위해 proteinase K 1ul를 넣어 준 뒤 37℃에서 10분간 처리했다. 그 후 DH5α competent cell에 heat
shock을 주고 30ug/ml kanamycin LB plate에 transformation하였다.
C. Cell culture and transfections
실험에 사용된 NIH 3T3, HEK 293T 세포는 10%의 fetal bovine serum (FBS)와 penicillin/streptomycin이 첨가된 Dulbecco’s modified Eagle’s medium (DMEM)을 이용
10
하여 5% CO2, 37℃ 항온기에서 배양시켰다. 1. for fluorescence microscopy
인간 인산화 효소 유전체를 스크리닝할 세포를 준비하기 위해, 먼저 24well plate (1well/500ul)에 cover slip을 얹고 70% 에탄올로 cover를 두 번 씻은 후 UV를
20분 이상 쬔다. 멸균이 끝나면 멸균된 3차 증류수로 에탄올을 세 번 씻어낸다.
NIH 3T3 세포의 경우 그 위에 1× 105 cells/ml로 세포를 분주해 준다. HEK 293T
세포의 경우 에탄올로 멸균을 끝낸 후 poly-L-lysine (Sigma aldrich) 100ug/ml을 20 분간 처리하고 멸균한 3차 증류수로 3번 씻어낸다. 후에 물기를 제거한 cover slip 위에 1.25× 105 cells/ml로 분주한다. 세포를 쪼갠 후 24시간동안 5% CO2, 37℃에서
배양한다. 60-80%정도 cell이 자랐을 때 attractene (Qiagen)을 사용해서 transfection 을 수행한다. 24well plate의 경우 총 plasmid의 양을 400ng (스크리닝의 경우, clock protein(s)-EGFP 200ng: kinase(s)-RFP 200ng=1:1) 넣어주고 serum free media로 총 양 을 60ul로 맞춰준다. 그 후 잘 섞인 attractene reagent를 1.5ul 첨가해 잘 섞어준 후 complex가 잘 형성되도록 RT에서 10-20분간 둔다. 시간이 지나면 하루 전에
24well plate에 배양해둔 세포의 10% DMEM으로 교체해준 후, 준비한 시료를 방울
방울 세포표면에 떨어뜨린다. 2. for western blot and IP
Western blot과 immunoprecipitation은 HEK 293T 세포에서 수행하였다. Plate에
11
3번 씻어낸다. 실험에서는 12well (1well/1ml) 혹은 6well (1well/2ml) plate를 사용했
으며 세포는 1.67× 105 cells/ml으로 분주 후 24시간 동안 5% CO2, 37℃에서 배양
한다. 세포가 60-80%정도 자라면 attractene (Qiagen)을 사용해서 transfection을 수행 한다. 12well plate의 경우 plamid 총량은 1ug에 총 양을 serum free media를 이용해 80ul로 맞추고, 6well의 경우 plasmid 총량은 1.2ug, serum free media의 총 양은
100ul로 맞춘다. 준비된 시료에 attractene reagent를 12well에는 3ul, 6well에는 4.5ul
를 넣어준 후 vortex로 잘 섞어 10-20분간 RT에 둔다. 시간이 지나면 미리 배양해 둔 세포의 media를 교체해준 후, 준비한 시료를 한 방울씩 세포표면에 떨어뜨린 다. D. Fluorescence microscopy Transfection시킨 세포를 형광현미경으로 관찰하기 위해서 세포 고정과 핵 염색을 수행하였다. Transfection후 24시간 후에 1X PBS로 두 번 헹궈주고 4% paraformaldehyde로 20분간 세포를 고정한다. 시간이 지나면 1X PBS로 4% paraformaldehyde를 두 번 씻어낸 후 DAPI 염색 전에 핵에 구멍을 뚫어주기 위해 0.1% triton을 포함하는 1X PBS에 RT에서 5분, 4℃에서 30분간 처리한다. 핵을 염 색하기 위해 bis-benzimide를 1X PBS에 1:5000으로 희석시켜 RT에서 30분에서 1 시간 정도 담가둔다. DAPI 염색이 끝나면 현미경 관찰 시 깨끗하게 볼 수 있도록 1X PBS로 3번 씻는다. Cover slip은 3차 증류수에 헹궈준 후 휴지 (kimtech wipers
12
mounting media를 한 방울 정도 떨어뜨린 후 세포가 media에 닿도록 엎어준다. 시
료는 현미경에서 관찰하고 AxioVision Imaging System (Carl Zeiss MicroImaging GmbH, 07740 Jena, Germany)으로 분석하였다.
E. Quantitative real-time RT-PCR
NIH 3T3 세포를 -세포의 양에 따라 200-500ul- Tri-reagent (Molecular Research
center, Inc)로 lysis하여 RNA를 얻었다. 이렇게 얻은 RNA를 oligo dT primer, dNTP
와 함께 65℃에서 5분동안 끓여준 후 reverse transcriptase (Takara)와 RNase inhibitor 를 넣어 PCR을 통해 cDNA를 합성했다. 만들어진 cDNA를 주형으로 Quantitect SYBR Green RCR kit (Qiagen)을 이용하여 Corbett research Rotor gene 6000 (Corbett
Life Science)에서 real-time RT-PCR을 수행했다.
F. Cell lysis
Transfection후 원하는 시간 동안 배양한 세포는 얼음 위에 올려 1X PBS로
한 번 씻어낸다. 미리 만들어 놓은 lysis buffer를 6well 기준 120ul 넣고 scrapper를 이용하여 세포를 긁어 모았다. 세포 용해를 위해 5분 간격으로 vortex 하면서 15-30분 동안 얼음 위에 둔다. 시간이 지나면 4℃ 15000rpm으로 15분 동안 원심 분
리 후 상층액을 새로운 tube로 옮겨준다. 상층액은 ELIZA leader (BioTex Inc., Houston, Tx, U.S.A)를 이용하여 595nm에서 흡광도를 측정한다.
13
Lysis를 하기 전에 RIPA buffer (150mM NaCl, 10mM Na2HPO4 pH7.2, 0.5%
sodium deoxycholate, 1% NP-40)에 1mM DTT, 0.5mM PMSF, 1mM EDTA, 2.5mM NaF,
2mM Na3VO4, 1X protease inhibitor가 되도록 첨가하여 lysis buffer를 만들었다.
