저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Master's Thesis in the Department of
Biomedical Sciences
Inhibition of discoidin domain receptor 1
prevents stroma-induced peritoneal
metastasis in gastric carcinomas
Ajou University Graduate School
Major in Cancer Biology
Inhibition of discoidin domain receptor 1
prevents stroma-induced peritoneal
metastasis in gastric carcinomas
Hoon Hur, Advisor
I submit this thesis as the Master's thesis
in the Department of Biomedical Sciences.
August, 2017
Ajou University Graduate School
Major in Cancer Biology
The Master's thesis of Hyejin Jin
in the Department of Biomedical Sciences
is hereby approved.
Thesis Defense Committee
Hoon Hur
Tae Jun Park
Kyeong Sook Choi
Ajou University Graduate School
22th, June, 2017
i
ABSTRACT
Inhibition of discoidin domain receptor 1 prevents
stroma-induced peritoneal metastasis in gastric carcinomas
Introduction: Although the interactions between cancer cells and the tumor
stroma are known to play a role in the peritoneal metastasis of gastric carcinomas (GCs), effective targeting agents that block these interactions have never been reported. Discoidin domain receptor 1 (DDR1) is activated by triple-helix collagens, which are major components of tumor stroma; thus, DDR1 might have a key role in the communication between cancer cells and stroma. The aim of this study was to investigate the effect of DDR1 inhibition on stroma-induced peritoneal metastasis in GCs.
Methods: We analyzed by immunohistochemistry the correlation between DDR1
expression and the pattern of recurrence in GC tissues from a cohort established in a previous study. We also co-cultured human GC cell lines with gastric cancer-associated fibroblasts (CAFs), and investigated DDR1 expression and activation by western blotting. We evaluated the CAF-induced tumorigenic ability of GC cell lines and the effects of a DDR1-specific inhibitor in organotypic cultures and a peritoneal seeding xenograft animal model.
Results: The expression of DDR1 in gastric cancer tissues was significantly
positively associated with early recurrence (p = 0.043) and a high incidence of peritoneal recurrence (p = 0.036). We confirmed that co-culture with CAFs elevated DDR1 protein expression in GC cell lines. CAFs also enhanced GC cell line spheroid formation in organotypic cultures in a tumor cell DDR1-dependent manner. Co-implantation of CAFs with GC cells enhanced peritoneal tumor formation in vivo, an effect that was sensitive to pharmacologic inhibition of DDR1.
ii
cells enhances their peritoneal tumorigenesis, and that the inhibition of DDR1 signaling is an attractive strategy for the treatment of GC peritoneal metastasis.
Keywords: Gastric carcinomas, Peritoneal metastasis, Discoidin domain receptor,
iii
TABLE OF CONTENTS
ABSTRACT ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ i TABLE OF CONTENTS ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ iii LIST OF FIGURES ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ iv LIST OF TABLES ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙v I. INTRODUCTION ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 1
II. MATERIALS AND METHODS ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 3 1. Human samples and immunohistochemical staining ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙3 2. Cell lines and chemicals ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙3 3. CRISPR/Cas9-mediated knockout of DDR1 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 4 4. Western blot ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 5 5. Cell viability test ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 5 6. RT-PCR ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 6
7. Organotypic culture and immunocytochemical staining ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 6 8. Peritoneal xenograft mouse models ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 6
9. Statistical analysis ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙7 III. RESULTS ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 8 1. DDR1 expression correlates with peritoneal recurrence in primary GC
tissues ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 8 2. CAFs enhance the peritoneal tumorigenic potential of GCs∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙11 3. CAFs-induced upregulation of DDR1 in GC cell lines ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙14 4. DDR1 inhibition using the CRISPR/Cas9 system suppresses the effects of CAFs ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙20 5. Blocking DDR1 signaling through 7rh represses the effects of CAFs on GCs ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙22 IV. DISCUSSION ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 30 V. REFERENCES ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙35
iv
LIST OF FIGURES
Figure 1. Genetic inhibition of DDR1 in gastric cancer cell line ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 4 Figure 2. Analysis of recurrence pattern in GC patients according to DDR1 expression ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 9 Figure 3. Tumorigenesis of GC cells was stimulated by CAFs in organotypic
culture ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙12 Figure 4. Peritoneal tumorigenesis of GC cells was stimulated by CAFs ∙∙∙∙∙∙∙∙∙∙∙13
Figure 5. CAFs induced upregulation of DDR1 protein in GC cell lines ∙∙∙∙∙∙∙∙∙∙∙∙∙15
Figure 6. CAFs did not induce upregulation of DDR1 gene in GC cell lines ∙∙∙∙∙∙∙16
Figure 7. CAFs enhanced phosphorylation of DDR1 in organotypic culture ∙∙∙∙∙∙17
Figure 8. CAFs enhanced expression of DDR1 in mouse xenograft models ∙∙∙∙∙18
Figure 9. Screening of collagen expression in GC cell lines and CAFs ∙∙∙∙∙∙∙∙∙∙∙∙∙∙19
Figure 10. Genetic inhibition of DDR1 in GC cells suppressed the CAF-induced tumorigenesis ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙21 Figure 11. Screening of 7rh dose in GC cell lines and CAFs ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 23
Figure 12. 7rh decreased the CAF-induced upregulation and phosphorylation of DDR1 in GC cell line ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙24
Figure 13. 7rh suppressed the CAF-induced tumorigenesis of GC cells in organotypic culture ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 25
Figure 14. 7rh reduced the CAF-induced phosphorylation of DDR1 in organotypic culture ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 26 Figure 15. CAF-stimulated peritoneal tumorigenesis of GC cells was suppressed by 7rh treatment ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 27 Figure 16. CAF-enhanced expression of DDR1 was decreased by 7rh treatment in mouse xenograft models ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 28 Figure 17. 7rh treatment had no side effects ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 29 Figure 18. Schematic displaying the proposed consequences of peritoneal metastasis of gastric carcinomas ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 34
v
LIST OF TABLES
1
I. Introduction
Gastric carcinoma (GC) is one of the most common malignant tumors and the third leading cause of cancer-related death worldwide (1). Although mortality from GCs has recently gradually decreased, patients with late-stage disease still have poor prognoses due to tumor non-resectability or recurrence after resection. Large-scale clinical trials showed that adjuvant chemotherapy following curative resection could reduce the recurrence rate of stage II or III GCs. However, about one-fourth of all patients enrolled in those trials suffered recurrence during the follow-up period, and the peritoneum is one of most common sites of recurrence (2, 3). To date, various studies have reported that patients with peritoneal recurrence have worse responses to treatments than patients with hematogenous or lymphatic metastasis (4-6). However, specific modalities to prevent GC peritoneal metastases have not been established. Thus, new strategies must be developed to prevent GC peritoneal recurrence based on specific molecular biomarkers related to peritoneal metastasis.
