허베이 스피리트 유류사고 이후 태안 조간대 해역 저서 규조류의 군집 변동
박종규
1*·김소연
2·김병석
2·최충현
2·국현근
21군산대학교 해양생명응용과학부, 2군산대학교 해양학과
Change of Benthic Diatom Community in Taean Tidal Flats
after Hebei Spirit Oil Spill
Jong-Gyu Park
1*, So-Yeon Kim
2, Byound-Seok Kim
2, Chung-Hyun Choi
2and Hyeon-Geun Kook
21Department of Marine Life and Applied Sciences, College of Ocean Science and Engineering,
Kunsan National University, Gunsan 54150, Korea
2Department of Oceanography, College of Ocean Science and Engineering, Kunsan National University,
Gunsan 54150, Korea 요 약 :2007년 겨울 태안 해역에서 발생한 허베니 스피리트호 유류 오염 사고가 조간대 저서 규조류에 미치는 영향을 알아보기 위해 태안해안국립공원의 5개 해역(학암포, 만리포, 파도리, 백사장, 바람아래)에서 10년간 저서 규 조류의 생물량과 군집 변동을 모니터링 하였다. 유류 오염사고 이후 5개 조간대 해역과 해수 중에 미세조류의 엽 록소 농도는 크게 증가하였지만 이후 급격히 감소하여 2012년에 최소치를 보였다. 2014년 이후에는 다시 증가하였 고 최근에는 수중에서 2-3 g L1, 갯벌 해역에서 0.3-0.6 g cm2의농도를 유지하고 있다. 저서 규조류 세포수도 2012년에 최소치를 보인 후 2014년 이후 회복하여 2 × 105 cells cm2 내외를 유지하고 있다. 오염사고 이후 저서 미세조류의 종조성은 오염 해역과 비오염 해역 사이에 크게 차이가 났지만, 2011년 이후 해역간 차이가 줄어들기 시작하여 최근에는 두 해역 간 차이가 거의 없다. 이런 결과는 저서 규조류에 대한 유류 오염의 국지적 영향이 거 의 사라졌음을 의미한다. 주요어 :허베이 스피리트, 유류오염사고, 태안해역, 저서미세조류, 조간대, 규조류
Abstract : To investigate the effects of the Hebei Spirit oil spill on the intertidal benthic diatoms in the Taean Sea area, which occurred in winter 2007, the biomass and community changes of benthic diatoms were monitored in the intertidal zone of Hagampo, Manripo, Padori, Baeksajang and Baramarae of Taean National Park for 10 years. The chlorophyll concentrations of microalgae in the five intertidal zones and seawater increased significantly after the oil spill, but then declined sharply, reaching a minimum in 2012. It has increased again since 2014, and recently maintained the concentration of 2-3 g L1 in seawater and 0.3-0.5 g cm2 on tidal flat every year. The number of benthic diatom cells
also reached a minimum in 2012 and has been recovered since 2014, maintaining around 2 × 105 cells cm2. The species
composition of benthic microalgae varied greatly between polluted and non-polluted areas after oil spill, but since 2011, the differences between two areas have begun to diminish, and has recently become very similar in the entire surveyed area. These results implies that the local impact of oil pollution on benthic diatoms has almost disappeared.
Key words : Hebei Spirit, Oil Spill, Taean, Benthic Microalgae, Tidal Flats, Diatoms
서
론
지난 2007년 12월 7일 충청남도 태안군 만리포 앞 해역에 서는 15만톤급 홍콩 선적 유조선, Hebei Spirit호가 원유 10,800여 톤을 유출하는 대형 해양 유류오염 사고가 발생하 였다. 사고 발생 해역은 태안해안국립공원이 위치한 해역으 로 서해안의 특성상 조수 간만의 차가 크고 넓은 조간대가 형성되어 있으며 태안반도와 안면도의 해안선을 따라 *Corresponding author E-mail: [email protected][총설]
326.574 km2에 이르는 넓은 지역에 모래 갯벌이 펼쳐져 아 름다운 풍광을 자랑한다. 유조선에서 흘러 나온 원유는 겨 울철 굳은 날씨의 영향을 받아 사고 해역 주변으로 급격히 퍼져나갔고 태안해안국립공원의 조간대 해역은 물론, 해안 선, 해안사구, 해빈 등을 심각하게 오염시켰다. 이러한 기름 오염의 충격적인 피해 현황은 언론을 통해 대대적으로 보도 되어 국립공원 해역의 생태 환경 파괴와 피해 복구에 대한 관심은 지역 주민뿐만 아니라 전국민에게 확대되는 계기가 되었다. 해양의 유류 사고가 해양생태계에 미치는 영향이나 피해 에 대한 연구 결과는 다양한 관점에서 보고되었다(Bautista and Rahman 2016a, b; Dunnet et al. 1982; Ehrenhauser et al. 2014; Nance et al. 2015; Tidwell et al. 2015, Wolfaardt et al. 2009). 유조선 사고 등을 통해 해양으로 유입된 기름 은 유막을 형성하여 해수의 용존산소량을 떨어뜨리고 태양 빛을 차단하여 식물플랑크톤의 일차생산력을 떨어뜨린다. 