2. for IP
Modified RIPA buffer (50mM NaCl, 50mM Tris-HCl pH7.5, 0.25% sodium deoxycholate, 1% NP-40, 5% glycerol)에 1mM EDTA, 1X protease inhibitor cocktail, 1X
phosphatase inhibitor Ⅱ, 1X phosphatase inhibitor Ⅲ가 되도록 lysis buffer를 만든다.
G. Western blotting
단백질의 양적인 변화를 알아보기 위해 western blot을 수행했다. 용도에 맞 춰 5-15% polyacrylamide gel을 제작하여 사용했다 (30% polyacrylamide/Bis solution, 37.5:1, BIO-RAD). 이전에 준비해둔 단백질 시료를 원하는 농도로 계산한 후 4X
SDS-PAGE sample buffer (2.5ml 1M Tris-HCl pH6.8, 4ml 100% Glycerol, 0.8ml
β-mercaptoethanol, 0.8g SDS, 2.5ml Autoclaved water)를 첨가하여 95℃에서 3분간 끓여
변성시켰다. 준비된 샘플들을 가지고 SDS-PAGE 전기영동법 (Sodium Dodecyl Sulfate-Polyacrylamide gel Electrophoresis, Laemmli method)을 통해 단백질을 분리하
였다. Mini-Protein Tetra Cell (BIO-RAD Laboratories, Inc., Hercures, DA, U.S.A)을 이용 하였고, 전기영동용 running buffer (1L; 3.025g Tris, 14.4g Glycine, 1% SDS, DW)를 사 용하여 전기영동 하였다. Protein marker는 GangNam stain Prestained Protein Ladder (iNtron)를 이용했으며 원하는 사이즈의 밴드가 알맞게 보일 때까지 내린 후
14
transfer를 진행했다. Gel과 함께 transfer buffer (1L; 5.82g Tris, 2.93g Glycine, 20%
Methanlol, 0.0375% SDS, DW)를 충분히 적신 gel blot paper (Whatman Inc., Springfield
Mill, Kent, U.K)와 PVDF membrane (Immobilon, Millipore Inc., U.S.A)을 sandwich를 만들어 Trans-Blot SD semi-dry electrophoretic transfer cell (BIO-RAD Laboratories Inc., Hercures, CA, U.S.A)를 이용해 240mA에서 40분간 진행하였다. Membrane blocking
은 5% milk에서 40분 이상 이루어졌고 이어서 1차 항체를 포함하는 5% milk로 4℃ 에서 O/N으로 반응시킨 후 2차 항체가 포함된 5% milk에서 3-6시간 반응시켰다. 항체 반응을 끝낸 후에는 10분 간격으로 6번, 한 시간 동안 1X TBST (4M NaCl, 1M Tris-HCl pH7.5, 0.05% Tween20, DW)로 헹궈내었다. Developing은 Pierce ECL
Western Blotting Substrate (Thermo Co., Meridian Rd., Rockford, U.S.A) 혹은 Immobilon
Western chemiluminescent HRP substrate (Millipore Co., Billerica, MA, U.S.A)를
membrane에 묻혀 CP-BU NEW medical X-ray film blue (Agfa HealthyCare Co.,
Soptestraat, Mortsel, Belgium)에 적정시간 노출시켜 현상하였다.
H. Immunoprecipitation(IP) and λPhosphatase treatment
HEK 293T 세포를 용해하여 얻어 낸 단백질 시료를 동일한 농도로 맞춘 후
HA (12CA5) (Roche Co., Mannheim, Germany) 혹은 V5 (Invitrogen Co., Carlsbad, CA,
U.S.A) 1차 항체를 한 튜브당 3ul씩 넣고 4℃에서 O/N으로 rotator를 이용하여 반
응시켰다. 반응이 끝나면 세포를 용해할 때 사용했던 buffer (Modified RIPA)로 3번 바꿔 준 GammaBind G sephrose (GE Health care Bio-Sciences AB., Bjorkgatan, Uppsala,
15
Sweden) Bead를 한 튜브당 25ul씩 첨가하고 4℃에서 3-6시간 rotator를 이용하여
반응시켰다. 시간이 지나면 Modified RIPA buffer로 3번 헹궈내고 bead만 남긴다. 준비한 1.2X sample buffer를 한 샘플에 30ul씩 첨가하고 95℃에서 7분간 끓여 bead를 끊어낸 후 western blotting을 수행하였다.
λPPase를 처리하기 위해서는 bead를 첨가하고 반응시킨 다음 1X lambda
protein phosphatase buffer (5mM Tris-HCl pH7.5, 0.01mM EDTA, 0.5mM DTT, 0.001% Triton X-100, 0.2mM MnCl2, 0.01mg/ml bovine serum albumin, autocleaved water) 첨가하
여 buffer를 바꿔준다는 생각으로 씻어낸다 (Lee et al., 1998). 그리고 1X λPPB 50ul 로 bead를 풀어내고 λPhosphatase (New England Biolabs) 200units을 첨가하여 30℃에 서 10분간격으로 tapping하면서 30분간 반응시킨다. 반응이 끝난 후에는 원심분리 로 bead가 딸려오지 않도록 buffer를 버리고 준비한 1.2X sample buffer를 한 샘플 에 30ul씩 첨가하고 95℃에서 7분간 끓여 bead를 끊어낸 후 western blotting을 수 행하였다.
I. Fly genetics
hprkra knockdown 실험을 위하여 유전자를 조직 특이적으로 발현하기 위한 방법으로 잘 알려진 GAL4-UAS 시스템을 이용했다 (Brand and Perrimon, 1993). hprkra 유전자에 특이적인 dsRNA를 발현하는 초파리는 VDRC108358, VDRC22453 를 사용하였으며 VDRC (Vienna Drosophila Research Center)로부터 구입하였다. timeless (tim) 발현세포에서만 knockdown을 유발하기 위하여 timeless enhancer
16
sequence에 의해 Gal4의 발현이 조절되고, RNA interference (RNAi) 효과를 증가시
키기 위해 dicer2가 발현되도록 UAS-dicer2; timeless-UAS-Gal4 초파리를 사용하였 다. UAS-dicer2; Timeless-UAS-Gal4 초파리와 hprkra RNAi 초파리를 교배하여 얻은 자손에서는 TIM 단백질을 발현하는 세포 내에서 특이적으로 hprkra가 knockdown 된다.