Several histologic features of primary GCs have implied the role of the tumor stroma in the peritoneal metastasis of GCs. First, it is well known that the likelihood of peritoneal recurrence increases when primary GCs more deeply invade the gastric wall (6). During the invasion of cancer cells that originate from the mucosal layer of gastrointestinal tract, the cells are exposed to stromal tissues and activate various non-cancerous cells, like fibroblasts and inflammatory cells, contained in the intra-tumoral stroma (7). Reciprocally, activated non-cancerous cells enhance the aggressiveness of cancer cells, thereby increasing their metastatic potential to distal organs (8). Second, the subtypes of GCs with high proportions of tumor stroma preferentially metastasize to the peritoneum (6). Moreover, several studies have reported that cancer-associated fibroblasts (CAFs), non-cancerous cells in the intra-tumoral stroma, contribute to the peritoneal dissemination of GCs (9, 10). Taken together, these findings indicate that the interaction between cancer cells and stromal cells could be a therapeutic target to
2
prevent the peritoneal metastasis of GCs. However, drugs targeting the tumor stroma have not been applied to treat GCs in a clinical setting.
Discoidin domain receptor 1 (DDR1) is a member of the transmembrane receptor tyrosine kinases with a discoidin homology domain in its extracellular region (11-13). Distinct from other receptor tyrosine kinases, DDR1 is activated by the fibrous collagens, like type I, II, III, IV, V, VIII, and XI, which are major components of the extracellular matrix in solid tumors. Various studies have reported that increased DDR1 expression is associated with a poor prognosis in malignant tumors (14-17). Moreover, our previous report described that approximately half of GCs showed positive expression of DDR1, which was related to the depth of GC invasion. In addition, the GC patients with DDR1 expression showed worse overall survival rate than others. We also demonstrated that type I collagen could activate the DDR1 signal transduction pathway and enhance the aggressive phenotype of GC cells (18). However, the significance of DDR1 expression on their pattern of recurrence after the resection of primary GC was not evaluated. If the tumor stroma contributes to the peritoneal recurrence of GCs, DDR1 could be a key regulator that links peritoneal metastasis and tumor stroma in GCs.
Here, we found that DDR1 expression correlated with GC peritoneal recurrence. The aim of this study was to evaluate if tumor stromal cells enhance the peritoneal metastasis of GCs through upregulation of DDR1, and if the inhibition of DDR1 expression could be applied therapeutically to prevent peritoneal recurrence.
3
II. Materials and methods
1. Human samples and immunohistochemical staining
We analyzed the features of GC recurrence in 202 patients, who had undergone curative gastric resections with proper lymphadenectomy at the Department of Surgery, Ajou University Hospital, from May 2003 to December 2005. Here, we examined the incidence of tumor recurrence and its features in that cohort during the follow-up period. The sites of recurrence were classified into groups: the peritoneum, hematogenous sites, and distant lymph nodes. We analyzed the association of DDR1 expression in GCs with the recurrence-free survival rate and the recurrence rate at the specific sites.
2. Cell lines and chemicals
We purchased the GC cell lines MKN45 (KCLB No. 80103), MKN28 (KCLB No. 80102), MKN74 (KCLB No.80104), and KATO-III (KCLB No. 30103) from the Korean Cell Line Bank (Seoul, Korea). These cells were cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Equitech-Bio, Ingram, TX, USA), 1% penicillin/streptomycin (Invitrogen), and 1% amphotericin B (Sigma-Aldrich, St. Louis, MO, USA). The cells were incubated at 37 °C in a humidified atmosphere containing 5% CO2.
CAFs were isolated from fresh GC specimens, as described in our previous study (19). For co-cultures, CAFs were seeded into the upper chambers of 6-well transwells and GC cell lines were seeded in 6-well tissue culture dishes.
4
3. CRISPR/Cas9-mediated knockout of DDR1
We established DDR1-deficient GC lines using the CRISPR-Cas9 system, in order to investigate the role of DDR1 in the interactions between CAFs and GC cell lines (Fig 1). The target sequences for human DDR1 are #1 (forward, 5′-CACCGGTGGAATGTCGCTTCCGGCG-3′; reverse, 5′-AAACCGCCGGAAGCGA CATTCCACC-3′) or #2 (forward: 5′-CACCGCCCCCTAGGTTGTGGCGCAT-3′; reverse, 5′-AAACATGCGCCACAACCTAGGGGGC-3′). The oligonucleotides with BsmB1 restriction sites for guide RNAs were synthesized at Xenotech (Daejeon, Korea), then phosphorylated by T4 kinase (NEB, Ipswich, MA). The phosphorylated oligonucleotides were cloned into LentiCRISPR v2 and the sequences of the cloned plasmids that were extracted from several selected colonies were confirmed at Cosmo Genetech (Seoul, Korea).
MKN28 cells were transfected with pLentiCRISPR-sgRNA DDR1 using Lipofectamine® 2000 (Invitrogen, Carlsbad, CA), according to the manufacturer’s instructions. When the cells were confluent in dishes, they were treated with 1 to 2 µg/ml puromycin for approximately 2 weeks. The silencing of DDR1 in the surviving cells after puromycin selection was validated by western blotting.
Fig. 1. Genetic inhibition of DDR1 in gastric cancer cell line
Establishment of DDR1 knockout MKN28 cell lines using CRISPR-CAS9 system and western blot revealed that DDR1 expression was significantly decreased in
5
4. Western blot
The cells were washed with phosphate-buffered saline and lysed in phospho-specific lysis buffer. Lysates were incubated on ice for 20 min, then centrifuged at 13,000 rpm for 20 min at 4 °C. Protein concentrations were determined by Bradford assay (Bio-Rad, Richmond, CA, USA). Equal amounts of protein from each sample were resolved by sodium dodecyl sulfate– polyacrylamide gel electrophoresis and transferred onto a polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA). The immunoblots were blocked by incubation in 5% skim milk, 25 mM tris(hydroxymathyl) aminomethane–HCl (pH 8.0), 150 mM NaCl, and 0.1% Tween® 20 for 1 h at 25 °C. Membranes were incubated with the following primary antibodies: DDR1 (1:1,000 dilution, #5583, Cell Signaling Technology, Danvers, MA, USA), phosphorylated-DDR1 (1:1,000 dilution, #11994, Cell Signaling Technology), and beta-actin (1:5,000 dilution, sc-47778, Santa Cruz Biotechnology, Dallas, TX), followed by the corresponding HRP-conjugated secondary antibodies (Jackson ImmunoResearch Labs, West Grove, PA, USA). Protein detection was performed with an enhanced chemiluminescence kit (Abclon, Seoul, Korea).
5. Cell viability test
Cells were seeded at 1 × 104 cells per well in 96-well plates, and cell viability was measured using the high sensitive water soluble tetrazolium salt (WST) based cell viability, cytotoxicity assay kit (DoGen, Seoul, Korea). Next, 10 μl of the EZ-Cytox solution was added per well, cells were incubated for 1.5 h, and the absorbance was measured by spectrophotometry at 450 nm. To evaluate the effect of 7rh benzamide on the cell viability, the cells were treated with different concentrations during 72 h.