갯벌이나 모래, 자갈 등 저서 기질을 덮어 저서생물을 질식 사하게 하고 해조류 엽상체에 붙어 대사작용을 방해하기도 한다. 대형생물에 붙어 운동성을 저하시키고, 해양 생물에 유독 피해를 주기도 하며 새의 깃털이나 포유류의 털에 침 투하여 단열 능력을 감소시키고 부력을 떨어뜨린다. 체내에 들어가면 탈수를 일으키고 소화기를 손상시키며 폐나 간으 로 들어가 죽음에 이르게 할 수도 있다. 해양생태계에서 규조류는 다양한 미세조류 중에서 가장 우세한 분류군으로 해양 일차생산량의 약 40%를 차지하는 것으로 알려져 있다(Berger and Wefer 1991). 연안 저서생태 계에서는 미세 진핵생물과 남세균류를 포함하여 부유 규조 류와 부착 또는 저서 규조류가 모두 관찰되며, 그 중에서 가장 종이 많고 양적으로 풍부한 분류군이 저서 규조류이다 (Admiraal 1984). 이들은 조간대 갯벌과 조하대의 주요 일차 생산자로서 어패류와 갑각류 등의 먹이원이 된다(Cahoon 1999; Cahoon et al. 1993; Miller et al. 1996). Martin et al.(1987)와 Longhurst et al.(1995)는 전체 해양 생산량의 0.7%와 대륙붕 생산량의 2.4-3.7%가 저서 미세조류에 의한 것으로 추정하였고 Schwinghamer et al.(1986)은 Fundy 만 의 Pecks Cove에서 전체 생산력의 26~64%가 저서 미세조 류에 의한 것으로 보고한 바 있다. 저서 규조류는 퇴적물 표면에 분포하기 때문에 퇴적물과 물기둥 사이의 영양염 교 환을 조절하는 역할을 하며(Sundbftck et al. 1991; Rizzo et al. 1992; Underwood and Kromkamp 1999), 점액질 필름을 분비함으로써 퇴적 안정화와 같은 갯벌의 생태적 기능을 보 존하는 데에도 크게 기여한다(Miller et al. 1996; Holland et al. 1974; Paterson et al. 1990; Delgado et al. 1991). 이러한 기능으로 인해 저서 규조류는 오래 전부터 그 생태와 다양 성에 대해 많은 관심을 받아왔다(Hustedt 1955; Smyth 1955; Round 1957, 1960). 저서 미세조류는 담수의 생태학적 상태를 평가하기 위해 유럽 연합 수질 기본 지침 2000/60(W.F.D.)에서 권장하는 주 요 생물 그룹이다(CEC 2000). 이중에서 규조류는 저서 미세 조류의 주요 구성 생물로서 수생 생태계를 모니터링 하는 가장 중요한 조류 그룹 중 하나이다(Kelly et al. 1998; Mancini 2005; King et al. 2006). 이들의 생물학적, 생태학 적 특성은 수질의 좋은 지표가 되며, 유수(Prygiel et al. 1999)와 정수, 특히 호수(Kitner & Poulí ková 2003; DeNicola & Eyto 2004; Denys 2004)의 질을 평가하는 데에 널리 사 용되고 있다. 규조류는 얕은 바닷물에서 채취하기 쉬워서 실용적으로 사용할 수 있는 장점이 있으며(King et al. 2000), 환경 조건의 변화와 인간에 의한 인위적 압력에 빠르 게 반응하기 때문에 고생물학적 연구에 종종 사용되기도 한 다(Deny 2003; Bennion et al. 2004; Bradshaw et al. 2006). 이처럼 규조류는 강과 호수에서 지표로 자주 사용되지만 갯 벌을 포함한 습지 시스템의 저서 규조류에 대해서는 거의 알려지지 않았다. 미국(Pan & Stevenson 1996; Gaiser et al. 2005; Wang et al. 2006; Zheng & Stevenson 2006)과 호주 (Gell et al. 2002)에서 습지의 생태학적 상태에 대한 지표로 부착 미세조류를 연구한 적이 있다. 국내 저서 규조류에 대 한 연구는 주로 담수 규조류에 중점을 두고 있으며 해양 저 서 규조류에 대해서는 분류학적 연구가 제한적으로 이루어 지고 있다(An et al. 2017; Kim et al. 2017, 2019; Gastineaua 2019). 본 연구는 Hebei Spirit호의 유류유출 사고 이후 해양 환 경과 생태계에 미친 영향을 파악하는 것이 목적이다. 이를 위해 환경 변화를 예민하게 반영하는 특성이 있고 생태계 먹이사슬을 따라 연속적으로 2차, 3차 피해를 줄 수 있는 저서 규조류에 대하여 그 현존량과 종조성 변화를 장기간에 걸쳐 조사하였다.
재료 및 방법
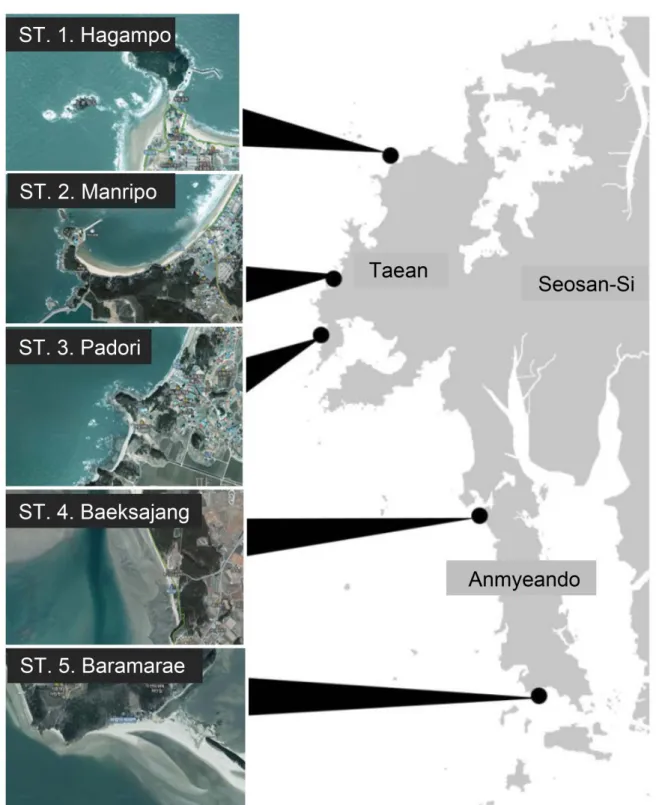
1. 조사지역 및 분석방법 2015년부터 2018년까지 계절별로 연간 4회에 걸쳐 유류 유출 사고의 주요 피해 지역인 학암포, 만리포, 파도리 3개 조간대 해역과 유류 유출 피해가 적은 백사장 해역과 바람 아래 해역의 조간대에서 현장 조사를 수행하였으며(Table 1, Figure 1), 2009년부터 2014년까지 결과는 기존 보고서 자료 를 이용하였다(국립공원관리공단 2009~2014). 2. 수온과 염분 해수의 수온과 염분은 휴대용 T-S meter(YSI, Pro 80)로 측정하였다. cêTable 1. Administrative districts, latitude and longitude of the survey area
Station Location Latitude Longitude 1 2 3 4 5 Hagampo Manripo Padori Baeksajang Baramarae 36o 53′ 50.4″ 36o 47′ 10.5″ 36o 44′ 15.0″ 36o 34′ 33.0″ 36o 24′ 40.9″ 126o 12′ 10.3″ 126o 08′ 22.1″ 126o 07′ 49.7″ 126o 18′ 38.3″ 126o 22′ 20.2″
3. 엽록소-a 저서 미세조류의 엽록소-a 농도를 측정하기 위해 저서 기 질의 특성에 따라 학암포, 만리포, 백사장, 바람아래에서 저 서 퇴적물을 채집하였고 파도리에서는 작은 자갈 시료를 채 집하였다(Figure 1). 저서 미세조류와 해수 내 식물플랑크톤 의 엽록소 농도를 비교하기 위해 해수 1 L도 채집하였다. 저 서 미세조류의 엽록소 농도 측정을 위해 각 정점에서 채집 한 퇴적물 시료 중 일정한 양을 취하여 아세톤 15 mL을 넣 고 냉암소에서 24시간 동안 추출하였다. 파도리에서 채집한 자갈 시료의 경우에는 시료가 충분히 잠길 만큼 아세톤을 넣고 냉암소에서 24시간 동안 추출하였다. 추출한 엽록소는 형광측정기(TU-10, Fluorometer, USA)로 형광량을 측정한 후, 단위 면적 당 엽록소-a 농도(g cm2)로 환산하였다. 해