J. Locomotor activity assays
Drosophila Activity Monitoring system (DAM system, Trikinetics)을 이용하여 초 파리의 locomotor activity를 측정했다. 유리 튜브 안에 어린 수컷 초파리를 한 마 리씩 넣고 원하는 온도 (25℃ 혹은 29℃) 항온기에서 12시간 간격으로 빛과 어둠 으로 4일동안 적응시켰다. 이후 연속으로 constant darkness condition (DD)을 12시간 간격으로 7일간 유지시켰다. 12:12LD zeitgeber time zero [ZT0] sum light phase가 시작 되는 시점으로 정의하며 12:12DD circadian time으로 명명한다. 초파리의 locomotor activity 분석은 Dr. F. Rouyer (Institut de Neurobiologie Alfred Fessard, CNRS, France)로
부터 받은 FassX software (MacOSX를 위한 Fly Activity Analysis Suite)를 이용하였다. Period는 초파리 각각을 chi-square periodogram analysis로 계산하여 각 유전형의 그
룹별 평균을 산출하였다. Power는 DD동안 초파리 리듬의 strength의 정도를 나타 낸다. 각 초파리들은 power가 10이상을 가지고 width (periodogram 95% confidence line보다 30분 위에 peak된 수를 표시)값이 2이상의 리듬을 가지는 초파리로 규정
17
Ⅲ. 결과
A. 인간 인산화 효소 라이브러리 구축 생체시계 단백질의 인산화는 시간에 따른 인산화에 의해 생체시계 단백질 의 안정성, 활성, 그리고 세포 내 분포를 조절함으로써 생체시계의 작동에 매우 중요하게 작용할 것으로 알려져 있다. 예를 들어, 생체시계 핵심 단백질인 mPER2는 casein kinase I δ (CKIδ)와 casein kinase I ε (CK1ε)을 통해 핵 안으로 이동하지 못하고 세포질에 머문다 (Maier et al., 2009). 이는 Figure 4B 그래프에서도 확 인할 수 있다. 앞에서 언급했듯이 현재까지 포유류에서 밝혀진 생체시계 단백질 인산화 효소는 CKⅠ α/ε/δ, CKⅡ, GSK3β, AMPK, CDK5 5개가 전부이다 (Table 1). 이러한 인산화 효소를 발굴하기 위해 먼저 human kinome expression library를 확립 하고자 했다. 인간의 유전체에 존재하는 533개의 인산화 효소를 포유류 세포에서 발현 가능한 DNA vector로 세가지 방법을 사용하여 클로닝을 수행하였다. 첫 번 째 클로닝 방법으로 각각의 인산화 효소를 일정한 방향으로 human kinase ORF (Open Reading Frame)에 붉은색 형광을 나타내는 RFP 단백질을 표지하는 Creator
시스템 (New England Biolabs)을 사용하였다 (Figure 1A). Creator 클로닝은 많은 양 의 목적유전자를 다양한 tag vector (Acceptor vector)에 Cre recombinase를 이용하여 간단하게 클로닝을 완료할 수 있는 장점을 가지고 있다. Creator 시스템을 이용하 여 177개의 인산화 효소에 RFP표지를 완료하였다. 이 방법으로 클로닝 되지 않
18
은 인산화 효소는 gateway 시스템 (Center for Cancer Systems Biology (CCSB)/Broad Institute Kinase Open Reading Frame Collection, Invitrogen) (Figure 1B)과 제한효소를
이용한 전통적인 방법으로 클로닝 하였다. 이 클로닝은 공동연구자인 동국대학교 약학대학, 고혁완 교수님 실험실의 송지은, 이나영 학생과 공동연구로 진행하였 다. 그 결과 전체 인산화 효소 533개 중 385개가 클로닝이 완성되어 72% 이상 RFP표지를 완료하였다.
19
Figure 1 Schematic diagram of establishment of fluorescent peptide tagged human kinome expression library. A, Via Creator system (New England Biolabs) pLP-RFP
acceptor and pDNR-DURL donor vectors are recombined in the presence of Cre-recombinase to generate RFP tagged kinase expression vector. B, Via Gateway system (Invitrogen) pDEST-mCherry-N1 and pDONR 223 vectors are recombined in the presence of LR clonase to generate mCherry tagged kinase expression vector. CmR, Chloramphenicol resistance; Kan/NeoR, kanamycin and neomycin resistance; SacB, levansucrase of Bacillus subtilis which confers lethality in the presence of sucrose; ccdB, a lethal gene that targets DNA gyrase.