6
6. RT-PCR
Total RNA was prepared from gastric cancer cell lines and fibroblasts and converted to cDNA using 1 ug of each RNA in a final volume of 20 ul. 3 ul of the cDNA mixture was used for 35 cycles of PCR amplification (denatured at 94 °C for 5 min, annealed at 62 °C (DDR1), or 64 °C (COL1A1) for 30 sec, and extended at 72 °C for 5 min). The primers were as follows: DDR1 (forward, 5′-CGAGCA GGTCATCGAGAACG-3′; reverse, 5′-CCCACCTGCAGTCTCACTG-3′), COL1A1 (forward, 5′-GTCACCCACCGACCAAGAAACC-3′; reverse, 5′-AAGTCCAGGCT GTCCAGGGATG-3′), β-actin (forward, 5′-TCAAGATCATTGCTCCTCCTGAGC-3′; reverse, 5′-TGCTGTCACCTTCACCGTTCCAGT-3′).
7. Organotypic culture and immunocytochemical staining
For organotypic culture experiments, the artificial extracellular matrix (ECM) consisted of a mixture of Matrigel (5 mg/ml, BD Biosciences, San Jose, CA) and collagen I (1.5 mg/ml, BD Biosciences, San Jose, CA) with or without 3,000 CAFs. Gastric cancer cell lines (3,000 cells/well) were seeded on the artificial ECM. To measure the outcomes, we counted the number of spheroids with 200 um or more size and the mean size of largest 5 spheroids at 7th day after seeding the cell under 40x magnification. Immunocytochemical staining was conducted after measuring outcomes approximately 1 week after the plating of the gastric cancer cell lines. Additional methodologic details for organotypic cultures and immunocytochemical staining were previously published by Dang et al. (20).
8. Peritoneal xenograft mouse models
Animal care and handling procedures were performed in accordance with the Ajou University School of Medicine Institutional Animal Care and Use
7
Committee guidelines, and all animal experiments described were approved by the Animal Research Committee of the institution. The peritoneal seeding animal model was established using 6- to 8-week-old male athymic nude mice (Orient Bio, Gyonggi-Do, Korea) weighing 16 to 18 g. We performed 2 types of experiments. In experiment 1, mice were randomly assigned to 3 groups. Mice in the first group were intraperitoneally injected with 1 × 107 cancer cells alone (experiment 1 group 1), or with 1 × 106 (experiment 1 group 2) or 4 × 106 (experiment 1 group 3) CAFs. In experiment 2, mice were assigned to 2 groups. Mice were intraperitoneally injected with 1 × 107 cancer cells only (experiment 2 group 1), or with 1 × 106 CAFs (experiment 2 group 2). Each group was randomly divided into 2 subgroups, and mice were treated orally with the DDR1 inhibitor 7rh (25 mg/kg) or vehicle every 2 days over 3 weeks, starting on day 1 after cell injection. The mice were sacrificed 3 weeks after injection, and the number and the size of the peritoneal nodules were assessed.
9. Statistical analysis
Relapse-free survival rates were evaluated using log-rank tests, and survival curves were generated using the Kaplan–Meier method. The differences between the expression of DDR1 and the patterns of recurrence were evaluated using the χ2 test. All experimental studies were performed independently 3 times. Statistical
analysis was performed using IBM SPSS statistics (version 21 for Mac OS X, IBM, Armonk, NY, USA) and GraphPad Prism (version 6.0 for Mac OS X, GraphPad, La Jolla, CA, USA) software.
8
III. RESULTS
1. DDR1 expression correlates with peritoneal recurrence in primary GC tissues
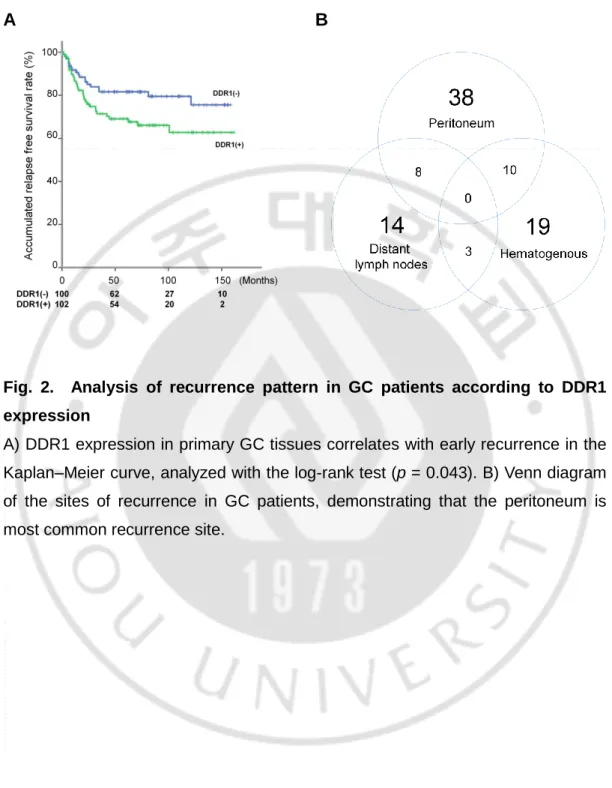
In a previous report, GC tissues with DDR1 expression were shown to be associated with deeper invasion and worse overall surgical outcomes, compared to tissues that did not express DDR1 (18). When we performed additional analyses with follow-up data including recurrence, we found that the patients with DDR1 expression showed significantly shorter relapse-free survival times than DDR1-negative patients (p = 0.043) (Fig. 2A). During the follow-up period, 51 patients (25.2%) presented with recurrence, and 74.5% of all patients with recurrence had metastatic lesions in the peritoneum, the most common site of recurrence (Fig. 2B). The patients with DDR1 expression had significantly higher rates of peritoneal recurrence compared to negative for DDR1. However, hematogenous or distant lymph node recurrences were not significantly related to DDR1 expression (Table 1).
9
A
B
Fig. 2. Analysis of recurrence pattern in GC patients according to DDR1 expression
A) DDR1 expression in primary GC tissues correlates with early recurrence in the Kaplan–Meier curve, analyzed with the log-rank test (p = 0.043). B) Venn diagram of the sites of recurrence in GC patients, demonstrating that the peritoneum is most common recurrence site.