수 식물플랑크톤의 엽록소 농도의 경우에는 현장에서 채수 한 시료를 직경 45 mm GF/F 여과지로 여과하여 여과지를 20 mL vial에 담아 빛을 차단하여 냉동 보관한 후, 해양환경
공정시험기준(해양수산부 2013)의 아세톤 추출법에 따라 냉 동 여과지를 상온에서 녹인 다음, 90% 아세톤 15 mL로 24 시간 동안 추출하고 형광측정기로 측정하였다. 식물플랑크 톤은 단위 부피당 엽록소-a 농도(g L1)로 환산하였다. 4. 저서 규조류 분석 저서 규조류의 정량분석을 위하여 학암포, 만리포, 백사장, 바람아래에서는 저서 퇴적물을 채집하고 파도리에서는 작은 자갈 시료를 채집한 후 실험실에서 영구슬라이드를 제작하 였다. 단위 면적당 세포수로 환산하기 위해 자갈 시료의 표 면적과 퇴적물 시료의 건중량과 표면적을 측정하였으며 광 학현미경 ×1,000 배율로 종 동정과 계수를 하였다. 영구 슬라이드는 Witkowski et al.(2010)를 참고하여 돌말 껍질(frustule)의 미세구조 관찰이 가능하도록 세척 과정을 거쳐 제작하였다. 슬라이드 제작 순서는 아래와 같다. 파도 리 시료의 경우에는 자갈 전체 표면을 칼로 긁어 부착 규조 류 시료를 채집한 후 (i) 과정을 생략하고 슬라이드를 제작 하였다. (i) 비커에 퇴적물 시료를 담아 무게를 재고 110~130℃에 서 말린 후 건중량을 측정하였다. (ii) 10% HCL 20 mL를 넣고 2-3시간 대기한 후, (iii) 증류수를 가득 채우고 12시간 동안 규조류를 침전시켜 농축하고 상등액을 최소 2번 이상 제거하였으며, (iv) 30% 과산화수소를 채우고 250~300℃에 2~12시간 동안 가열하였다. (v) 시료에 증류수 100 mL를 다 시 넣고 10% 염화암모늄 8방울 떨어뜨린 다음, (vi) petri dish에 증류수로 커버글라스(1.8×1.8 cm2)를 고정시키고 건조 후 커버슬라이드(7.6×2.6 cm2)가 잠길 정도로 골고루 뿌려주 고 완전히 건조시켰으며, (vii) 핀셋으로 커버슬라이드 2개를 떼어 슬라이드 글라스에 올려 핫플레이트를 이용하여 2-3분 가열하였으며, (viii) Naphrax 한 방울을 떨어뜨려 커버슬라 이드로 덮은 후 150℃로 짧게 가열하고 기포가 생기면 제거 한 후 (ix) 봉입제가 흘러내리지 않도록 서늘한 곳에 수평으 로 보관하였다. 위 과정을 거쳐 제작한 영구슬라이드를 ×1,000배 광학현 미경(Axioimager, A2, Zeiss, Gmbh 37081, Gottigen, Germany) 으로 관찰하고 현미경 부착 사진기(eXcope X3)로 촬영하여 Witkowski et al.(2000), Hustedt and Jensen(1985) 등을 참조 하여 동정하였다.
결과 및 고찰
1. 출현 종 조성 2007년 12월 26-28일 예비조사 수행 후, 2008년 1월~12월 에 출현한 저서 규조류는 29속 44종으로 매우 빈약하였다. 동계, 춘계, 하계 3계절만 조사한 2009년과 2010년에는 각 각 36속 124종, 51속 210종이 출현하였다. 각 계절별로 1회 씩 연간 4회 조사한 2011년부터 2014년에는 52속 111종, 53 속 153종, 47속 112종, 42속 118종이 출현하였고, 2015년부 터 2018년에는 각각 47속 253종, 46속 160종, 37속 113종, 42속 127종이 출현하였다. 출현 종 수는 10년간 뚜렷한 변화를 보이지 않았다. 2010 년과 2015년에 200종 이상 출현하였지만, 지속되지는 않았 다. 유류 오염 이후 뚜렷한 변화가 보이지 않는 것은 연구 자의 연속성이 없었기 때문인 것으로 보인다. 지난 10년간 연구자가 3번 교체되었으며, 2009-2010년, 2011-2014년, 2015-2018년 결과는 각각 다른 연구자들의 결과이다. 2010 년과 2015년은 모두 연구자가 변경되어 종 분류와 분석 방 법에 변화가 생긴 해다. 최근 들어 규조류 분류에 전자현미 경과 유전자 염기서열 분석 방법이 도입됨으로써 종의 수가 급격히 늘고 기존 속에서 새로운 속이 분화되는 속도도 크 게 증가하고 있다. 2009-2010년에는 광학현미경으로 분류하 였으나, 2011년 이후 전자현미경의 도움을 받은 것도 영향 을 미쳤을 것이다. 이런 차이를 감안하면 출현 종 수는 연 구자에 따라 조금씩 다를 수 있다. 다만, 각 연구자의 연구 기간 내에는 일관성을 유지하였을 것으로 보인다. 2009년에 5종 이상 출현한 속은 Cocconeis(15종), Amphora, Navicula, Nitzschia(이상 11종), Achnanthes(10종), Fallacia (8종), Planothidium(7종), Diploneis, Fragilaria(이상 6종)이 었고, 2010년에는 Navicula(32종), Amphora(30종), Fragilaria (16종), Cocconeis(15종), Nitzschia(14종), Diploneis(13종), Achnanthes, Fallacia(이상 7종), Hatzschia(6종), Ophepora, Planothidium(이상 5종)이었다(국립공원관리공단 국립공원연 구원 2009a, b, 2010).2011년-2014년에는 5종 이상 출현한 속의 수가 줄어들어 2011년에 Navicula(15종), Thalassiosira(10종), Nitzschia(9종), Amphora, Cocconeis(이상 5종), 2012년에 Nitzschia(19종), Navicula, Amphora(이상 13종), Achnanthes, Thalassiosira(이 상 9종), Cocconeis(8종), Actinocyclus(6종), Tabularia, Planothidium (이상 5종), 2013년에 Nitzschia(17종), Amphora(10종), Navicula (9종), Achnanthes, Thalassiosira(이상 6종), Cocconeis(5종), 2014년에 Nitzschia(17종), Achnanthes(11종), Navicula(9종), Amphora(8종), Cocconeis(7종), Planothidium(6종)이었다(국 립공원관리공단 국립공원연구원 2011~2014).