20
B. 인간 인산화 효소 스크리닝
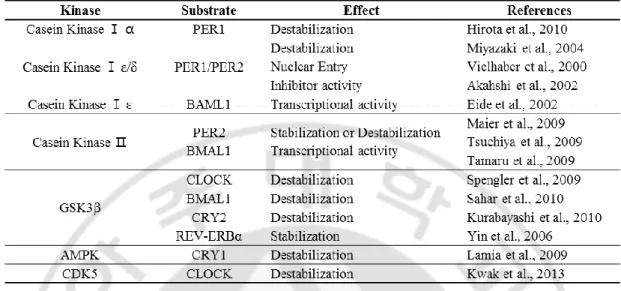
현재까지 스크리닝을 완료한 인산화 효소를 표로 나타내었다 (Table 2). 생 체시계 단백질을 안정적으로 발현한다고 알려진 NIH 3T3 세포에 EGFP로 표지된 생체시계 단백질 mCRY1과 human kinome expression library의 인산화 효소들을 각 각 일시적으로 transfection 시킨 후 형광 현미경 관찰을 통해서 스크리닝했다. mCRY1을 과 발현시키면 핵에 존재한다. mCRY1 스크리닝 결과 118개 중 114개
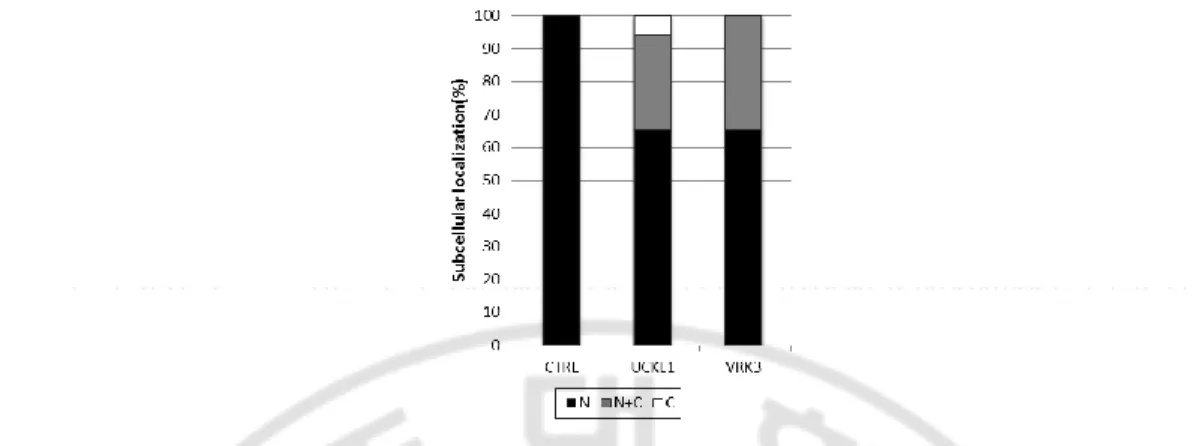
의 인산화 효소는 100% mCRY1이 핵에 존재 했으며, mCRY1의 위치 변화를 일으 키는 인산화 효소는 hUCKL1, hVRK3 2개가 전부였다 (Figure 2). hUCKL1의 경우 mCRY1은 약 66%가 핵, 약 28%가 핵과 세포질, 약 6%가 세포질에 존재했고 hVRK3의 경우 약 65%가 핵, 약 35%가 핵과 세포질에 존재했다. 사실, transfection 효율이 떨어지는 NIH 3T3세포에서 스크리닝을 진행하는 것은 매우 어려웠다. 이에 다른 생체시계 단백질인 mPER2의 위치 조절에 관여하 는 인산화 효소 스크리닝은 HEK 293T 세포를 가지고 진행했다. mPER2가 과 발 현되면 70-80%가 핵에 나머지가 핵과 세포질 혹은 세포질에만 존재한다. 이러한 mPER2의 위치를 두드러지게 변화시키는 인산화 효소에는 hNEK2, hCKMT1B,
hPRKRA, hETNK1, hCSNK1E, hMST1R 그리고 hVRK3가 있었다 (Figure 3). hNEK2
와 hCKMT1B는 핵과 세포질에 존재하는 mPER2를 약 50%로 증가시켰으며, hPRKRA는 mPER2의 위치를 핵이 대략 44%, 핵과 세포질이 40%, 세포질이 16%
21
로 세포질에 존재하는 mPER2가 증가했다. hCSNK1E는 이전 연구를 통해 잘 알 려진 mPER2의 인산화 효소로 대략 핵에 5%, 핵과 세포질에 25%, 세포질에 70% 존재하게 했다 (Maier et al., 2009). hMST1R의 경우 mPER2의 위치를 대략 핵에 46%, 핵과 세포질에 41%, 세포질에 13%로 변화시켰으며 hVRK3의 경우 핵에
22
Table 3 List of screened kinases
This table shows kinases with which screening is performed o indicates kinases which alter subcellular localization of mCRY1 or mPER2. x indicates kinases which did not alter subcellular localization of mCRY1 or mPER2.
23
Figure 2 Kinases altering subcellular localization of mCRY1 in NIH 3T3 cells.
Subcellular localization of pUB6(A)-mcry1-egfp transfected with pUB6(A)-rfp (CTRL) or with pLP-rfp-huckl1 and pLP-rfp-hvrk3 (indicated) in NIH 3T3 cells was quantified. Among 118 kinases screened in this study, only 2 kinases alter subcellular localization of mCRY1 in NIH3T3 cells and the results are shown.. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). At least 20cells were counted for each experiment.
26
Figure 3 Quantification of subcellular localization of mPER2 in HEK 293T cells. HEK 293T cells were transiently transfected with pUB6(A)-mper2-egfp either with RFP (CTRL) or with RFP-kinase (indicated). Note that co-expression of hNEK2, hCKMT1B, hPRKRA, hETNK1, hMST1R, or hVRK3 dramatically changes subcellular localization of mPER2 in HEK 293T cells. As previously reported, co-expression of hCSNK1E increased the ratio of cytoplasmic localization of mPER2. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white).
27
C. VRK3
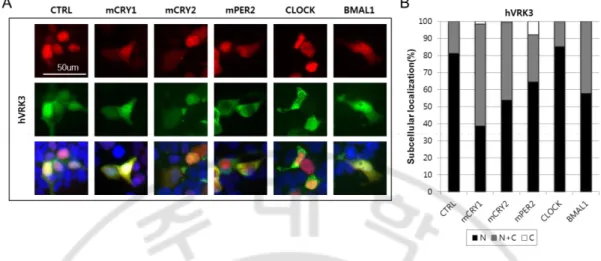
1. hVRK3는 mCRY1/2, mPER2, BMAL1 단백질의 세포 내 위치를 조절한다.