10
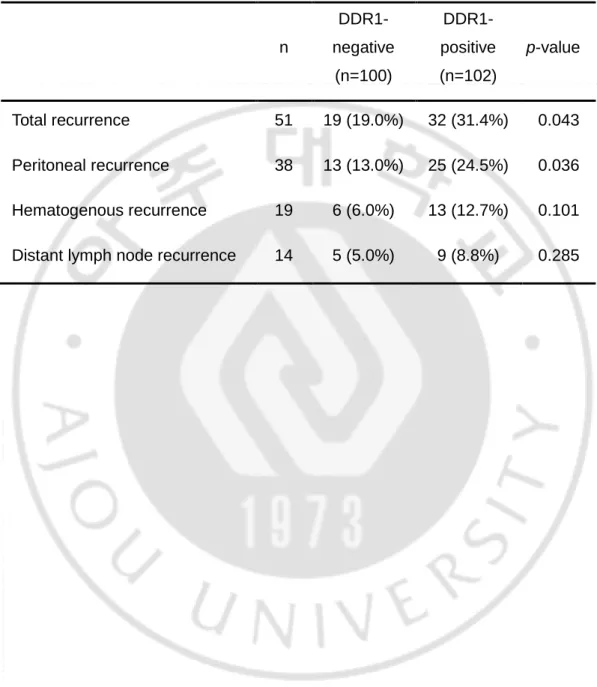
Table 1. Pattern of recurrence according to DDR1 expression in GCs
n DDR1-negative (n=100) DDR1-positive (n=102) p-value Total recurrence 51 19 (19.0%) 32 (31.4%) 0.043 Peritoneal recurrence 38 13 (13.0%) 25 (24.5%) 0.036 Hematogenous recurrence 19 6 (6.0%) 13 (12.7%) 0.101 Distant lymph node recurrence 14 5 (5.0%) 9 (8.8%) 0.285
11
2. CAFs enhance the peritoneal tumorigenic potential of GCs
To identify if CAFs enhance the tumorigenesis of GC cell lines, we conducted organotypic cultures of GC cell lines (Fig. 3A and B). We found that the number of spheroids and the mean size of the 5 largest spheroids in MKN28 and MKN74 cultures were significantly higher in the presence of CAFs relative to cultures on ECM only (Fig. 3C). In addition, we established the peritoneal seeding xenograft mouse model to assess the effects of CAFs on the peritoneal tumorigenesis of GC cell lines in vivo (Fig. 4A). As expected, the number of tumors and mean weight of the largest nodules were significantly increased in groups injected with CAFs, and they depended on the number of CAFs (Fig. 4B and C). These results indicated that CAFs enhance the peritoneal tumorigenesis of GC cell lines.
12
A B
C
Fig. 3. Tumorigenesis of GC cells was stimulated by CAFs in organotypic culture
A) Schematic figure of organotypic culture and B) representative photos of MKN28 spheroids (scale bar, 200 μm). C) The number of colonies larger than 200 μm and the mean sizes of the 5 largest MKN28 and MKN74 spheroids were significantly increased in the presence of CAFs (*p < 0.05, independent t-test).
13
A
B C
Fig. 4. Peritoneal tumorigenesis of GC cells was stimulated by CAFs
Peritoneal tumorigenesis of MKN28 gastric cancer cell line in BALB/c nude mice (n=8, each group). A) Representative photos of peritoneal nodules (arrow) 18 days after injection of MKN28 with/without CAFs into the abdominal cavity. B) The number of tumor nodules in each group and C) mean weight of the largest nodules (*p < 0.05, **p < 0.01, independent t-test).
14
3. CAFs-induced upregulation of DDR1 in GC cell lines
We used a transwell co-culture system to investigate CAF-induced changes in DDR1 expression and activation. In GC cell lines, MKN28 and MKN45, that co- culture with CAFs using the transwell co-culture system increased the expression and phosphorylation of DDR1 relative to cultures without CAFs (Fig. 5). However,
DDR1 gene expression in MKN28 cells was not increased despite co-culture with
CAFs (Fig. 6). We also identified the phosphorylation of DDR1 in organotypic cultures of MKN28 through immunocytochemistry (Fig. 7). Moreover, we identified through immunohistochemistry the upregulation of DDR1 in the peritoneal seeding xenograft mouse model, in the groups that received co-injection of MKN28 and CAFs (Fig. 8). We identified that expression of the COL1A1 gene is almost undetectable in GC cell lines, whereas it is highly expressed in CAFs (Fig. 9). These results indicated that CAF-induced upregulation of DDR1 are involved in the peritoneal tumorigenesis of GCs.
15
Fig. 5. CAFs induced upregulation of DDR1 protein in GC cell lines
Western blotting showed that co-culture with CAFs enhanced DDR1 expression and phosphorylation in MKN28 and MKN45 cells. The pDDR1, DDR1 and β-actin bands were quantified with densitometric program Image J and then relative pDDR1 and DDR1 bands intensities were normalized to β-actin band intensity.
16
Fig. 6. CAFs did not induce upregulation of DDR1 gene in GC cell lines
RT-PCR showed that there was no difference in DDR1 gene level between MKN28 co-cultured with CAFs and MKN28 cultured without CAFs. The DDR1 and β-actin bands were quantified with densitometric program Image J and then relative DDR1 band intensity was normalized to β-actin band intensity.
17
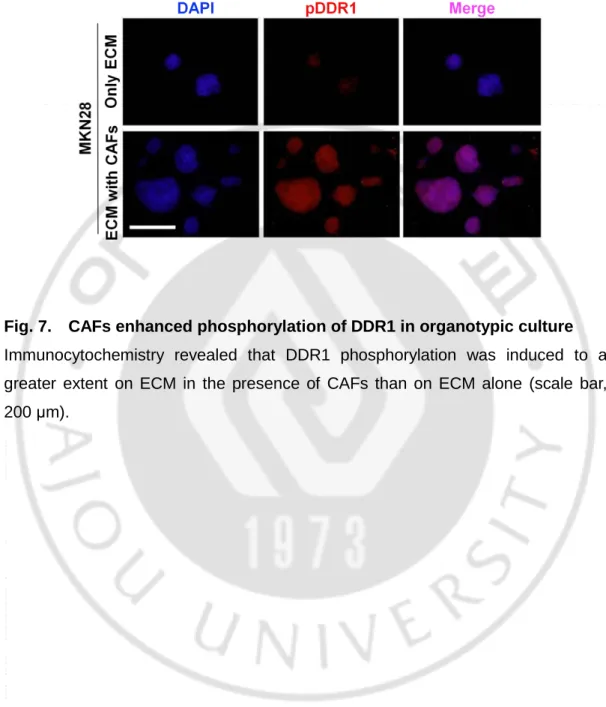
Fig. 7. CAFs enhanced phosphorylation of DDR1 in organotypic culture
Immunocytochemistry revealed that DDR1 phosphorylation was induced to a greater extent on ECM in the presence of CAFs than on ECM alone (scale bar, 200 μm).