2015년-2018년에는 5종 이상 출현한 속의 수가 비교적 크 게 증가한다. 2015년에는 Cocconeis(26종), Amphora(19종), Fallacia, Planothidium(이상 18종), Navicula(14종), Nitzschia (12종), Delphineis, Diploneis(이상 11종), Halamphora(9종), Fragilaria(8종), Cocconeiopsis, Opephora, Psammodictyon(이 상 7종), Achnanthes, Biremis, Eunotogramma, Hippodonta, Plagiogrammopsis(이상 6종), Anorthoneis, Plagiogramma(이 상 5종), 2016년에 Cocconeis(19종), Navicula(14종), Amphora, Fallacia(이상 11종), Planothidium(9종), Delphineis, Nitzschia, Plagiogramma(이상 8종), Diploneis(7종), Eunotogramma(5종), 2017년에 Cocconeis(13종), Amphora, Diploneis, Fallacia, Navicula(이상 8종), Halamphora, Planothidium(이상 6종), Nitzschia(5종), 2018년에는 Cocconeis(18종), Navicula(11종), Amphora(8종), Diploneis, Eunotogramma, Fallacia, Halamphora, Nitzschia, Planothidium(이상 6종), Delphineis(5종)이었다(국
립공원관리공단 국립공원연구원 2015~2018).
10종 이상 출현한 속으로 축약하면 2009-2010년에는 Amphora, Cocconeis, Navicula, Nitzschia, 2011-2014년은 Nitzschia(3년), Amphora, Navicula(이상 2년), 2015-2018년은 Cocconeis(4년), Navicula(3년), Amphora, Fallacia(이상 2년)이었다. 이들은 모두 종의 수가 100여종 이상인 것으로 알려져 있으며 최근 새로운 속으로 분화되고 있는 추세이다. 연구 해역의 출현 종은 광학현미경에 의존하여 분류한 종이 많기 때문에 신종 이나 신속이 많이 있을 것으로 판단된다. 각 조사 정점에서 매 조사 시기에 관찰한 저서 규조류의 출현 종 수는 5~64종으로 변화하였고, 미약하게 학암포에서 다른 정점보다 지속적으로 높은 특성을 보였다(Figure 2
Figure 2. Annual change in the mean species number of benthic diatoms observed at each station (upper) and each season. Since no samples were collect during the fall of 2009 and 2010, only three seasons were taken into account.
Figure 3. Annual change in the number of species. ■ indicates all species appearing in the entire survey area per a year, ●, the yearly average of all species emmergent in the entire survey area at each season, and □, the yearly average of species appearing at each site at each survey period. No samples were collect during the fall of 2009 and 2010.
upper). 2015년 이후에는 파도리에서 종 수가 확연하게 적었 지만 이는 조사 방법의 차이 때문이다. 2014년까지는 퇴적 물 시료의 규조류를 분석한 반면, 2015년 이후에는 작은 돌 멩이에 붙어 있는 규조류를 분석하였다. 연간 변화와 계절 별 변화도 뚜렷하지 않았지만, 2015년 이전에는 가을철에 다른 계절보다 조금 더 많은 종이 출현한 것을 볼 수 있다 (Figure 2 lower). 각 계절별로 전체 조사 해역에 출현한 종은 33~149종이 었다. 2015년에 평균 137종으로 가장 많았고, 2011년에 평 균 47종으로 가장 적었다. 연간 조사해역에 출현한 종의 수 를 나타내는 방법은 각 조사 시기에 각 정점에서 출현한 종 수의 연평균, 매 조사 시기별 전체 정점에서 출현한 종 수 의 연평균, 연간 전체 정점에서 출현한 모든 종의 수 등 여 러 가지가 있다. 하지만 각 수치의 장기적인 변화 유형은 뚜렷한 차이 없이 모두 동일한 양상을 보였다(Figure 3). 따 라서 연구자들이 저서 규조류을 연구하는 데에 일관성 있는 조사 방법이나 분석 방법을 유지하는 것이 매우 중요하다는 것을 알 수 있다. 2. 엽록소 농도 유류 오염사고 이후 10년간 5개 조간대 해역의 각 정점에 서 수중 식물플랑크톤의 엽록소 농도는 0.02~19.67 mg L1 의 범위를 보였고, 10년 평균은 3.17 mg L1이었다. 연도별 평균은 유류 오염 사고가 발생하였던 해와 가장 가까운 2009년에 10.83 mg L1으로 가장 높았고 2012년에 0.05 mg L1으로 가장 낮았다(Figure 4 upper). 2007년 12월 처음 유 류 오염 사고가 발생한 직전이나 직후의 엽록소 농도는 알 수 없지만, 가장 근접한 시기인 2009년에는 10년 평균치를 3배 이상 상회하는 10.83 mg L1의 높은 농도를 보였고, 그 이후 매년 급격히 감소하기 시작하여 2012년에는 0.05 mg L1에 이르는 매우 낮은 농도를 보였다. 2013년 이후부터 조 금씩 증가하여 2015년에 3.96 mg L1의 농도를 보였고, 그 이후로 2~3 mg L1를 오르내리고 있다. 조간대 해역의 식물 플랑크톤 생물량은 2009년에 정점에 도달한 이후 2012년 최소치를 거쳐 큰 순환을 마치고 현재는 소규모의 생물량 순환을 겪고 있는 것으로 판단된다. 반면 5개 조사 해역의 조간대 갯벌 기질 내 엽록소 농도 에 대해서는 이러한 변화를 뚜렷하게 확인하기 어렵다. 2011년부터 2014년까지는 갯벌 기질 내 엽록소 농도를 분석 하지 않았기 때문이다. 그러나 분석이 이루어진 연도의 갯 벌 기질 내 엽록소 농도를 보면 전체적인 변동 유형이나 진 폭이 수중 엽록소 농도 변화와 매우 유사하다는 것을 알 수
Figure 4. Annual change in chlorophyll-a concentrations of phytoplankton in water column (upper) and benthic diatoms on tidal flat at each station.
있다(Figure 4 lower). 이런 측면에서 갯벌 기질 내 엽록소 농도도 수중 엽록소 농도와 마찬가지로 2009년에 연평균 1.34 mg cm2의 최고 농도에 도달한 이후 2011년과 2014년 사이에 최소 농도를 거치고 2005년 이후에는 새로운 순환 주기를 맞이하여 0.3-0.6 mg cm2의농도를 유지하고 있는 것으로 추정된다. 수중 엽록소 농도와 해저 갯벌 기질 내 엽록소 농도를 비 교하는 것은 쉽지 않다. 수중 엽록소 농도는 단위 부피 속 에 들어있는 양(mg L1)이고 갯벌 기질 내 엽록소 농도는 단 위 면적에 들어 있는 양(mg cm2)으로 서로 차원이 다르기 때문이다. 하지만 특정 부피의 엽록소 양이 해당 면적을 수 직으로 적분한 값으로 생각하면 비교가 가능하다. 예를 들 어 1 L를 1 cm × 1 cm × 10 m로 생각하면 1 mg L1은 단면 적 1 cm2의 10 m 물기둥 안에 1 mg의 엽록소가 들어 있는 것이다. 만약 이때 갯벌 기질 속 엽록소 농도가 1 mg cm2 이라면, 갯벌 기질 속 엽록소 양은 수중 10 m 안에 들어 있 는 엽록소 양과 동일한 양이 들어 있는 것이다. 이런 방식 으로 조사 해역의 갯벌 기질 내 엽록소 양을 같은 양의 엽 록소가 들어 있는 수중의 물기둥 높이로 변환한 것이 Table 2이다. 계산식은 아래와 같다. D = 10 × 여기에서 D는 물기둥의 높이(m), Chls는 갯벌 기질 내 엽 Chls Chlw
---Figure 5. Annual change in the abundances of benthic diatoms observed at each station (upper) and each season. Since no samples were collect during the fall of 2009 and 2010, only three seasons were taken into account.