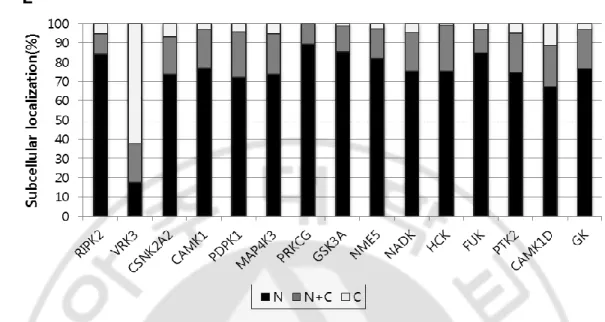
포유류의 핵심 생체시계 단백질인 mCRY1의 세포 내 분포를 변화시키는 인산화 효소를 NIH 3T3 세포에서 스크리닝하여 hVRK3 (vaccinia-related kinase 3)을 찾았다. 다른 포유류 세포주인 HEK 293T 세포에서 mCRY1의 세포 내 분포에 미 치는 영향을 조사하였을 때에도 동일한 결과를 얻을 수 있었다 (Figure 4). hVRK3 는 mCRY1 단백질뿐만 아니라 mCRY2, mPER2 그리고 BMAL1 단백질의 세포 내 위치도 변화시켰다. 흥미롭게도 핵심 생체시계 단백질 중 CLOCK 단백질의 분포 는 변하지 않았다. mCRY1/2와 BMAL1은 주로 핵에서 관찰되다가 hVRK3와 함께 발현되면 핵과 세포질에 동시에 발현되는 양상을 보였다. 또한 mPER2는 핵에 75%, 핵과 세포질에 25% 존재하다가 hVRK3와 일시적으로 동시에 발현시킬 때 세포질에만 60% 이상 존재했다. 반면 CLOCK 단백질은 hVRK3의 유무와 관계없 이 핵에 절반, 핵과 세포질에 절반 가까이 위치했다. hVRK3는 핵에 존재한다고 알려져 있으며 (Nichols and Traktman, 2004; Scheeff et al., 2009), Figure 5와 같이 실제 로 핵에 80%이상 존재하는 것을 관찰했다. 또한 hVRK3가 CLOCK을 제외한 mCRY1/2, mPER2, BMAL1과 함께 발현되면 핵과 세포질에 존재하는 것을 확인하
였다. 종합하면, hVRK3와 mCRY1/2, mPER2, BMAL1 단백질이 함께 발현되면 mCRY1/2, mPER2, BMAL1과 hVRK3의 위치가 핵 안에 있는 비율보다 세포질에
28
Figure 4 Co-transfection of hvrk3 changes the subcellular localization of mCRY1/2, mPER2 or BMAL1 in HEK 293T cells. A, HEK 293T cells were transiently transfected
with mcry1/2-egfp, mper2-egfp, clock-egfp, or pUB6(A)-bmal1-egfp (indicated on top of the panel) either with pUB6(A)-rfp (CTRL) or with pUB6(A)-rfp-hvrk3 (hVRK3). Hoechst stained nuclei were depicted as blue. B, Quantification of subcellular localization of clock proteins. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). More than 80 numbers of cells were counted for each experiment.
29
Figure 5 hVRK3 increases subcellular localization of nucleus and cytoplasm more than nucleus in co-expression with mCRY1/2, mPER2, BMAL1. A, HEK 293T cells were
transiently transfected with pUB6(A)-rfp-hvrk3 with pUB6(A)-egfp (CTRL) or with pUB6(A)-mcry1/2-egfp, pUB6(A)-mper2-egfp, pUB6(A)-clock-egfp, or pUB6(A)-bmal1-egfp, as indicated on top of the panel. Hoechst stained nuclei were depicted as blue. B, Quantification of subcellular localization of hVRK3. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white).
30
2. hVRK1과 hVRK2는 mCRY1의 세포 내 위치를 바꾸지 않는다.
hVRK3는 vaccinia related kinase family 중 하나로 hVRK3 외에 hVRK1과
hVRK2가 여기에 속하며, ATP 결합 모티프 안에 몇몇 잔기가 치환되어 효소 활 성이 없는 인산화 효소 (pseudokinase)이다 (Nichols and Traktman, 2004). hVRK1과 hVRK2가 mCRY1과 같은 생체시계 단백질들의 분포에도 영향을 주는 지 실험했
다. hVRK1과 hVRK3는 각각 C-말단과 N-말단에 핵 안으로 이동할 수 있는 신호 (Nuclear Localization Sequence)를 가지고 있어 핵 안에 위치한다. 반면 hVRK2는
세포질에 있으며 이미 이전 연구에 의해 밝혀졌지만 이 실험에서도 확인할 수 있다 (Nichols and Traktman, 2004). 결과적으로 hVRK1,2는 mCRY1의 세포 내 위치 를 바꾸지 않았다 (Figure 6). 따라서 VRK3가 생체시계 단백질의 세포 내 위치를 바꾸는 것은 VRK의 직접적인 인산화 활성과는 관련이 없는 것으로 보여진다.
31
Figure 6 Co-transfection of hvrk1/2 did not alter the nuclear localization of mCRY1. A,
B; NIH 3T3 cells were transfected with pUB6(A)-mcry1-egfp in combination with either pUB6(A)-rfp (CTRL) or pLP-rfp-hvrk1, pLP-rfp-hvrk2, pUB6(A)-rfp-hvrk3 as indicated on the left side of the column. Hoechst stained nuclei were depicted as blue. B, Quantification of subcellular localization of mCRY1. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). Numbers in each bar show total number of cells counted.
32
3. hVRK3는 mSTAT3의 세포 내 위치에는 영향을 미치지 않았다
앞에서 hVRK3가 CLOCK을 제외한 mCRY1/2, mPER2, BMAL1 단백질의 세 포 내 위치를 변화시키는 것을 확인했다. 이러한 hVRK3가 생체시계 단백질에 국 한되게 작용하는 것인지 아니면 또 다른 단백질들의 세포 내 위치 조절에도 관 여하는 지를 알아보기 위해 핵과 세포질을 이동한다고 알려진 mSTAT3 (signal transducer and activator of transcription-3)를 HEK 293T 세포에서 hVRK3와 함께 일시
적으로 발현시켰다 (Reich, 2013). mSTAT3 단백질을 HEK 293T 세포에서 일시적으 로 발현시켰을 때 핵과 세포질에 함께 존재했으며, hVRK3와 함께 발현되어도 그 위치가 변하지 않았다 (Figure 7). 또한 mSTAT3가 핵과 세포질에 고르게 퍼져 있 어도 hVRK3의 세포 내 위치는 핵에 주로 존재했다. 다시 말해 hVRK3와 mSTAT3는 서로 세포 내 위치에 영향을 미치지 않았다.
33
Figure 7 Co-transfection of hvrk3 did not alter the subcellular localization of mSTAT3.
A, HEK 293T cells were transfected with pUB6(A)-stat3-egfp in combination with pUB6(A)-rfp (CTRL) or pUB6(A)-rfp-hvrk3 as indicated on the left side of the panel. When pUB6(A)-rfp-hvrk3 was transfected without mstat3, pUB6(A)-egfp was co-transfected (CTRL). Hoechst stained nuclei were depicted as blue. B,C; Quantification of subcellular localization of hVRK3 (B) and mSTAT3 (C). N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). Numbers in each bar show total number of cells counted.
34
4. hVRK3는 핵 안에서 작용하여 생체시계 단백질 -mCRY1, mPER2, BMAL1-
을 세포질에 머물게 한다.