18
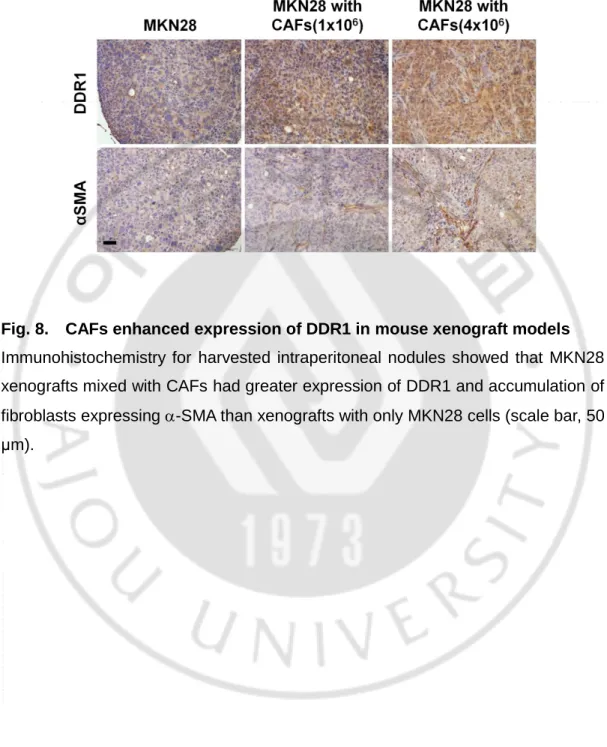
Fig. 8. CAFs enhanced expression of DDR1 in mouse xenograft models
Immunohistochemistry for harvested intraperitoneal nodules showed that MKN28 xenografts mixed with CAFs had greater expression of DDR1 and accumulation of fibroblasts expressing -SMA than xenografts with only MKN28 cells (scale bar, 50 μm).
19
Fig. 9. Screening of collagen expression in GC cell lines and CAFs
Collagen was definitely expressed higher level in CAFs than gastric cancer cell lines. The COL1A1 and β-actin bands were quantified with densitometric program Image J and then relative COL1A1 band intensity was normalized to β-actin band intensity.
20
4. DDR1 inhibition using the CRISPR/Cas9 system suppresses the effects of CAFs
We conducted organotypic cultures for GC cell lines using DDR1-knockout MKN28. We found that the DDR1-knockout MKN28 cells generated equivalent numbers of spheroids with a similar mean size of the 5 largest spheroids in the ECM only and ECM with CAFs conditions (Fig. 10A and B). Moreover, we investigated the activation of DDR1 in the DDR1-knockout MKN28 cells from the organotypic cultures through immunocytochemistry. As expected, the ECM with CAFs conditions did not enhance the activation of DDR1 in the DDR1-knockout MKN28 cells (Fig. 10C). These data suggested that the genetic inhibition of DDR1 in GC cells suppressed the effect of CAFs.
21
A B
C
Fig. 10. Genetic inhibition of DDR1 in GC cells suppressed the CAF-induced tumorigenesis
Organotypic cultures of DDR1-knockout MKN28 cells. A) Representative photos of colonies (scale bar, 200 μm). B) No differences were observed in the number of colonies larger than 200 μm or the mean size of the 5 largest spheroids from
DDR1-knockout MKN28 cells in the presence or absence of CAFs. C)
Immunocytochemistry showed that DDR1-knockout MKN28 cells did not have enhanced DDR1 phosphorylation on ECM in the presence of CAFs, unlike DDR1-positive MKN28 cells (scale bar, 200 μm).
22
5. Blocking DDR1 signaling through 7rh represses the effects of CAFs on GCs
We next evaluated the effect of 7rh, a specific inhibitor of DDR1, which may be a new candidate therapeutic agent. First, we demonstrated that low-dose 7rh does not affect the viability of gastric cancer cell lines or fibroblasts (Fig. 11), and that it reduces DDR1 expression in MKN28 (Fig. 12). Then, we conducted organotypic cultures of GC cell lines with 7rh. The organotypic cultures revealed that 7rh reduced the number of spheroids and the mean size of the 5 largest colonies despite co-culture with CAFs (Fig. 13A and B). Immunocytochemistry for the organotypic cultures showed that 7rh suppressed the phosphorylation of DDR1 in MKN28 (Fig. 14). Moreover, to further clarify the effect of 7rh, we performed in vivo experiments using the peritoneal seeding xenograft mouse model of gastric cancer (Fig. 15A). As expected, 7rh significantly reduced the number of tumor nodules in the CAF co-injection group (Fig. 15B). Immunohistochemistry on the harvested tumor nodules revealed that 7rh reduced the expression of DDR1 in peritoneal tumors (Fig. 16). However, the weight of mice in each group did not show the difference during the treatment (Fig 17). Thus, 7rh suppresses the CAF-induced peritoneal tumorigenesis of GCs without side effects through inhibition of DDR1 signaling
23
Fig. 11. Screening of 7rh dose in GC cell lines and CAFs
24
Fig. 12. 7rh decreased the CAF-induced upregulation and phosphorylation of DDR1 in GC cell line
Western blotting revealed that 7rh reduced DDR1 expression in MKN28. The pDDR1, DDR1 and β-actin bands were quantified with densitometric program Image J and then relative pDDR1 and DDR1 bands intensities were normalized to β-actin band intensity.
25
A
B
Fig. 13. 7rh suppressed the CAF-induced tumorigenesis of GC cells in organotypic culture
Organotypic cultures of MKN28 cells with 7rh treatment. A) Representative photos of colonies (scale bar, 200 μm). B) The number of colonies larger than 200 μm and the mean size of the 5 largest spheroids from MKN28 cells were markedly decreased by 7rh treatment on ECM with CAFs (*p < 0.05, **p < 0.01, independent t-test).
26
Fig. 14. 7rh reduced the CAF-induced phosphorylation of DDR1 in organotypic culture
Immunocytochemistry showed that DDR1 phosphorylation in MKN28 was suppressed by 7rh treatment on ECM in the presence of CAFs (scale bar, 200 μm).
27
A
B
Fig. 15. CAF-stimulated peritoneal tumorigenesis of GC cells was suppressed by 7rh treatment
Peritoneal tumorigenesis of gastric cancer cell lines in BALB/c nude mice with 7rh treatment (n=6, each group). Mice were treated with 25 μg/g 7rh 3 times per week for 3 weeks. A) Representative photos of peritoneal nodules (arrow) from mice that were treated with 7rh for 18 days after MKN28 injection into the abdominal cavity with/without CAFs. B) The number of tumor nodules was reduced by 7rh treatment in mice injected with MKN28 and CAFs, whereas 7rh treatment did not statistically affect the number of tumor nodules in mice injected with MKN28 cells only. The effects on the MKN45 tumor nodules were the same as for MKN28.
28
Fig. 16. CAF-enhanced expression of DDR1 was decreased by 7rh treatment in mouse xenograft models
Immunohistochemistry on harvested intraperitoneal nodules showed that 7rh reduced DDR1 expression in MKN28 xenograft tumors (scale bar, 50 μm).
29
Fig. 17. 7rh treatment had no side effects
Weight of mice showed the similar pattern among each treatment groups during treatment.
30
IV. DISCUSSION
This study reveals the potential of DDR1 as a novel therapeutic target to prevent stroma-induced GC peritoneal recurrence after resection. In our cohort of patients, who underwent curative resection for GCs, we found that DDR1 expression in primary tumors could be a biomarker to predict peritoneal recurrence. The in vitro and in vivo experiments showed that DDR1 was upregulated in GC cells stimulated with CAFs. Furthermore, inhibition of DDR1 induced significant anti-tumorigenic effects in CAF-stimulated GC cells in organotypic cultures and a peritoneal xenograft mouse model. Our findings corroborate the favorable evidence for the use of a DDR1 inhibitor to prevent peritoneal recurrence after curative resection for GCs and are summarized in Fig. 18.