Table 2. Water column thickness containing equivalent content of chlorophyll-a in the sediment. The amount of chlorophyll-a in the sediment is correspond to the chlorophyll amount in the water column of the thickness.
(unit: m) Station 2009 2010 2015 2016 2017 2018 Mean 1 2 3 4 5 0.22 2.61 3.18 0.92 0.48 0.81 3.04 2.28 1.31 2.65 1.08 0.52 2.05 0.79 0.84 2.00 2.02 2.02 1.00 1.53 6.55 3.33 5.87 5.86 6.07 1.82 1.90 7.37 2.09 2.32 2.08 2.24 3.79 1.99 2.32 Mean 1.48 2.0 2 1.0 6 1.71 5.54 3.10 2.48
록소 농도(mg cm2), Chl w는 수중 엽록소 농도(mg L1)이다. 표에 따르면 조사 해역의 갯벌 기질 내 엽록소 양은 평균 적으로 해수 약 2.5m 안에 들어 있는 엽록소 양에 해당하였 다. 자갈 위 엽록소 농도를 측정한 파도리를 제외하면 평균 적으로 대략 해수 2.1m 안에 들어 있는 엽록소 양이었다. 3. 퇴적물 내 저서 규조류 세포수 유류 오염사고 이후 10년간 5개 조간대 해역의 각 정점에 서 계수한 갯벌 기질 내 저서 규조류의 연 평균 세포수는 312~411,460 cells cm2의 범위를 보였고, 10년 평균은 147,246
cells cm2이었다(Figure 5 upper). 공간적으로는 유류 오염
사고가 발생했던 만리포와 학암포에서 다른 정점보다 많은 편이었지만 뚜렷한 차이는 없었다. 파도리는 자갈 위에 부 착 규조류를 계수하였기 때문에 타 정점에 비해 현격하게 적은 양을 보였다. 각 계절에 5개 조간대 갯벌 해역에 출현 한 저서 규조류의 평균 세포수는 1,105~352,500 cells cm2 의 범위를 보였고 가을철에 가장 풍부하였다(Figure 5 lower). 하지만 계절 간 차이도 뚜렷한 것은 아니었다. 안타깝게도 유류 오염 사고가 발생한 직후인 2008~2010 년에는 조간대 갯벌 기질 내 저서 규조류의 세포수를 정량 적으로 측정하지 않은 것으로 보인다. 당시 보고서에는 출 현 종간의 상대비만 있고 절대적인 세포수는 제시되지 않았 기 때문이다. 이 기간을 제외하면 저서 규조류의 세포수는 전체적인 변동 양상이 수중 엽록소 농도와 유사하게 변화하 였다. 2011~2012년에 세포수가 가장 적었고 2015년까지 조 금씩 증가하여 최대치를 보인 후 2 × 105 cells cm2를 기준
Table 3. Seasonal changes in the dominant species of benthic diatoms in the research area, 2009-2010.
Season Station Upper Intertidal Zone % Mid-intertidal Zone % Lower Intertidal Zone %
2009 Spring
1 Nitzschia miserabilis 18.8 Nitzschia miserabilis 31.8 Achnanthes curvirostrum 29.8 Rhaphoneis amphiceros 18.8
2 Cocconeiopsis kantsiensis 53.9 Cocconeiopsis kantsiensis 68.9 Cocconeis nugalas 48.2 3 Navicula sp. 48.6 Navicula cf. leptoloba 36.7 Navicula cf. leptoloba 94.5 4 Catenula adhaerens 41.2 Catenula adhaerens 29.8 Catenula adhaerens 43.5 5 Cocconeis scutellum v. parva 25.2 Cocconeis scutellum v. parva 38.3 Cocconeis scutellum v. parva 23.9
2009 Summer
1 Cocconeiopsis kantsiensis 42.0 Planothidium delicatulum 30.6 Halamphora wisei 52.4 Navicula sp. 39.1
2 Cocconeiopsis kantsiensis 97.1 Planothidium delicatulum 20.7 Cocconeis scutellum v. parva 16.9 3 Navicula sp. 54.3 Cocconeiopsis kantsiensis 31.9 Haslea duerrenbergiana 65.7 4 Seminavis barbarae 46.5 Catenula adhaerens 41.0 Catenula adhaerens 55.8 5 Amphicocconeis disculoides 30.4 Achnanthes fogedii 22.5 Planothidium engelbrechtii 26.5
Fallacia cryptolyra 22.5
2010 Winter
1 Planothidium delicatulum 22.0 Plagiogramma staurophorum 14.3 Cocconeis scutellum 18.3 2 Navicula sp. 31.7 Cocconeis scutellum v. parva 22.1 Navicula sp. 33.3
Prestauroneis protracta 31.0
3 Rhaphoneis sp. 17.5 Cymatosira belgica 15.7 Diploneis papula 15.6 4 Cymatosira belgica 21.2 Catenula adhaerens 20.4 Catenula adhaerens 20.6 Plagiogrammopsis crawfordii 16.4 5 Navicula phylleptosoma 61.0 Cocconeis distans 13.3 Cymatosira belgica 16.7 Planothidium delicatulum 24.0 Fragilaria amicorum 13.3 Cocconeis distans 13.6
2010 Spring
1 Planothidium delicatulum 18.3 Planothidium delicatulum 33.8 Planothidium delicatulum 15.1 2 Planothidium delicatulum 31.0 Planothidium delicatulum 27.6 Planothidium delicatulum 31.8 3 Navicula rolandii 28.2 Navicula rolandii 41.1 Navicula rolandii 33.0 4 Navicula sp. 63.9 Catenula adhaerens 20.4 Catenula adhaerens 19.9 5 Navicula sp. 19.6 Planothidium delicatulum 18.3 Planothidium delicatulum 22.0 Amphora sp. 17.9 Catenula adhaerens 14.6 Catenula adhaerens 14.3
2010 Summer
1 Amphora sp. 20.1 Navicula bipustulata 22.9
2 Cocconeiopsis kantsiensis 89.1 Catenula adhaerens 33.0 Cymatosira belgica 28.8 3 Catenula adhaerens 22.2 Cymatosira belgica 15.8 Diploneis decipiens var. parallela 15.7
Hyaloneis hyalinum 16.7