이번에는 hVRK3가 과 발현 됨으로써, mCRY1/2, mPER2, BMAL1의 핵으로의 이동을 막는 지, 혹은 핵에서 세포질로 나오도록 유도하는 지 알아보고자 hVRK3-NLS mutant를 제작했다. hVRK3의 NLS로 알려진 아미노산 49–64번 중 49,
61-62, 64번 리신 (K)과 50번 아르기닌 (R)을 알라닌 (A)으로 치환시켜 NLS의 변
이를 만들었다. (Nichols and Traktman, 2004; Inagaki et al., 2006; Lupberger et al., 2013) (Figure 8A). hVRK3-NLS mutant는 100% 핵으로의 이동을 막진 못했지만, 60%이상
이 세포질 (C)에만 존재했으며 40% 정도가 핵과 세포질 (N+C)에 함께 존재하는 것을 볼 수 있었다 (Figure 8B,C). 이는 알려진 NLS 이 외에 다른 NLS가 존재할 수 있음을 암시한다.
hVRK3-NLS mutant를 생체시계 단백질들과 각각 발현시켰을 때 생체시계
단백질들의 세포 내 분포 변화를 살펴보았다 (Figure 9). hVRK3-NLS mutant를 발 현시켰을 때 hVRK3보다 mCRY1, mPER2, BMAL1이 핵에 더 많이 존재하는 것을 볼 수 있다. 다시 말하면 hVRK3로 인해 생체시계 단백질 -mCRY1, mPER2, BMAL1- 의 세포 내 위치가 달라졌는데 (Figure 4B), hVRK3를 핵 안으로 들어가
지 못하게 했을 때 그 작용을 완화시키기는 하였으나 hVRK3가 발현되지 않았을 때의 mCRY1/2, mPER2, BMAL1의 세포 내 위치로 회복시키지는 못하였다. CLOCK의 경우 hVRK3와 hVRK3-NLS mutant간의 차이가 없었다. 이 결과는
35
hVRK3 단백질이 mCRY1, mPER2, BMAL1과 같은 생체시계 단백질이 핵 안으로
수송되는 단계에 작용한다기 보다는 핵 안에 있는 mCRY1, mPER2, BMAL1이 세 포질 바깥으로 수송되는 단계에 작용하여 세포 내 위치 분포에 영향을 미치는 것을 나타낸다.
36
Figure 8 hVRK3-NLS mutant is present predominantly in the cytoplasm in HEK 293T cells. A, Schematic diagram of hVRK3 protein NLS sequence (amino acids 49-64) is shown.
The putative bipartite NLS motifs are indicated with a red box. The catalytic domains are indicated by diagonal box. The arginine residues of 49, 61, 62, 64 amino acid and a lysine residue of 50 amino acid were all mutated to alanine. B,C; HEK 293T cells were transfected with pUB6(A)-egfp in combination with either pUB6(A)-rfp-hvrk3 or pUB6(A)-rfp-hvrk3 nls mut as indicated on the left side of panel. Hoechst stained nuclei were depicted as blue. Note that NLS mutation of hVRK3 affected the subcellular localization in HEK 293T cells. Wild-type version of hVRK3 (hVRK3) is predominantly located in nucleus (N). Whereas NLS mutant version of hVRK3 (hVRK3-NLSmut) is present predominantly in the cytoplasm (C) or distributed evenly in the nucleus and cytoplasm (N+C).
37
Figure 9 The extent of alterations in subcellular localization of mCRY1, mPER2, and BMAL1 was attenuated in the case of co-expressing NLS mutant version of hVRK3 in HEK 293T cells. Wild-type or NLS mutant versions of hVRK3 was expressed with clock
proteins (indicated on top of the panel) respectively in HEK 293T cells. Representative images show the subcellular localization of mCRY1, mPER2, CLOCK, and BMAL1. In the case of mCRY1, mPER2 and BMAL1, nuclear localization was increased by co-transfection with hVRK3-NLS mutant compared to the case of co-expression of wild-type version of hVRK3. On the contrary, localization of CLOCK is not affected by hVRK3-NLS mutant. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). Counted numbers of cells are mCRY1 hVRK3, 320; mCRY1 hVRK3-NLSmut, 292; mPER2 hVRK3, 385; mPER2 hVRK3-hVRK3-NLSmut, 235; CLOCK hVRK3, 83; CLOCK hVRK3-NLSmut, 86; BMAL1 VRK3, 147; BMAL1 hVRK3-NLSmut, 144.
38
D. PRKRA
1. hPRKRA는 mPER2 특이적으로 세포 내 위치를 변화시키고, mCRY1/2와
mPER2, CLOCK, BMAL1의 양을 증가시킨다.
HEK 293T 세포에서 mPER2를 발현시켰을 때 70%의 세포가 핵에 위치하였
다. 이러한 mPER2의 분포가 hPRKRA와 같이 발현시키면 핵 (N)에 존재하는 세 포가 약 40%, 핵과 세포질 (N+C)이 약 40%, 그리고 세포질 (C)에만 존재하는 세 포가 약 20%이다 (Figure 10). hPRKRA가 mCRY1과 BMAL1, CLOCK, mSTAT3의 세 포 내 위치는 바꾸진 않았지만 mPER2를 포함하여 mCRY1, BMAL1, CKOCK, mSTAT3 단백질들의 형광 시그널이 강한 것을 확인했다. 즉 단백질의 양적인 변
화가 동반되는 것을 의미한다. 이를 좀 더 정확하게 정량적으로 확인하기 위해 western blot을 수행한 결과, hPRKRA와 함께 발현시킬 경우 mCRY1/2, mPER2,
39
Figure 10 Co-expression of hPRKRA altered subcellular localization of mPER2 in HEK 293T cells. A, HEK 293T cells were transfected with mcry1-egfp,
pUB6(A)-mper2-egfp, pUB6(A)-bmal1-egfp, pUB6(A)-clock-egfp or pUB6(A)-mstat3-egfp in combination with either pUB6-rfp (CTRL) or with pLP-rfp-hprkra, as indicated on the left side of panel. Hoechst stained nuclei were depicted as blue. B, Quantification of subcellular localization of clock proteins and mSTAT3. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). Counted cell numbers for mCRY1 CTRL, 75; mCRY1 hPRKRA, 149; mPER2 CTRL, 203; mPER2 hPRKRA, 447; BMAL1 CTRL, 64; BMAL1 hPRKRA, 182; CLOCK CTRL, 70; CLOCK hPRKRA, 90; mSTAT3 CTRL, 182; mSTAT3 hPRKRA, 313.