Peritoneal metastasis is a distinctive pattern of recurrence in GCs, and is associated with worse outcomes than hematogenous or lymphatic metastasis (21, 22). In particular, small peritoneal seeding nodules in the peritoneal cavity are not adequately detected in imaging studies like abdominal computed tomography, so peritoneal recurrence is diagnosed at late stages and associated with poor outcomes. Therefore, a prediction of peritoneal metastasis through analysis of the resected primary tumor is required to suggest additional modalities to prevent metastasis after curative resection for GCs. However, reliable markers for site-specific recurrence have only rarely been investigated in GCs. Recently, a transcriptome analysis of curatively resected GC tissues revealed that SYT8 could be a candidate to predict peritoneal recurrence during the follow-up period (23). However, the researchers obtained transcriptome data from only 16 patients, so the statistical power of the data might be insufficient to confirm SYT8 as a reliable marker. In addition, therapeutic agents targeting SYT8 protein are not available. Others reported that the infiltration of S100-positive dendritic cells could be a reliable marker for peritoneal recurrence of GCs (24). However, the studies lacked supportive experimental data to demonstrate the correlation between dendritic
31
cells and the peritoneal tumorigenesis of GC cells. In the present study, we analyzed the clinical data of 202 GC patients, whose primary carcinoma tissues were processed for DDR1 expression by immunohistochemistry. These results showed that DDR1 expression in patients was related to peritoneal recurrence (the most common recurrence), but not to hematogenous or lymphatic metastasis. Moreover, our experimental studies indicated that stroma-induced DDR1 upregulation in GC cell lines enhances their peritoneal tumorigenesis, whereas the inhibition of DDR1 reduces it. Our findings provide support for the use of DDR1 expression as a predictive biomarker for peritoneal recurrence in patients with GCs, and for the application of 7rh, a DDR1 inhibitor, for the prevention of peritoneal recurrence after curative resection.
Our previous report using the same cohort as the present study indicated that the proportion of DDR1-positive cells in primary GC tumors increased among the tumor cells that had more deeply invaded the gastric wall (18). Other studies, which evaluated the clinical relevance of DDR1 expression for gastrointestinal malignancies, like gastric and esophageal carcinomas, showed similar results (25, 26). Carcinoma cells of the gastrointestinal tract usually originate from epithelial cells in the mucosal layer, and consequently invade the submucosa, the muscle, and the serosal layers. The initial step of cancer invasion is the degradation of the basement membrane, followed by increased cross-talk between the invading carcinoma cells and the stromal tissue (27). Stromal tissue consists of various components, including non-cancerous cells and ECM. Among those components, fibroblasts are essential to contribute to ECM remodeling by secreting proteases like metalloproteinases, which effectively degrade ECM (28). In addition, fibroblasts can produce various soluble autocrine and paracrine secretory proteins that regulate the activity of the surrounding cells as well as themselves (29). In the present study, we speculated that GC cells invading the gastric wall upregulate DDR1 through interaction with CAFs. Our experimental data support a role for CAFs in enhancing the upregulation of DDR1 in co-cultures, organotypic cultures, and peritoneal xenograft tumors, but DDR1 gene transcription was not changed after co-culture with CAFs. These findings are consistent with a recent report that
32
insulin like growth factor 1 (IGF-1) induces upregulation of DDR1 by suppressing miR-119 through the PI3K/AKT pathway, and not by enhancing the transcription of DDR1 (30). Although several reports indicate that IGF-1 secreted from fibroblasts stimulates the migration, survival, and growth of cancer cells (31, 32), the exact mechanisms of CAF-induced upregulation of DDR1 in GCs should be more fully explored in the future.
In the present study, we hypothesized that the upregulation of DDR1 is essential for the tumorigenic ability of CAF-stimulated, free-floating cancer cells in the peritoneum. Our data support this hypothesis, as DDR1-knockout cells did not undergo CAF-stimulated increases in colony number or clone size. There are several studies describing the ability of DDRs to promote tumor growth in metastatic sites like the bone and liver (33, 34). However, there had been no proof of the role of DDR1 in the peritoneal metastasis of cancer cells. In the early stage of peritoneal metastasis, the floating cells disseminated from GC primary tumors must survive in the peritoneal cavity (35). In some tumor cells, DDR1 directly enhances pro-survival mechanisms through interactions with Notch1 or the activation of NF-B and the downstream effector COX-2 (36, 37). Finally, the disseminated cells aggregate into metastatic nodules on the peritoneal surface. DDR1 is known to play a critical role in supporting cell–cell adhesion in a variety of non-cancerous cells (38-40) and malignant cell lines (41). Studies suggest that DDR1 stabilizes E-cadherin at the cell surface and promotes cell-to-cell aggregation (38, 40). Although we have not proposed an exact mechanism, our results support the hypothesis that the upregulation of DDR1 in GC cells is a critical step in the formation of tumor nodules from disseminated cancer cells on the peritoneal surface.
The efficacy of adjuvant intraperitoneal chemotherapy has been studied for the prevention of peritoneal recurrence after curative resection in patients with locally advanced GCs. Cisplatin and mitomycin C have been used as cytotoxic agents for intraperitoneal administration, but their survival benefits have been controversial (42). A meta-analysis showed that the risk of postoperative abscess in the abdominal cavity was increased by 137% in patients with intraabdominal
33
chemotherapy relative to those receiving surgery alone (43). Those findings have resulted in physicians doubting the benefit of intraperitoneal chemotherapy for the prevention of peritoneal recurrence in GCs. Based on the findings of the present study, we suggest that the novel, orally available drug 7rh may prevent the tumorigenesis of free-floating cancer cells in the abdominal cavity. In the experiments performed by Gao et al., 7rh potentially inhibited the proliferation, adhesion, and tumorigenicity of cancer cells expressing high levels of DDR1, and had a good pharmacokinetic profile with oral bioavailability of 67.4% (44). Another group described that 7rh could attenuate DDR1 expression in nadopharyngeal cancer cells as our data in GC cells, and they suggested the Janus kinase (JAK1)-signal transducer and activator of transcription (STAT3) (JAK1)-signaling pathway as the downstream of DDR1 (45). Our in vivo study revealed no adverse effects of oral administration of 7rh, and we confirmed that 7rh reduced the expression of DDR1 in xenograft tumors. These results suggest that 7rh could be orally administered to patients after resection for primary GCs, to prevent peritoneal recurrence in patients with high risk.
In conclusion, intra-tumoral stromal cells, particularly CAFs, enhance the upregulation of DDR1 in GCs, and DDR1 is a key factor in CAF-induced tumorigenesis. We suggest that treatment with 7rh, a specific inhibitor for DDR1, in the adjuvant setting after curative resection might prevent peritoneal recurrence of GCs.