4 Navicula borowkae 26.3 Achnanthes curvirostrum 27.5 Catenula adhaerens 24.9 5 Anorthoneis vortex 39.2 Fragilaria gedanensis 13.7 Navicula bipustulata 11.8
으로 다시 소규모의 감소와 증가를 오르내리는 과정을 겪고 있다(Figure 5). 조간대 갯벌 기질 내 저서 규조류의 세포수와 엽록소 농 도는 그 상관 관계가 높은 편은 아니었다. 세포수가 2012년 이후 2015년까지 증가하고 그 이후 조금 감소한 반면 엽록 소 농도는 2017년까지 증가하다가 감소하는 등 소규모 변동 에 차이가 있었다. 이와 같은 세포수 기반의 현존량과 엽록 소 농도 사이의 불일치는 식물플랑크톤에서도 빈번하게 관 찰되는 현상이다. 여러 원인이 있겠지만, 해수를 여과한 후 화학적으로 측정하는 엽록소 농도에는 1 m 내외의 작은 생물의 엽록소도 포함되는 반면, 광학현미경에 의존하는 세 포수 현존량에는 계수의 어려움으로 10 m 이하 작은 세포 를 세포수 산정에 누락되는 경우가 많기 때문이다. 게다가 저서 미세조류의 경우에는 여과 과정도 없이 퇴적물 시료에 직접 아세톤을 넣어 엽록소 농도를 측정하므로 퇴적물 내 모든 생물의 엽록소를 측정한다. 퇴적물 속에는 남세균이나 소형 광합성 진핵생물이 수중보다 더 많기 때문에 세포수와 엽록소 간의 상관관계는 더욱 떨어질 수 있다. 4. 우점 분류군의 분포와 의미 조사 해역에 우점하는 규조류는 조사 시기에 따라 크게 변화하였다. 앞서 기술하였듯이 연구자에 따라 세부적인 종 명은 다르겠지만 동일 연구자에 의한 연구결과는 일관성이 있는 것으로 판단할 수 있다. Table 3, 4는 2009-2010년과 2018년에 양적으로 우점한 저서 규조류를 각각 정리한 것이 다. Table 3을 보면 유류 오염사고가 발생하고 오래 지나지 않은 2009~2010년에는 사고 해역 근처의 오염 해역(학암포, 만리포, 파도리)과 사고 해역에서 비교적 멀리 떨어진 비오 염 해역(백사장, 바람아래) 사이에 저서 규조류의 종조성 차 이가 크다는 것을 알 수 있다. 2009년 봄부터 2010년 여름 까지 거의 모든 시기에 걸쳐 조사 해역의 각 정점에는 각각 다른 종들이 우점하였다. 게다가 Cocconeiopsis kantsiensis, Navicula cf. leptoloba 같은 종들은 오염 해역에서 90%를 상회하는 단일종 극우점 현상을 보였다. C. kantsiensis는 오 염 해역에서 50%를 상회하는 극우점 현상을 몇 차례 반복 하여 유류 오염 지사자로서의 가능성을 보였다. 또한 최고 우점종의 점유율은 대부분 20%를 상회하였다. 이렇게 높은 최고 우점종의 점유율은 상부, 중부, 하부 조간대 모두에서 동일하게 관찰되었고, 2009년에 더욱 뚜렷하였다. 반면 2018년의 경우에는 2009~2010년과 확연하게 다른 결과를 보여주었다. 조사 해역의 모든 정점에 걸쳐 Fallacia floriniae, Halamphora turgida, Planothidium delicatulum, Catenula adhaerens 같은 소수의 몇몇 종들이 조사 시기에 상관없이 최고 우점종으로 반복적으로 출현하였다(Table 4). 또한 한 종에 의한 단일종 우점 현상을 관찰되지 않았고, 위의 4종 이 서로 10~20% 내외의 점유율을 차지하며 우점하였다. 이
Table 4. Seasonal changes in the dominant species of benthic diatoms in the research area, 2018
Season Station 1st Dominant Species % 2nd Dominant Species %
Winter
1 Fallacia floriniae 12.0 Halamphora turgida 8.4 2 Fallacia floriniae 11.9 Planothidium delicatulum 9.5 3 Halamphora turgida 23.8 Planothidium delicatulum 19.0 4 Fallacia floriniae 11.4 Catenula adhaerens 10.1 5 Fallacia floriniae 13.2 Planothidium delicatulum 9.7
Spring
1 Fallacia floriniae 13.6 Halamphora turgida 9.6 2 Planothidium delicatulum 11.5 Fallacia floriniae 10.8 3 Catenula adhaerens 20.0 Halamphora turgida 16.0 4 Halamphora turgida 11.6 Planothidium delicatulum 10.9 5 Fallacia floriniae 16.8 Halamphora turgida 13.3
Summer
1 Fallacia floriniae 13.3 Halamphora turgida 11.3 2 Fallacia floriniae 12.7 Planothidium delicatulum 10.8 3 Catenula adhaerens 26.7 Halamphora turgida 20.0 4 Catenula adhaerens 10.4
Fallacia floriniae
5 Fallacia floriniae 15.9 Halamphora turgida 10.6
Fall
1 Fallacia floriniae 11.7 Planothidium delicatulum 10.8 2 Fallacia floriniae 10.5
Planothidium delicatulum
3 Catenula adhaerens 25.0 Cocconeis scutellum
4 Fallacia floriniae 15.3 Catenula adhaerens 10.2 5 Fallacia floriniae 16.3 Catenula adhaerens 10.9
런 분포 경향을 2017년에도 동일하게 관찰되었다. 저서 규 조류의 이와 같은 분포 특성에 대해 두 시기를 대별하여 정 리하면 2009~2010년에는 한 두 종이 특정 시기에 특정 해 역에서 극도로 높은 점유율을 보인 반면, 2017~2018년 시기 에는 여러 종들이 조사 시기에 상관없이 전체 해역에 걸쳐 서로 비슷한 점유율로 폭넓게 우점하였다. 두 시기의 극단 적인 분포 형태 차이는 유류 오염이 저서 규조류에 미치는 영향을 명확하게 보여주는 것이다. 일반적으로 특정 해역에 오염물질이 투입되어 해양환경이 변하면 그 곳에 가장 먼저 등장하는 생물이 기회종이다. 이런 종들은 오염 환경에 빨 리 적응하여 빠른 속도로 성장하는 것이 특징이다. 2009~ 2010년 시기에 90% 이상의 점유율을 보였던 우점종 C. kantsiensis와 N. cf. leptoloba는 그런 기회종의 예에 해당하 는 것으로 볼 수 있다. 또한 2009~2010년 시기에 각 정점 별로 서로 다른 종들이 우점하였던 것은 유류 오염사고로 유출된 기름 성분이 각 조사 정점에 국지적으로 다른 환경 조건을 제공했기 때문인 것으로 추정된다. 선박에서 유출된 기름이 해안에 도달하는 양과 피해 정도는 사고 지점과 대 상 해역 간의 거리, 대상 해역의 물리해양학적 특성, 지형적 특성, 날씨 등 다양한 요건에 따라 다를 것이다. 이런 요건 은 유출 기름에 대해 각 해역마다 국지적으로 차별화된 환 경 조건을 제공하고 저서 규조류에도 차별화된 서식 환경을 제공한다. 이런 국지적인 환경 변화는 각 해역마다 독특한 저서 규조류가 성장하도록 하고 나아가 그들의 종조성도 변 하게 한다. 따라서 2009~2010년 당시 해역별로 각기 다른 우점종의 출현은 기름 오염의 영향일 수 있으며, 이를 다른 측면에서 평가하면 2017~2018년 시기의 동일종의 폭?은 우 점 현상은 저서 규조류에 대한 유류 오염의 국지적 영향이 더 이상 존재하지 않음을 시사한다.