40
Figure 11 Clock proteins level is increased by co-expression of hPRKRA. HEK 293T
cells were transfected with 600ng of mcry1/2, mper2, pCMV-HA-clock, or pCMV-HA-bmal1 in combination with 600ng of either rfp-V5 or pUB6-hprkra-V5. shown are the presence (+) or absence (-).
41
2. mPER2와 hPRKRA는 결합하며 hPRKRA는 mPER2 단백질의 인산화를 증
가시킨다.
mPER2가 hPRKRA가 함께 발현되었을 때 SDS-PAGE 상에서 running
mobility가 느린 isoform이 관찰되었기에 (Figure 11A), 이를 확인해 보기 위하여
hprkra와 mper2를 co-transfection 한 후 시간에 따른 변화를 western blot analysis를 사용하였다. hPRKRA가 함께 발현되었을 때 mPER2의 양이 증가하고 running mobility가 늦은 isoform이 관찰되었다 (Figure 12A). 이 isoform이 인산화된 PER2
의 isoform인지 확인해 보기 위해 IP로 mPER2를 분리한 후 λPhosphatase를 처리 하였다 (Figure 12B). 그 결과 running mobility가 증가하는 것을 확인하였으며 이것 은 hPRKRA가 발현되었을 때 mPER2의 인산화가 증가하는 것을 의미한다. 또한 mPER2와 hPRKRA를 함께 발현시킨 세포를 co-immunoprecipitation (co-IP)를 수행
42
Figure 12 Expression of hPRKRA induced phosphoylation of mPER2 via interaction in HEK 293T cells. HEK 293T cells were transiently transfected with 600 ng of
pCMV-HA-mper2 in combination with 600 ng of pUB6(A)-rfp-V5 or pUB6(A)-hprkra-V5 respectively. Cell were harvested at 12h, 24h, 36h (A), or 24h (B, C) after transfection. Protein extracts were either directly analyzed by immunoblotting (A) or proceeded to immunoprecipitation (IP) followed by immunoblotting (B, C). B, Protein extract were incubated with anti-HA (12CA5) antibodies and immune complexes recovered by centrifugation. Immune complexes were incubated in the presence (+) or absence (-) of lambda protein phosphatase (λPPase). Following incubation, the mixtures were subjected to immunoblotting in the presence of anti-HA (3F10) antibodies. Note that the appearnce of slowly running mobility isoforms of mPER2 is increased with co-expression of hPRKRA. C, Protein extracts were incubated with indicated antibodies (top of the panel) and immune complexes were anlayzed by immunoblotting.
43
3. CG6866 -prkra Drosophila homolog- knockdown 초파리는 rhythmicity가 낮다.
hprkra의 초파리 homolog는 loquacious (CG6866)이다. hPRKRA가 생체시계의 작동에 영향을 미치는지를 in vivo에서 확인하고자 초파리에서 loquacious의 발현 을 변화시키고 생체리듬을 측정해보았다. Binary GAL4-UAS system (Brand and perrimon, 1993)을 이용하여 loquacious의 발현을 tim 발현 세포에서만 저하시켰다.
VDRC (Vienna Drosophila Resource Center)로부터 획득한 CG6866 RNAi 초파리
(V108358)를 tim(UAS)-Gal4 초파리와 교배시킨 후 자손의 생체리듬을 분석하였다.
이 때 대조군은 tim(UAS)-Gal4 초파리와 w1118
초파리를 교배시킨 후 얻은 자손을
사용하였다. 25℃에서 12h light : 12h dark에 4일 동안 적응 시킨 후 불을 꺼 계속 어둠에 7일 동안 두면서 Drosophila Activity Monitoring system을 이용하여 초파리의 운동량 (locomotion)을 측정하였다. 결과는 FassX software를 통해 분석하였다. 대 조군 초파리는 각각 rhythmicity가 62.1%, 100% 인데 비해 CG6866 loquacious를 knockdown한 초파리의 rhythmicity는 9.4%, 35.7%로 매우 낮아지는 것을 확인하였
다 (Table 4). 이것은 locomotor activity를 보여주는 eduction graph에서도 볼 수 있었 다 (Figure 13). tim>dicer2,CG6866 RNAi (V108358) (A)는 tim>dicer2 (B) 보다 activity 가 떨어지고, 시간이 지남에 따라 morning peak (ZT0)과 evening peak (ZT12)이 사라 지면서 arrhythmic해지는 것을 볼 수 있다. tim>dicer2,CG6866 RNAi (V22453) (C) 역 시 대조군인 tim>dicer2 (D) 보다 시간이 지남에 따라 arrhythmic해지면서 evening peak (ZT12)이 밀려 period가 약 한 시간 길어지는 것을 볼 수 있다.
44
45
Figure 13 The daily distribution of locomotor activity in loquacious knockdown flies.
Adult male files for a given genotype were entrained in 12L:12D light dark cycle (LD) for 4 days and maintained in constant dark condition (DD) at 25℃. Each panel represents the average activity of flies. Each vertical bar represents relative activity levels during a 15min bin. White horizontal bar represent the light-on phase, black horizontal bar represent the light-off phase, and gray horizontal bar represent subjective day phase.
46
E. UCKL1, NEK2, CKMT1B, ETNK1, MST1R
CRY1의 세포 내 위치를 조절하는 또 다른 인산화 효소는 hUCKL1
(uridine-cytidine kinase 1-like 1)이다. hUCKL1은 mPER2, BMAL1과 같은 생체시계 단백질이 나 mSTAT3와 같은 일반적인 단백질의 세포 내 위치 변화에는 관여하지 않고 mCRY1의 위치만을 변화시켰다 (Figure 14). 앞서 언급했듯이 mCRY1을 HEK 293T
세포에서 과 발현 시키면 대부분 핵 안에 존재하지만 hUCKL1과 함께 발현시키 면 핵 (N)에 약 65%, 핵과 세포질 (N+C)에 약 35% 존재하는 것을 볼 수 있다. hUCKL1에 의해 mPER2와 BMAL1, mSTAT3의 위치는 거의 변하지 않았다.