34
Fig. 18. Schematic displaying the proposed consequences of peritoneal metastasis of gastric carcinomas
This includes invasion of cancer cells from mucosal layer to serosa, and activation of cancer cells by cancer-associated fibroblasts during invasion. Activated cancer cells increase DDR1 expression and are exfoliated into the peritoneal cavity as free floating cells. Finally those cells can make tumor nodules at the peritoneal surface.
35
V. REFERENCES
1. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. International journal of cancer Journal international du cancer. 2015;136(5):E359-86.
2. Bang YJ, Kim YW, Yang HK, Chung HC, Park YK, Lee KH, et al. Adjuvant capecitabine and oxaliplatin for gastric cancer after D2 gastrectomy (CLASSIC): a phase 3 open-label, randomised controlled trial. Lancet. 2012;379(9813):315-21. 3. Sakuramoto S, Sasako M, Yamaguchi T, Kinoshita T, Fujii M, Nashimoto A, et al. Adjuvant chemotherapy for gastric cancer with S-1, an oral fluoropyrimidine. The New England journal of medicine. 2007;357(18):1810-20.
4. Ahmed A, Ukwenya AY, Makama JG, Mohammad I. Management and outcome of gastric carcinoma in Zaria, Nigeria. Afr Health Sci. 2011;11(3):353-61. 5. Maehara Y, Hasuda S, Koga T, Tokunaga E, Kakeji Y, Sugimachi K. Postoperative outcome and sites of recurrence in patients following curative resection of gastric cancer. Br J Surg. 2000;87(3):353-7.
6. Roviello F, Marrelli D, de Manzoni G, Morgagni P, Di Leo A, Saragoni L, et al. Prospective study of peritoneal recurrence after curative surgery for gastric cancer. Br J Surg. 2003;90(9):1113-9.
7. Clark AG, Vignjevic DM. Modes of cancer cell invasion and the role of the microenvironment. Curr Opin Cell Biol. 2015;36:13-22.
36
Cancer Associated Fibroblasts for the Regulation of Cancer Invasion. Cancers (Basel). 2015;7(4):2054-62.
9. Kurashige J, Mima K, Sawada G, Takahashi Y, Eguchi H, Sugimachi K, et al. Epigenetic modulation and repression of miR-200b by cancer-associated fibroblasts contribute to cancer invasion and peritoneal dissemination in gastric cancer. Carcinogenesis. 2015;36(1):133-41.
10. Yamaguchi H, Yoshida N, Takanashi M, Ito Y, Fukami K, Yanagihara K, et al. Stromal fibroblasts mediate extracellular matrix remodeling and invasion of scirrhous gastric carcinoma cells. PloS one. 2014;9(1):e85485.
11. Alves F, Vogel W, Mossie K, Millauer B, Hofler H, Ullrich A. Distinct structural characteristics of discoidin I subfamily receptor tyrosine kinases and complementary expression in human cancer. Oncogene. 1995;10(3):609-18. 12. Valiathan RR, Marco M, Leitinger B, Kleer CG, Fridman R. Discoidin domain receptor tyrosine kinases: new players in cancer progression. Cancer metastasis reviews. 2012;31(1-2):295-321.
13. Vogel W. Discoidin domain receptors: structural relations and functional implications. FASEB J. 1999;13 Suppl:S77-82.
14. Ford CE, Lau SK, Zhu CQ, Andersson T, Tsao MS, Vogel WF. Expression and mutation analysis of the discoidin domain receptors 1 and 2 in non-small cell lung carcinoma. British journal of cancer. 2007;96(5):808-14.
15. Hou G, Vogel W, Bendeck MP. The discoidin domain receptor tyrosine kinase DDR1 in arterial wound repair. The Journal of clinical investigation. 2001;107(6):727-35.
37
16. Quan J, Yahata T, Adachi S, Yoshihara K, Tanaka K. Identification of receptor tyrosine kinase, discoidin domain receptor 1 (DDR1), as a potential biomarker for serous ovarian cancer. Int J Mol Sci. 2011;12(2):971-82.
17. Yang SH, Baek HA, Lee HJ, Park HS, Jang KY, Kang MJ, et al. Discoidin domain receptor 1 is associated with poor prognosis of non-small cell lung carcinomas. Oncology reports. 2010;24(2):311-9.
18. Hur H, Ham IH, Lee D, Jin H, Aguilera KY, Oh HJ, et al. Discoidin domain receptor 1 activity drives an aggressive phenotype in gastric carcinoma. BMC Cancer. 2017;17(1):87.
19. Lee D, Ham IH, Son SY, Han SU, Kim YB, Hur H. Intratumor stromal proportion predicts aggressive phenotype of gastric signet ring cell carcinomas. Gastric Cancer. 2016.
20. Dang TT, Prechtl AM, Pearson GW. Breast cancer subtype-specific interactions with the microenvironment dictate mechanisms of invasion. Cancer research. 2011;71(21):6857-66.
21. Dicken BJ, Bigam DL, Cass C, Mackey JR, Joy AA, Hamilton SM. Gastric adenocarcinoma: review and considerations for future directions. Ann Surg. 2005;241(1):27-39.
22. Van Cutsem E, Sagaert X, Topal B, Haustermans K, Prenen H. Gastric cancer. Lancet. 2016;388(10060):2654-64.
23. Kanda M, Murotani K, Kobayashi D, Tanaka C, Yamada S, Fujii T, et al. Postoperative adjuvant chemotherapy with S-1 alters recurrence patterns and prognostic factors among patients with stage II/III gastric cancer: A propensity
38
score matching analysis. Surgery. 2015;158(6):1573-80.
24. Saito H, Osaki T, Murakami D, Sakamoto T, Kanaji S, Ohro S, et al. Prediction of sites of recurrence in gastric carcinoma using immunohistochemical parameters. J Surg Oncol. 2007;95(2):123-8.
25. Lu X, Nemoto S, Lin A. Identification of c-Jun NH2-terminal protein kinase (JNK)-activating kinase 2 as an activator of JNK but not p38. The Journal of biological chemistry. 1997;272(40):24751-4.
26. Xie R, Wang X, Qi G, Wu Z, Wei R, Li P, et al. DDR1 enhances invasion and metastasis of gastric cancer via epithelial-mesenchymal transition. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine. 2016;37(9):12049-59.
27. Glentis A, Gurchenkov V, Matic Vignjevic D. Assembly, heterogeneity, and breaching of the basement membranes. Cell Adh Migr. 2014;8(3):236-45.
28. Chang HY, Chi JT, Dudoit S, Bondre C, van de Rijn M, Botstein D, et al. Diversity, topographic differentiation, and positional memory in human fibroblasts. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(20):12877-82.
29. Nakamura T, Matsumoto K, Kiritoshi A, Tano Y, Nakamura T. Induction of hepatocyte growth factor in fibroblasts by tumor-derived factors affects invasive growth of tumor cells: in vitro analysis of tumor-stromal interactions. Cancer research. 1997;57(15):3305-13.