사
사
본 연구는「HS호 유류유출 사고에 따른 생태계 영향 장기 모니터링」사업의 일환으로 수행되었습니다.참고문헌
국립공원관리공단 국립공원연구원. 2009a. 허베이 스피리트호 유 류유출사고에 따른 태안해안국립공원 일대의 자연자원 정밀 조사. 1,102 pp. 국립공원관리공단 국립공원연구원. 2009b. 허베이 스피리트호 유 류유출 사고에 따른 생태계 영향 장기 모니터링(1차년도). 856 pp. 국립공원관리공단 국립공원연구원. 2010. 허베이 스피리트호 유 류유출 사고에 따른 생태계 영향 장기 모니터링(2차년도). 763 pp. 국립공원관리공단 국립공원연구원. 2011. 허베이 스피리트호 유 류유출 사고에 따른 생태계 영향 장기 모니터링(3차년도). 872 pp. 국립공원관리공단 국립공원연구원. 2012. 허베이 스피리트호 유 류유출 사고에 따른 생태계 영향 장기 모니터링(4차년도). 926 pp. 국립공원관리공단 국립공원연구원. 2013. 허베이 스피리트호 유 류유출 사고에 따른 생태계 영향 장기 모니터링(5차년도). 838 pp. 국립공원관리공단 국립공원연구원. 2014. 허베이 스피리트호 유 류유출 사고에 따른 생태계 영향 장기 모니터링(6차년도). 854 pp. 해양수산부. 2013. 해양환경공정시험기준. 해양수산부고시제2013-230호. http://www.law.go.kr/admRulLsInfoP.do?admRulSeq= 2000000109042Admiraal W. 1984. The ecology of estuarine sediment-inhabiting diatoms. Prog. Phyco.l Res. 3: 269-322.
An SM, DH Choi, JH Lee, H Lee, JH Noh. 2017. Identification of benthic diatoms isolated from the eastern tidal flats of the Yellow Sea: Comparison between morphological and molecular approaches. PLoS ONE 12(6): e0179422. https://doi.org/10.1371/ journal.pone.0179422
Bautista H, K Rahman. 2016a. Effects of Crude Oil Pollution in the Tropical Rainforest Biodiversity of Ecuadorian Amazon Region. Journal of Biodiversity and Environmental Sciences 8(2): 249-254.
Bautista H, K Rahman. 2016b. Review On the Sundarbans Delta Oil Spill: Effects On Wildlife and Habitats. International Research Journal. 1(43), Part 2: 93-96.
Bennion H, J Fluin, GL Simpson. 2004. Assessing eutrophication and reference conditions for Scottish freshwater lochs using subfossil diatoms. J. Appl. Ecol. 41: 124-138.
Berger WH, G Wefer. 1991. Productivity of the glacial ocean: discussion of the iron hypothesis. Limnol Oceanogr. 36: 1899-1918.
Bradshaw EG, AB Nielsen, NJ Anderson. 2006. Using diatoms to assess the impacts of prehistoric, pre-industrial and modern land-use on Danish lakes. Reg Environ Change. 6: 17-24. Cahoon LB. 1999. The role of benthic microalgae in neritic
ecosystems. Oceanography and Marine Biology. An Annual Review. 37: 47-86.
Cahoon LB, GR Beretich, CJ Thomas, A McDonald. 1993. Benthic microalgal production at Stellwagen Bank, Massachusetts Bay. USA. Mar. Ecol. Prog. Ser. 102: 179-185.
CEC 2000. Council of European Communities Directive 2000/60/ EEC of 23 October 2000 establishing a framework for community action in the field of water policy. Official Journal of European Communities, L327/1.
Delgado M, V DeJonge, H Peletler. 1991. Experiments on the resuspension of natural microphytobenthos populations. Marine Biology 108: 321-328.
DeNicola DM, E Eyto. 2004. Using epilithic algal communities to assess trophic status in irish lakes. J. Phycol. 40: 481-495. Deny L. 2003. Environmental changes in man-made coastal dune
pools since 1850 as indicated by sedimentary and epiphytic diatom assemblages (Belgium). Aquatic Conserv: Mar. Freshw.
Ecosyst. 13: 191-211.
Deny L. 2004. Relation of abundance-weighted averages of diatom indicator values to measured environmental conditions in standing freshwaters. Ecological Indicators, 4: 225-275. Dunnet G, D Crisp, G Conan, W Bourne. 1982. Oil Pollution and
Seabird Populations [and Discussion]". Philosophical Transactions of the Royal Society of London B. 297(1087): 413-427. Ehrenhauser F, P Avij, X Shu, V Dugas, I Woodson, T
Liyana-Arachchi, Z Zhang, F Hung, K Valsaraj. 2014. Bubble bursting as an aerosol generation mechanism during an oil spill in the deep-sea environment: laboratory experimental demonstration of the transport pathway. Environ. Sci.: Process. Impacts 16(1): 65-73.
Gaiser E, A Wachnicka, P Ruiz, F Tobias, M Ross. 2005. Diatom indicators of ecosystem change in subtropical coastal wetlands. pp 127-144 in Estuarine Indicators. Bortone, SA. (ed). CRC Press Boca Raton, FL.
Gastineaua R, SY Kim, C Lemieuxc, M Turmelc, A Witkowskia, JG Park, BS Kim, D Mann, E Theriot. 2019. Complete mito-chondrial genome of a rare diatom (Bacillariophyta) Proschkinia and its phylogenetic and taxonomic implications. Mitochondrial DNA part B. 4(1): 25-26.
Gell P, IR Sluiter, J Fluin. 2002. Seasonal and interannual variations in diatom assemblages in Murray River connected wetlands in north-west Victoria, Australia. Mar. Freshw. Res. 53: 981-992. Gowda G, T Gupta, K Rajesh, H Gowda, C Lingadhal, A Ramesh. 2001. Seasonal distribution of phytoplankton in Nethravathi estuary, Mangalore. J Mar Biol Ass India. 43: 31-40. Haubois AG, F Sylvestre, JM Guarini, P Richard, GF Blanchard.
2005. Spatio-temporal structure of the epipelic diatom assemblage from an intertidal mudflat in Marennes-Oléron Bay, France. Estuar. Coast. Shelf. Sci. 64: 385-394.