hNEK2 (NIMA -never in mitosis gene A- related kinase 2)는 HEK 293T 세포에서 핵에 70% 이상 존재하는 mPER2를 핵과 세포질 (N+C)에 40% 이상 존재하게 했 다 (Figure 15). hNEK2와 같이 발현시킨 경우 세포의 모양이 동그랗게 변하는 것 을 관찰하였다. hCKMT1B (creatine kinase, mitochondrial 1B)는 mPER2의 세포 내 위 치를 핵과 세포질 (N+C)에 반 이상 머물게 했다. hETNK1 (ethanolamine kinase 1)과 hMST1R (macrophage stimulating 1 receptor) 역시 mPER2의 세포 내 위치를 변화시 켰다. mPER2가 hETNK1과 발현되면 핵 (N)에 70% 이상으로 관찰되었던 mPER2 가 세포질 (C)이 70% 이상, 핵과 세포질 (N+C)에 20% 이상으로 관찰되었다. hMST1R의 경우 약 45%의 mPER2가 핵 (N)에, 약 40%의 mPER2가 핵과 세포질
47
Figure 14 hUCKL1 alters the subcellular localization of mCRY1. A, HEK 293T cells
were transiently transfected with mcry1-egfp, mper2-egfp, pUB6(A)-bmal1-egfp pUB6(A)-mstat3-egfp in combination with either pUB6-rfp (CTRL) or pLP-rfp-huckl, as indicated on the left side of panel. Hoechst stained nuclei were depicted as blue. B, Quantification of subcellular localization of clock proteins. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). Counted cell numbers for mCRY1 CTRL, 75; mCRY1 hUCKL1, 295; mPER2 CTRL, 203; mPER2 hUCKL1, 176; BMAL1 CTRL, 64; BMAL1 hUCKL1, 116; mSTAT3 CTRL, 182; mSTAT3 hUCKL1, 150.
48
Figure 15 Co-expression of hNEK2, hCKMT1B, hETNK1, or hMST1R altered the subcellular localization of mPER2 in HEK 293T cells. A, HEK 293T cells were
transfected with pUB6(A)-mper2-egfp in combination with either pUB6-rfp, pLP-rfp-hnek2, pLP-rfp-hckmt1b, pLP-rfp-hetnk1 or pLP-rfp-hmst1r, as indicated on the left side of panel. Hoechst stained nuclei were depicted as blue. B, Quantification of subcellular localization of mPER2. N denote for percent of cells localized in nucleus (black); N+C denote for percent of cells localized in both in nucleus and cytoplasm (gray); C denotes for percent of cells localized in cytoplasm (white). Counted cell numbers for CTRL, 110; hNEK2, 236; hCKMT1B, 453; hETNK1, 166; hMST1R, 291.
49
Ⅳ. 고찰
지구상에 존재하는 모든 생명체는 내제되어있는 정교한 생체시계를 지닌다. 포유류에서 생체시계를 조절하는 핵심 중추는 시상하부의 SCN (suprachiasmatic nuclei)으로, 자기조절의 음성 되먹임 전사활동 (autoregulatory negative-feedback loop)
에 의해 약 24시간의 리듬을 가지게 된다 (Lowrey and Takahashi 2004; Takahashi et al., 2008). 이러한 되먹임 고리는 CLOCK/BMAL1의 이량체가 cryptochrome (cry1,
cry2)과 period (per1, per2, per3) 유전자의 E-box에 붙어 이들의 전사를 활성화 시 킴으로 시작된다. 세포질에 축적된 CRY와 PER 단백질들은 핵으로 이동하고 CLOCK과 BMAL1에 작용하여 이들의 전사를 억제시킨다. 생체주기를 정확하게 조절하기 위해서는 생체시계 단백질들의 양과 활성뿐 만이 아니라 인산화와 같은 번역 후 조절 (post-translational regulation)이 중요하다. 이전 연구들로부터, 몇 개의 생체시계 단백질의 세포 내 분포를 조절하는 인산화 효소들이 알려져 있다. 예를 들어 mPER2 단백질의 세포 내 위치를 조절하는 것 으로 알려진 casein kinase I ε (CK1ε)을 일시적으로 mPER2-EGFP와 동시에 발현 시 켰을 때 Figure 4B에서 보여지는 것과 같이 세포 내 위치가 변하였다. 인산화 효 소를 발현하지 않은 경우에는 mPER2-EGFP는 핵 안에 존재하였으나 인산화 효 소를 함께 발현 시켰을 경우에는 mPER2-EGFP가 핵 안으로 이동하지 못하고 세 포질에 머무르는 것을 확인할 수 있었다. 전체 인간의 인산화 효소는 533개 이며,
50
그 중 지금까지 알려진 생체시계 단백질을 조절하는 인산화 효소는 5개다. 따라 서 더 많은 인산화 효소가 존재할 것이라고 생각하여 본 연구를 진행하였다.
본 연구에서는 human kinome expression library를 구축하고 새로운 생체시계 조절 인산화 효소를 발굴하고자 했다. mSTAT3를 포함한 모든 생체시계 단백질들 은 마우스 단백질을 사용했으며, 포유류 간의 생체시계 단백질들의 상동성은 매 우 높아 사람의 생체시계에서도 유사한 작동기전을 나타낼 것이라고 생각된다. 현재 mCRY1의 인산화 효소 스크리닝은 118개를 완료하였고 그 결과 hVRK3, hUCKL1을 발굴하였다. 또한 mPER2 인산화 효소 스크리닝은 82개의 인산화 효
소를 완료하였으며 그 결과 hNEK2, hCKMT1B, hETNK1, hMST1R, hPRKRA 그리고 hVRK3을 찾았다.
hVRK3는 HEK 293T 세포와 NIH 3T3세포에서 생체시계 단백질들 중
CLOCK을 제외한 mCRY1/2, mPER2, 그리고 BMAL1의 세포 내 위치를 변화시켰
다. 이전에 시행된 연구로부터 hVRK3와 hVRK1,2 모두 Ran-GTPase와 결합하는 것으로 알려져 있다 (Sanz-García et al., 2008). Ran-GTPase는 핵 이동에 관여하는 small G protein으로 핵 이동 시 결합하여 중요하게 작용하는 것으로 알려져 있다
(Stewart, 2007; Sekimoto and Yoneda, 2012). 이에 hVRK3가 생체시계 단백질들뿐 만
아니라 다른 일반적인 단백질과 상호작용하여 위치를 변화시킬 수도 있다고 생 각했고 핵 이동을 한다고 잘 알려진 mSTAT3도 확인해보았다 (Reich, 2013). 흥미 롭게도 hVRK3가 mSTAT3 단백질의 위치를 바꾸지 않았다. hVRK3가 위치를 변화