30. Mata R, Palladino C, Nicolosi ML, Lo Presti AR, Malaguarnera R, Ragusa M, et al. IGF-I induces upregulation of DDR1 collagen receptor in breast cancer
39
cells by suppressing MIR-199a-5p through the PI3K/AKT pathway. Oncotarget. 2016;7(7):7683-700.
31. Satyamoorthy K, Li G, Vaidya B, Kalabis J, Herlyn M. Insulin-like growth factor-I-induced migration of melanoma cells is mediated by interleukin-8 induction. Cell Growth Differ. 2002;13(2):87-93.
32. Satyamoorthy K, Li G, Vaidya B, Patel D, Herlyn M. Insulin-like growth factor-1 induces survival and growth of biologically early melanoma cells through both the mitogen-activated protein kinase and beta-catenin pathways. Cancer research. 2001;61(19):7318-24.
33. Badiola I, Olaso E, Crende O, Friedman SL, Vidal-Vanaclocha F. Discoidin domain receptor 2 deficiency predisposes hepatic tissue to colon carcinoma metastasis. Gut. 2012;61(10):1465-72.
34. Valencia K, Ormazabal C, Zandueta C, Luis-Ravelo D, Anton I, Pajares MJ, et al. Inhibition of collagen receptor discoidin domain receptor-1 (DDR1) reduces cell survival, homing, and colonization in lung cancer bone metastasis. Clinical cancer research : an official journal of the American Association for Cancer Research. 2012;18(4):969-80.
35. Kanda M, Kodera Y. Molecular mechanisms of peritoneal dissemination in gastric cancer. World J Gastroenterol. 2016;22(30):6829-40.
36. Das S, Ongusaha PP, Yang YS, Park JM, Aaronson SA, Lee SW. Discoidin domain receptor 1 receptor tyrosine kinase induces cyclooxygenase-2 and promotes chemoresistance through nuclear factor-kappaB pathway activation. Cancer research. 2006;66(16):8123-30.
40
37. Kim HG, Hwang SY, Aaronson SA, Mandinova A, Lee SW. DDR1 receptor tyrosine kinase promotes prosurvival pathway through Notch1 activation. The Journal of biological chemistry. 2011;286(20):17672-81.
38. Eswaramoorthy R, Wang CK, Chen WC, Tang MJ, Ho ML, Hwang CC, et al. DDR1 regulates the stabilization of cell surface E-cadherin and E-cadherin-mediated cell aggregation. Journal of cellular physiology. 2010;224(2):387-97. 39. Wang CZ, Yeh YC, Tang MJ. DDR1/E-cadherin complex regulates the activation of DDR1 and cell spreading. Am J Physiol Cell Physiol. 2009;297(2):C419-29.
40. Yeh YC, Wu CC, Wang YK, Tang MJ. DDR1 triggers epithelial cell differentiation by promoting cell adhesion through stabilization of E-cadherin. Mol Biol Cell. 2011;22(7):940-53.
41. Hidalgo-Carcedo C, Hooper S, Chaudhry SI, Williamson P, Harrington K, Leitinger B, et al. Collective cell migration requires suppression of actomyosin at cell-cell contacts mediated by DDR1 and the cell polarity regulators Par3 and Par6. Nat Cell Biol. 2011;13(1):49-58.
42. Feingold PL, Kwong ML, Davis JL, Rudloff U. Adjuvant intraperitoneal chemotherapy for the treatment of gastric cancer at risk for peritoneal carcinomatosis: A systematic review. J Surg Oncol. 2017;115(2):192-201.
43. Yan TD, Black D, Sugarbaker PH, Zhu J, Yonemura Y, Petrou G, et al. A systematic review and meta-analysis of the randomized controlled trials on adjuvant intraperitoneal chemotherapy for resectable gastric cancer. Ann Surg Oncol. 2007;14(10):2702-13.
41
44. Gao M, Duan L, Luo J, Zhang L, Lu X, Zhang Y, et al. Discovery and optimization of 3-(2-(Pyrazolo[1,5-a]pyrimidin-6-yl)ethynyl)benzamides as novel selective and orally bioavailable discoidin domain receptor 1 (DDR1) inhibitors. Journal of medicinal chemistry. 2013;56(8):3281-95.
45. Lu QP, Chen WD, Peng JR, Xu YD, Cai Q, Feng GK, et al. Antitumor activity of 7RH, a discoidin domain receptor 1 inhibitor, alone or in combination with dasatinib exhibits antitumor effects in nasopharyngeal carcinoma cells. Oncol Lett. 2016;12(5):3598-608.
42
국문요약
기질 세포에 의해 유도되는 위암 복막 전이에서
discoidin domain receptor 1의 역할에 대한 연구
아주대학교 대학원 의생명과학과 진 혜 진
(지도교수: 허 훈)
암 세포와 종양 기질 세포 사이의 상호작용이 위암의 복막 전이에 관여한다는 사실은 이미 알려져 있지만, 이를 효과적으로 차단할 표적 물질은 아직 밝혀지지 않은 실정이다. Discoidin domain receptor 1 (DDR1)은 종양 기질의 주요한 구성물질인 삼중 나선 구조의 콜라겐에 의해 활성화된다. 그러므로, DDR1은 암 세포와 종양 기질 세포 사이의 커뮤니케이션에 중요한 역할을 할 것으로 생각된다. 본 연구는 DDR1을 억제함에 따라 기질 세포에 의해 유도되는 위암 복막 전이의 개선 여부를 밝혀내고자 하였다. 본 연구에서는 이전 연구에서 설립한 위암 환자들의 집단으로부터 얻은 위암 조직에서의 DDR1 발현과 재발 패턴의 연관성을 면역조직화학염색법을 이용하여 분석하였다. 그 결과, 위암 조직의 DDR1 발현은 짧은 무재발 생존 기관과 관련성을 (p = 0.043) 보였고 재발 부위 중 복막 전이의 발생 비율과 의미 있는 연관성을 (p = 0.036) 나타냈다. 이러한 임상데이터 분석 결과를 바탕으로 위암 세포주를 위암 관련 섬유모세포와 동시 배양함으로써 DDR1 단백질의 발현이 증가하는 것을 다양한 실험 기법을 통해 확인하였다. 또한, 위암 관련 섬유모세포가 위암 세포의 DDR1을 통해 기관형 배양에서 스페로이드 형성을 증가시키고, 복막 이종이식 마우스 실험에서 복막의 종양
43 형성 능력을 증가시키는 것을 확인하였다. 더 나아가 이러한 현상들이 DDR1에 특이적으로 작용하는 억제제인 7rh에 의해 저해됨을 확인하였다. 결과적으로, 본 연구에서는 위암 관련 섬유모세포에 의해 유도되는 위암 세포의 DDR1 발현 증가가 위암 복막 전이의 악화에 관여함을 밝혔으며, 앞으로 DDR1에 대한 억제가 위암 복막 전이의 치료 전략이 될 수 있을 것으로 기대한다.