Holland A, R Zingmark, J Dean. 1974. Quantitative evidence concerning the stabilization of sediments by marine benthic diatoms. Marine Biology 27: 191-196.
Hustedt F. 1955. Marine littoral diatoms of Beaufort, North Carolina. Duke Univ. Mar. Stat. Bull. 6: 1-67.
Hustedt F, NG Jensen. 1985. The pennate diatoms. Koeltz Scientific Books, Koenigstein. 918p.
Kelly MG, A Cazaubon, E Coring, A Dell’Uomo, L Ector, B Goldsmith, H Guasch, J Hurlimann, A Jarlman, B Kawecka, J Kwandrans, R Laugaste, EA Lindstrom, M Leitao, P Marvan, J Padisàk, E Pipp, J Prygiel, E Rott, S Sabater, H van Dam, J Vizinet, 1998. Recommendations for the routine sampling of diatoms for water quality assessments in Europe. J. Appl. Phycol. 10: 215-224.
Kim B-S, S-Y Kim, J-G Park, A Witkowski. 2017, New Records of the Diatom Species (Bacillariophyta) from the Seaweed and Tidal Flats in Korea. Korean Journal of Environmental Biology 35(4): 604-621.
Kim B-S, A Witkowski, J-G Park, C Li, R Trobajo, D Mann, S-Y Kim, M Ashworth, M Bąk, R Gastineau. 2019. Taxonomy and
diversity of a little-known diatom genus Simonsenia (Bacillari-aceae) in the marine littoral: novel taxa from the Yellow Sea and the Gulf of Mexico. Plant Ecology and Evolution 152(2): 248-261.
King L, G Clarke, H Bennion, M Kelly, M Yallop. 2006. Recom-mendations for sampling littoral diatoms in lakes for ecological status assessments. J. Appl. Phycol. 18: 15-25.
King L, P Barker, RI Jones. 2000. Epilithic algal communities and their relationship to environmental variables in lakes of the English Lake District. Freshwat. Biol. 45: 425-442.
Kitner M, A Poulí ková. 2003. Littoral diatoms as indicators for the eutrophication of shallow lakes. Hydrobiologia 506-509: 519-524.
Longhurst A, S Sathyendrenath, T Platt, C Caverhill. 1995. An estimate of global production in the ocean from satellite radiometer data. Journal of Plankton Research. 17: 1245-1271. Mancini L. 2005. Organization of biological monitoring in the European Union. pp. 171-201 in Biological Monitoring of Rivers. Ziglio G., Siligardi M, Flaim G. (eds). John Wiley & Sons, Ltd, London. 469p.
Martin JH, GA Knauer, DM Karl, WW Broenkow. 1987. VERTEX: carbon cycling in the northeast Pacific. Deep-Sea Research. 34: 267-285.
Miller D, R Geider, H Macintyre. 1996. Microphytobenthos: The ecological role of the “Secret Garden” of unvegetated, shallow-water marine habitats. II. Role in sediment stability and shallow-water food webs. Estuaries 19: 202-212.
Nance E, D King, B Wright, R Bullard. 2015. Ambient air con-centrations exceeded health-based standards for fine particulate matter and benzene during the Deepwater Horizon oil spill. Journal of the Air & Waste Management Association 66(2): 224-236.
Pan Y, RJ Stevenson. 1996. Gradient analysis of diatom assemblages in western Kentucky wetlands. J. Phycol. 32: 222-232. Paterson D, R Crawford, C Little. 1990. Subaerial exposure and
changes in sediment stability of intertidal estuarine sediments. Estuarine Coastal and Shelf Science 30: 541-556.
Prygiel J, M Coste, J Bukowska. 1999. Review of the major diatom- based techniques for the quality assessment of rivers. State of art in Europe. pp. 122-127 in Use of Algae for Monitoring Rivers, III. Prygiel J, Whitton BA, Bukowska J. (eds.). Proc. International Symposium, Douai, France 29 September-1 October 1997. Agence de l’Eau Artois-Picardie. 271 p.
Rizzo W, G Iackey, R Christian. 1992. Significance of euphoric, subtidal sediments to oxygen and nutrient cycling in a temperate estuary. Mar. Ecol. Prog. Ser. 86: 51-61.
Round FE. 1957. Studies on Bottom-Living Algae in Some Lakes of the English Lake District: Part II. The Distribution of Bacillariophyceae on the Sediments. J. Ecol. 45: 343-360. Round FE. 1960. Studies on Bottom-Living Algae in Some Lakes
of the English Lake District: IV. The Seasonal Cycles of the cê
Bacillariophyceae. J. Ecol. 48: 529-547.
Schwinghamer P, B Hargrave, D Peer, C Hawkins. 1986. Partitioning of production and respiration among size groups of organisms in an intertidal benthic community. Mar. Ecol. Prog. Ser. 31: 131-142.
Smyth JC. 1955. A study of the benthic diatoms of Loch Sween (Argyll). J. Ecol. 43: 149-171.
Sundback K, V Enoksson, W Granneli, K Pettersson. 1991. Influence of sublittoral microphytobenthos on the oxygen and nutrient flux between sediment and water: A laboratory continuous-flow study. Mar. Ecol. Prog. Ser. 74: 263-279.
Tidwell L, S Allan, S O’Connell, K Hobbie, B Smith, K Anderson. 2015. Polycyclic Aromatic Hydrocarbon (PAH) and Oxygenated PAH (OPAH) Air–Water Exchange during the Deepwater Horizon Oil Spill. Environmental Science & Technology 49(1): 141-149.
Witkowski A, C Riaux-Gobin, Daniszewska-Kowalczyk. 2010. New marine littoral diatom species (Bacillariophyta) from Kerguelen Islands. II. Heteropolar species of Fragilariaceae. Vie et Milieu - Life and environment. 60(3): 265-281.
Witkowski A, H Landge-Bertalot, D Metzeltin. 2000. Diatom flora of marine coasts I. Iconographia Diatomologica 7. A.R.G. Gantner Verlag K.G., Ruggell. 925 pp.
Wolfaardt A, A Williams, L Underhill, R Crawford, P Whittington. 2009. Review of the rescue, rehabilitation and restoration of oiled seabirds in South Africa, especially African penguins Spheniscus demersus and Cape gannets Morus capegnsis, 1983-2005. African Journal of Marine Science 31(1): 31-54. Underwood GJC, J Kromkamp. 1999. Primary production by
phytoplankton and microphytobenthos in estuaries. Adv. Ecol Res. 29: 93-153.
Wang Y-K, RJ Stevenson, PR Sweet, J DiFranco. 2006. Developing and testing diatom indicators for wetlands in the Casco Bay watershed, Maine, USA. Hydrobiologia. 561: 191-206. Zheng L, RJ Stevenson. 2006. Algal assemblages in multiple
habitats of restored and extant wetlands. Hydrobiologia. 561: 221-238.
(2019년 11월 11일 접수; 2019년 11월 24일 수정; 2019년 12월 14일 채택)